
临安杉木生物量估算模型及其通用性研究
2018-06-20 02:32:48孙燕飞王国英侯建花莫路锋
浙江林业科技 2018年2期
孙燕飞,王国英,侯建花,莫路锋
(1. 浙江农林大学 信息工程学院,浙江 杭州 311300;2. 浙江省景宁县林业局,浙江 景宁 323500)
生物量在研究森林生态系统功能方面具有重要意义,这一指标既能反映森林在调节全球碳平衡方面的重要作用,又可帮助管理者实现林地的可持续经营和更好的利用[1]。
目前森林生物量测定方法中的皆伐实测法因费时费力、难度大、破坏性大等缺点,不利于推广,一般仅在检验其它测定方法的精度时才使用,因此发现一种更方便准确且通用的测定方法具有现实意义。研究发现,应用较多的生物量测算方法是模型估算法[1-2],该方法多以胸径(D)、树高(H)等测树因子为自变量,对林木的生物量进行推算[3]。模型估算法前期必须准备实测样本数据,从而根据样本数据建立生物量模型,后期可直接根据相关的统计资料来估算对应林种的生物量。该方法不仅缩短了大范围森林生物量的调查周期,而且对林木生物量有较高的估算精度,具有较好的现实意义。该模型形式多样,其中应用较多的是幂函数模型、线性模型、指数模型和多项式模型等[2,4-6]。
杉木Cunninghamia lanceolata是我国栽培历史悠久的重要用材林树种,多为人工或半人工的纯林,在我国速生树种中占据着绝对的优势地位,在全国人工林种植面积中的比重已达到了28.54%[2],是我国南方地区重要的乡土树种。浙江省是我国历史上重要的杉木产地,主要分布在海拔1 000 m以下土壤肥沃、温暖湿润的山坡地块中。近年来有关杉木生物量的研究报道也不断深入。林开敏等通过模型优选法研究得到了6年生杉木和马尾松Pinus massoniana的生物量模型[7];唐坤银等以湖南省4县(市)为例建立并优选了杉木分器官和全株的生物量模型[8];李燕等从模型精度和通用性方面对杉木生物量模型进行优选[2];范仁志采用多模型优选法对杉木树根生物量进行研究[9]。以上学者都是从模型优选的角度对杉木生物量展开的研究,研究时仍存在对研究对象的筛选过于精细、忽视适用性等不足,且大多局限于单一树龄或器官[2,7-11],针对杉木林整个龄组(幼龄林、中龄林、近熟林和成熟林)生物量模型的研究偏少,对一种模型在不同龄组间运用时的精度状况进行描述的研究更是少见。因此,本研究选择了浙江省临安区为研究区域,以不同龄组的杉木林实测数据为样本依据,选择常用的自变量因子(D,胸径平方D2,H,树高和胸径乘积DH)和函数模型对实测数据进行拟合[12],并根据检验结果来确定具有最优适用性的杉木生物量模型,以此找到一种在大范围内可以同时对相同或不同龄组的杉木生物量进行测算且相对通用的模型,从而进一步提高杉木生物量测算的模型精确度以及效率,增强模型的有效性、可操作性和通用性,为今后杉木的经营管理提供依据,同时为后期的深化研究提供理论指导。
1 研究样区
研究样区为天目山林区,位于浙江省杭州市临安区西北部,119°24'~119°28' E,30°18'~30°24' N,面积为4 284 hm2,海拔300~1 506 m。处中亚热带季风气候区南缘,属季风型气候,温暖湿润,光照充足,雨量充沛,土壤肥沃,土壤厚度为50~90 cm,森林植被生长茂盛。自山麓到山顶植被分布规律为:海拔870 m 以下为针阔混交林、常绿阔叶林、毛竹Phyllostachys heterocycla‘Pubescens’林,主要乔木树种有杉木、枫香树Liquidambar formosana等;海拔870~1 100 m 为常绿落叶阔叶混交林,主要乔木林有小叶青冈Cyclobalanopsis myrsinifolia等;海拔1 100 ~1 380 m 为落叶阔叶林,主要乔木林有小叶白辛树Pterostyrax corymbosus等;海拔1 380 m以上为落叶矮林,主要树种为鸡树条Viburnum sargentii等[13]。本研究树种为杉木,样区的海拔位于400~800 m之间。虽然天目山林区禁止采伐,但当地会根据林木生长状况进行更新造林,因此有新林的植入,但面积都比较小。
截至2016年4月,临安区林业用地面积达26.10万hm2,占全市土地总面积的83.28%,森林总蓄积已突破1 000万m3,活立木蓄积达830.01万m3,林木年平均消耗量31.59万m3,森林覆盖率高达76.55%。境内以山地丘陵为主,地形复杂,受地貌影响,立体气候明显;丘陵地区红壤广布,是山区森林生长的主要土壤[14]类型。
2 研究方法
2.1 杉木样本数据采集
研究数据来自临安市2009年森林资源连续清查。2016年4月根据龄组设立了6个(20 m×20 m)标准地,林分密度分别为幼龄林2 700株·hm-2,中龄林2 200株·hm-2,近熟林1 950株·hm-2,1 900株·hm-2,成熟林1 350株·hm-2,1 300 株·hm-2,各龄组及不同林分密度各设一个标准地。每个标准地进行每木检测,以1 cm为径级,分别统计各径级的树高、株数,依此来确定林分胸径的径阶范围,其中,幼龄林的径阶(径阶距为2 cm,上限排除法)6~8 cm,中龄林8~12 cm,近熟林12~16 cm,成熟林16~22 cm。然后在各标准地内从各径级对应的株树较多的杉木中随机抽取一株生长正常的杉木作为样木,共选取23株,并对样木进行记录(表1)。
样木生物量数据的采集均采用称重法获取[15-16]。把树冠分为上下两层,砍下各层的枝条,采集所有树枝上的树叶,对树叶取样称重,编号带回;之后再从不同部位选取树枝,并从树枝的上中下部各截取5 cm小段圆柱进行称重,并编号带回;树干和去皮干的采集是标准木伐倒以后,去除所有枝条,再把主干分段,称量树干的鲜重并编号,然后对编号的树干样本去皮、称鲜、编号带回;树根采用全挖法,分别对主根(含根桩)、侧根及>2 mm的细根取样称重,记录数据,编号带回。
把采集的样品带回实验室,在烘箱中烘干,烘烤过程中,每隔一段时间对样品进行称重,直到两次称量的相对误差小于1%,这时把样品取出,冷却后再称量干重。最终得到相应的生物量实测值。
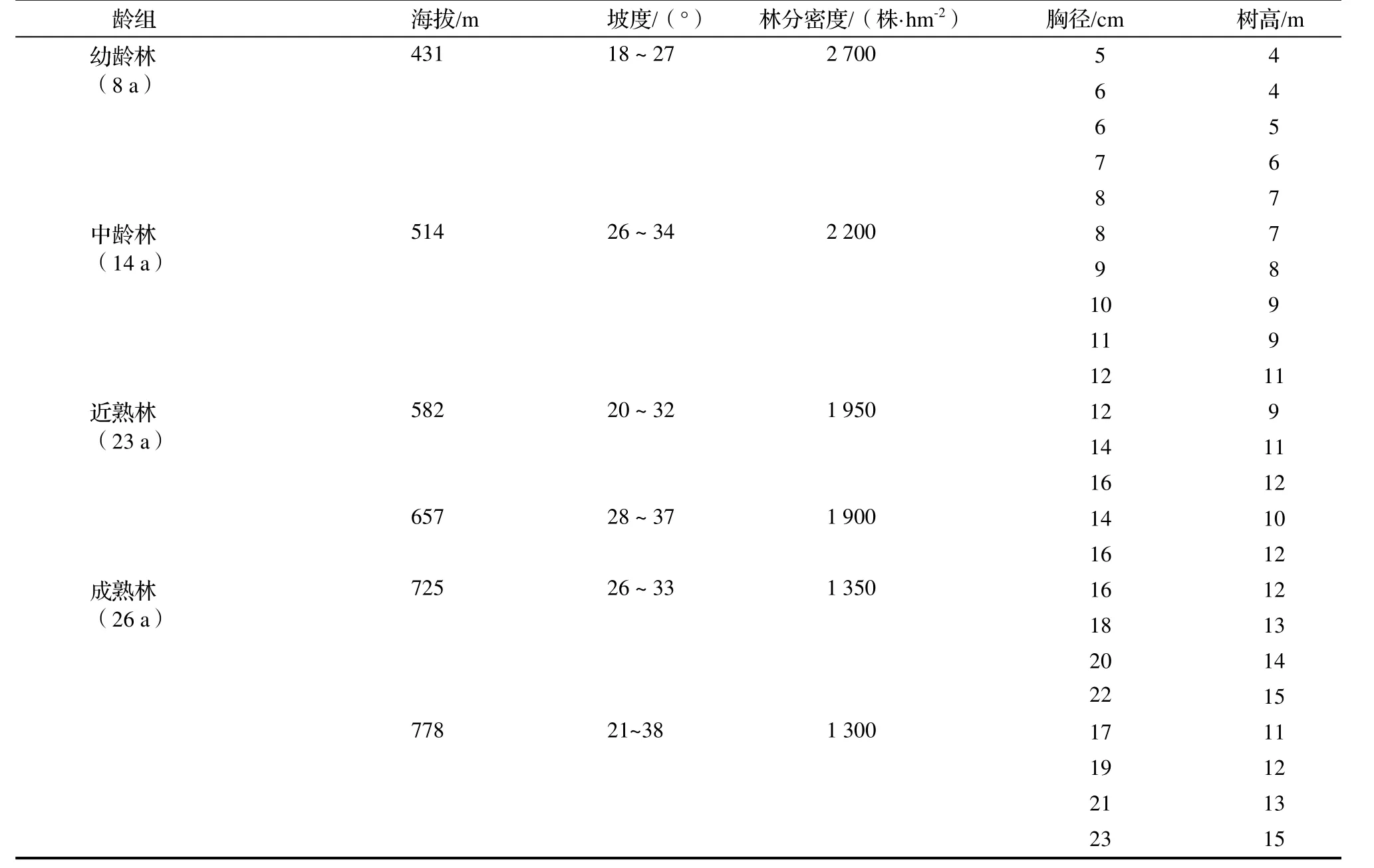
表1 杉木林样木测定数据Table 1 Analytic woods of C. lanceolata
2.2 样本数据的处理
用SPSS 19.0对杉木生物量模型进行拟合,并进行效果和精度的检验。选用6种常用的生物量模型,如表2所示,基于实测生物量和胸径、树高数据,运用表2中的6种模型进行回归分析,最终得到杉木幼龄林、中龄林、近熟林和成熟林的单木各器官和全株生物量估算模型[17-18],并把决定系数(R2)、残差平方和(RSS)作为模型拟合效果评价指标[19],平均精度A作为杉木生物量模型的通用性预测效果的检验指标。

表2 生物量预测模型Table 2 Biomass prediction models

式中,Nsamp指样本容量。A值越大说明模型的预测效果就越好,通常,A值>80%时,就表示模型比较符合实际[2]。
3 结果与分析
研究结果表明,拟合后生物量模型的R2在0.812~0.945,RSS在0.015 9~0.432 0,F检验达到了显著水平(P<0.01),因此以D和DH为自变量的杉木单木生物量模型可用于实际。同时发现幂函数、指数函数和多项式函数模型的拟合效果依次为幂函数优于指数函数优于多项式函数。
通过检验结果的对比,得到了24个效果较好的拟合模型,见表3。
由表3可知,杉木各龄组生物量最优模型均以幂函数模型最优,其中,生物量最优模型以D为自变量的模型有11个,以DH为自变量的模型有13个,均有较好的拟合效果。
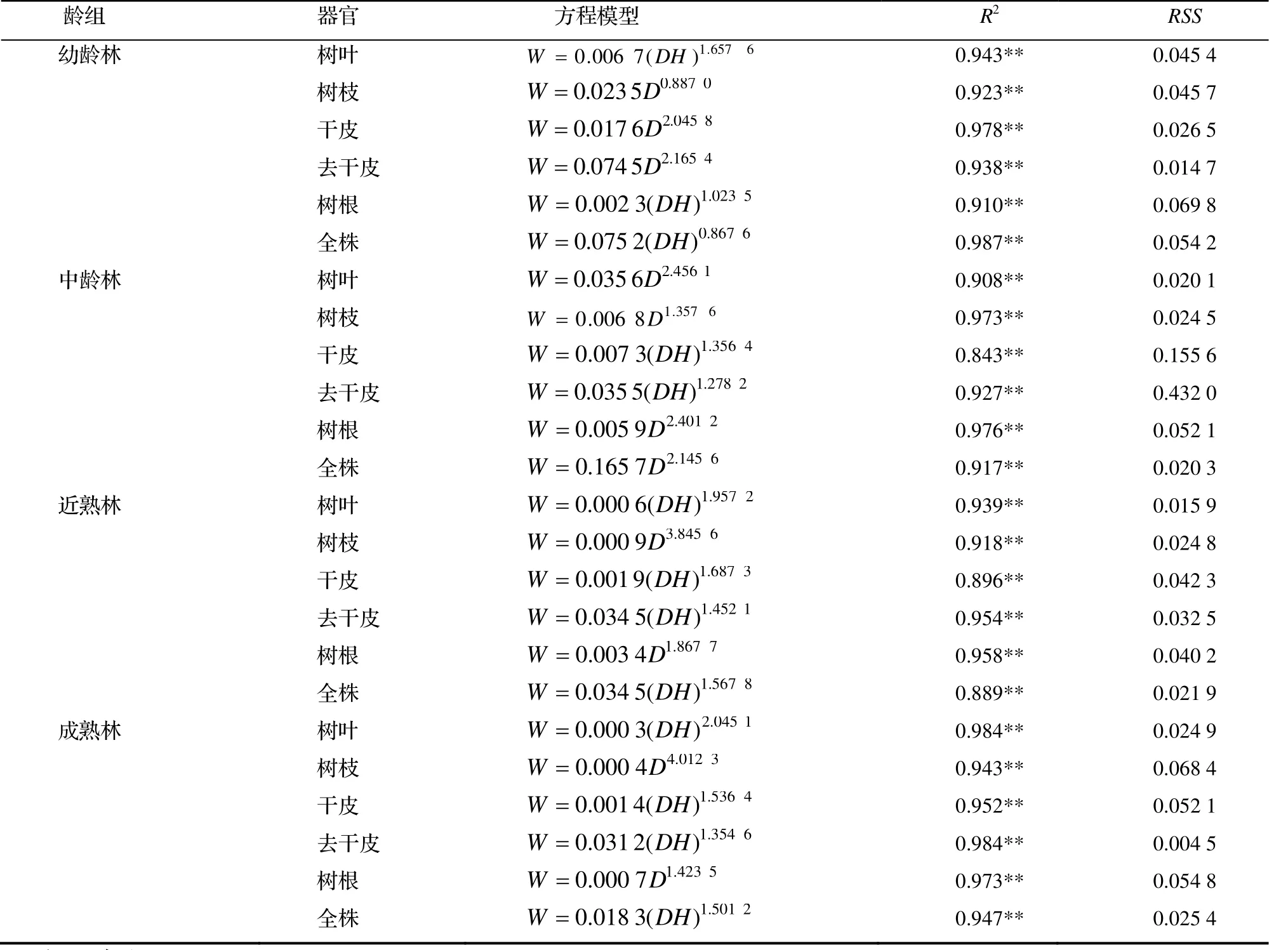
表3 杉木单株生物量估算模型Table 3 Biomass estimation model for individual tree of C. lanceolata
3.1 各龄组杉木生物量最优模型拟合结果分析
由表3可知,杉木幼龄林各器官和全株生物量最优模型的R2为0.910~0.978,RSS为0.014 7~0.069 8。在幼龄林中树叶、树根和全株的生物量模型形式为:W = a(DH)b,从而得出,杉木幼龄林的树叶、树根和全株的生物量主要受DH影响,而且树叶生物量模型和全株生物量模型的拟合效果均优于杉木树根的拟合效果。同理发现杉木树枝、干皮与去干皮的生物量最优模型形式为:W = aDb,表明此阶段杉木树枝、干皮和去干皮的生物量主要受到D的影响,其中去干皮的拟合效果较好,树枝和去干皮的生物量模型的拟合效果次之,但相差不大。
中龄林单木各器官和全株生物量最优模型的R2为0.843~0.976,RSS为0.020 1~0.432 1。其中树叶、树枝、树根和全株的生物量最优模型形式为:W = aDb,说明中龄林杉木的树叶、树枝、树根和全株的生物量主要受到D的影响;干皮和去干皮的生物量最优模型形式为:W = a(DH)b,说明中龄林的单木干皮和去干皮的生物量主要受DH综合影响。
近熟林单木各器官和全株生物量最优模型的R2为0.889~0.958,RSS为0.015 9~0.542 3。其中树枝和树根的生物量最优模型形式为:W = aDb,说明近熟林的杉木中,树枝和树根的生物量主要受到D的影响;树叶、干皮、去干皮和全株的生物量最优模型形式为:W = a(DH)b,说明近熟林的单木干皮和去干皮的生物量主要受DH综合影响。成熟林单木各器官和全株生物量最优模型的R2为0.943~0.984,RSS为0.004 5~0.521 7。其中树叶、干皮、去干皮和全株的生物量最优模型形式为:W = a(DH)b,说明成熟林的杉木中,树叶、干皮、去干皮和全株的生物量主要受到DH的影响;树枝和树根的生物量最优模型形式为:W = aDb,说明成熟林的单木树枝和树根的生物量主要受到D的影响。
以上分析表明,W = a(DH)b和W = aDb均可作为杉木幼龄林、中龄林、近熟林和成熟林的最优模型形式。各龄组内部拟合效果最优的模型分别为幼龄林的全株模型,中龄林、近熟林的树根模型和成熟林的去干皮模型。
3.2 杉木生物量模型的互换通用性
用本研究建立的模型对临安区的杉木生物量进行预测,通过调查数据比较模型的实际效果(表4)。
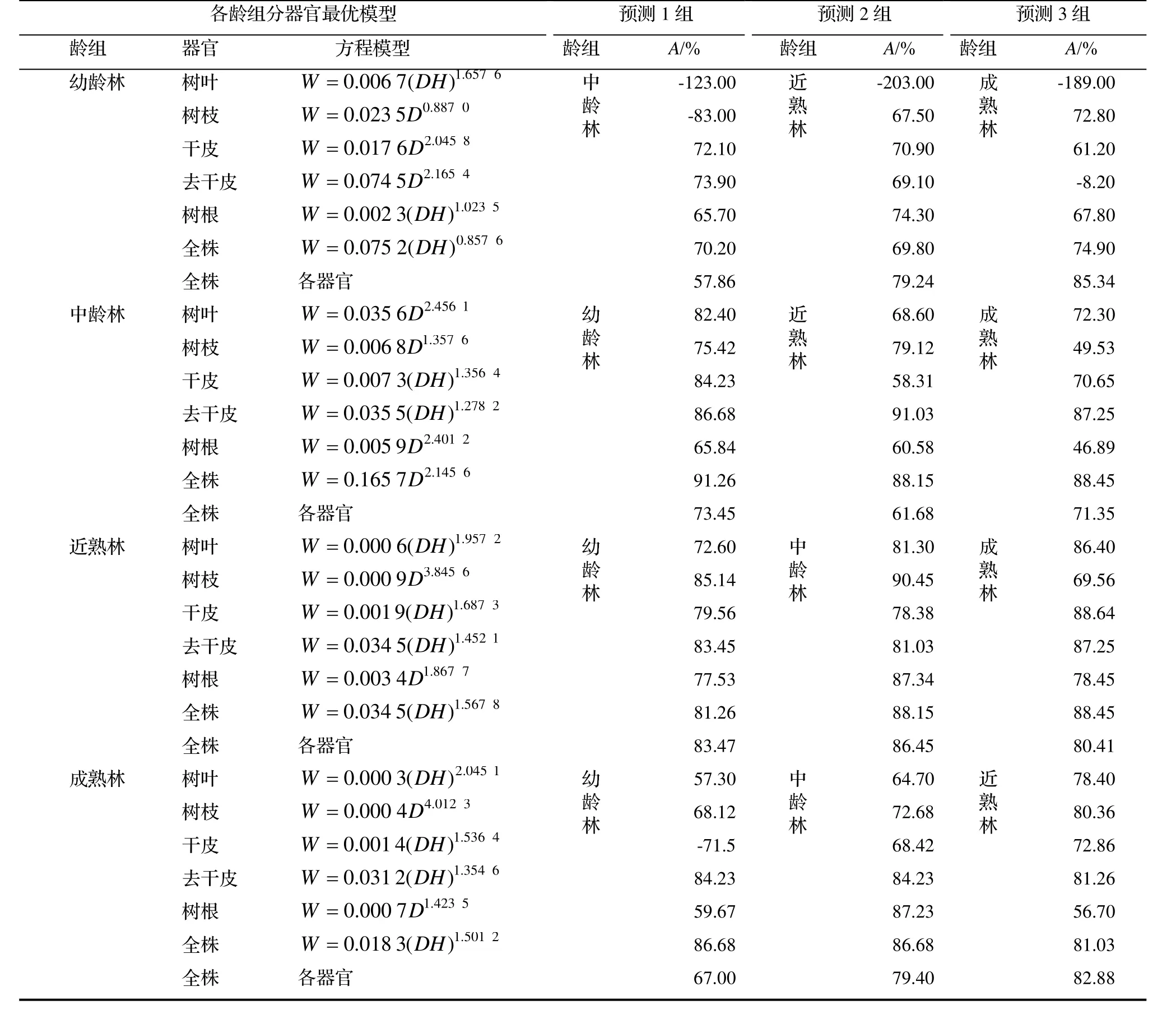
表4 杉木最优模型对各龄组的单木生物量的预测效果Table 4 Prediction of the optimal model for the individual biomass of C. lanceolata with different ages
根据表4可知,幼龄林各生物量的最优模型对中龄林、近熟林和成熟林模型预测的平均A普遍较低,均低于75%,但是幼龄林各器官模型对成熟林全株模型预测的A达到85.34%。由此可知,杉木幼龄林的生物量最优模型不能直接用于中龄林、近熟林和成熟林的全株生物量预测,且各器官模型仅适用于成熟林全株生物量模型的预测,整体的通用性较差。中龄林生物量最优模型对幼龄林的树叶、干皮、去干皮和全株以及近熟林与成熟林的去干皮和全株生物量预测的A均高于80%,说明可以相对精确的预测幼龄林、近熟林和成熟林全株的生物量。同时,杉木中龄林全株生物量最优模型对幼龄林、近熟林和成熟林的全株生物量的预测效果均优于各器官的效果,说明杉木林不分龄组时,中龄林全株生物量模型可以用于幼龄林、近熟林和成熟林全株的生物量测算,且具有较好的通用性。
近熟林生物量最优模型对幼龄林的树枝、去干皮和全株以及中龄林的树叶、树枝、去干皮、树根和全株以及对成熟林的树叶、干皮、去干皮和全株生物量预测的A均大于80%,说明可以相对精确的预测幼龄林、中龄林和成熟林全株的生物量。
成熟林生物量最优模型对幼龄林的去干皮和全株以及中龄林的去干皮、树根和全株以及近熟林的树枝、去干皮和全株生物量预测的A均高于80%,这说明可以相对精确的预测幼龄林、中龄林和近熟林全株的生物量。
综上所述,临安区杉木中龄林、近熟林和成熟林的全株生物量最优模型均可对不分龄组时的杉木全株生物量进行预测,其中以中龄林的通用性效果最好,对各龄组生物量的预测精度均大于88%。因此,中龄林全株生物量模型W = 0.165 7D2.1456可以对杉木生物量进行预测,具有较好的通用性。
4 结论与讨论
受自然条件和林分因子等多方面影响,杉木在不同龄组阶段生物量的主要影响因素不同。幼龄林阶段冠幅偏小,D和H的生长较快,根系生长旺盛[9];中龄林经过多年的自然选择,成活的树木已进入干材阶段,冠幅较大,D,H的生长相对缓慢,根系仍然在不断增长,材积增长迅速;近、成熟林阶段杉木的H逐渐停止生长,材积还在增长,但是日趋缓慢[20]。本研究发现,D和H是幼龄杉木树叶、树根和全株生物量的重要影响因素;中龄杉木的树叶、树枝、树根和全株生物量主要取决于D,而树干主要是受D和H影响;近、成熟杉木的树叶、树干和全株的生物量主要受D和H影响,而树根和树枝主要是受D的影响。
为保证模型的可行性,在拟合模型的检验和评价时选用了R2,RSS和A 3个常用指标,尽可能有效反映预测模型的拟合效果和通用性。最终发现,杉木幼龄林的生物量模型对其他龄组的通用性最差;近熟林和成熟林次之;中龄林的通用性相对较好,可用于临安地区不同龄组的杉木全株生物量的估测。因此,除D外,对小范围杉木生物量进行分器官高精度计算时,还需要考虑到各器官的其他影响因子,但是在野外大范围林区进行杉木总生物量估算时只需考虑D因子。
本研究模型所得生物量结果不像各龄组分器官生物量模型结果那样准确,但预测精度达到80%以上。生物量的计算本身就存在一定的误差,估算模型和自变量从多个简化为一个,误差同样存在,因此在不要求精确计算生物量的前提下,可用于临安区杉木生物量的估算,不仅节省分龄组分器官计算生物量的时间,又可以获得生物量的估算结果,降低了实际操作难度,现实意义明显。同时受样本采集区域和样本数量的限制,建立的杉木生物量分器官的最优模型是以D和DH为自变量的幂函数,模型的选择直接影响生物量的预测效果,所建立的模型可能更适用于临安,因此在估算与临安环境差异较大区域的杉木生物量时,模型的选择还需要进一步研究。
[1] 唐守正,张会儒,胥辉. 相容性生物量模型的建立及其估计方法研究[J]. 林业科学,2000,36(专刊):19-27.
[2] 李燕,张建国,段爱国,等. 杉木人工林生物量估算模型的选择[J]. 应用生态学报,2010,21(12):3036-3046.
[3] 罗天祥. 中国主要森林类型生物生产力格局及其数学模型[D]. 北京:中国科学院研究生院,1996:22-23.
[4] 冯宗炜,陈楚莹,张家武,等. 湖南会同地区马尾松林生物量的测定[J]. 林业科学,1982(02):127-134.
[5] 刘兴良,刘世荣,宿以明,等. 巴郎山川滇高山栎灌丛地上生物量及其对海拔梯度的响应[J]. 林业科学,2006(02):1-7.
[6] 吕晓涛,唐建维,何有才,等. 西双版纳热带季节雨林的生物量及其分配特征[J]. 植物生态学报,2007(01):11-22.
[7] 林开敏,郑郁善,黄祖清,等. 杉木和马尾松幼林生物产量模型研究[J]. 福建林学院学报,1993(04):351-356.
[8] 唐坤银,唐代生. 杉木生物量优化模型研究[J]. 林业调查规划,2010,35(01):47-49,52.
[9] 范仁志. 杉木人工林树根生物量模型的研究[J]. 林业勘察设计,2011(1):67-70.
[10] 田大伦,盘宏华,康文星,等. 第二代杉木人工林生物量的研究[J]. 中南林学院学报,1998(03):14-19.
[11] 赵坤,田大伦. 会同杉木人工林成熟阶段生物量的研究[J]. 中南林学院学报,2000(01):7-13.
[12] 骆期邦,曾伟生,贺东北,等. 立木地上部分生物量模型的建立及其应用研究[J]. 自然资源学报,1999,14(3):271-277.
[13] 王敬,韦新良,徐建,等. 天目山针阔混交林林木空间分布格局特征[J]. 浙江农林大学学报,2014,31(05):668-675.
[14] 韦新良,刘永富. 浙江省森林景观地理概述[J]. 浙江林学院学报,1996,13(3):316-321.
[15] 闫文德,田大伦,焦秀梅. 会同第二代杉木人工林林下植被生物量分布及动态[J]. 林业科学研究,2003,16(3):323-327.
[16] 秦立厚,张茂震,钟世红,等. 森林生物量估算中模型不确定性分析[J/OL]. 生态学报,2017,37(23):1-8.
[17] 惠刚盈,童书振,刘景芳,等. 杉木造林密度试验研究Ⅰ.密度对幼林生物量的影响[J]. 林业科学研究,1988(04):413-417.
[18] 段爱国,张建国,何彩云,等. 杉木人工林生物量变化规律的研究[J]. 林业科学研究,2005,18(2):125-132.
[19] 施文涛,谢昕云,刘西军,等. 安徽大别山区杉木人工林乔木层生物量模型及碳贮量[J]. 长江流域资源与环境,2015,24(5):758-764.
[20] 佟金权. 杉木人工林广义干曲线模型及其生物生产力研究[D]. 北京:中国林业科学研究院,1999:64.
猜你喜欢
江西农业大学学报(2022年1期)2022-02-04 11:51:42
江西农业大学学报(2021年2期)2021-05-16 02:18:24
林业勘查设计(2021年2期)2021-04-08 08:27:42
土壤学报(2020年6期)2021-01-05 13:26:54
农民致富之友(2020年8期)2020-05-11 05:54:38
广东农业科学(2019年2期)2019-04-09 08:32:54
农业与技术(2018年16期)2018-11-28 11:24:08
现代农业科技(2017年10期)2017-07-12 14:14:49
防护林科技(2016年5期)2016-09-05 01:19:50
中国农业信息(2016年17期)2016-02-06 09:50:05
