
应用絮凝基因表达量分析技术评价酵母菌株的絮凝性
2018-06-14 06:45:58郭立芸谢鑫梁云
食品与发酵工业 2018年5期
郭立芸,谢鑫,梁云
(北京燕京啤酒股份有限公司,燕京啤酒酿造技术北京市重点实验室,北京,101300)
酵母絮凝是酵母细胞在多种因素影响下形成可逆的、无形聚集并可以再次扩散到发酵介质中的一种能力的体现。对于啤酒工业来说,酵母细胞的絮凝是相当重要的。
STRATFORD[1]根据糖对酵母絮凝抑制的情况,区分酵母的2种絮凝表型,一种称之为FLO1型,另一种为NewFLO型,这2个表型主要体现在糖抑制特征上,不同的糖类对于2种不同絮凝表型的酵母其抑制作用存在差别,FLO1型受FLO1、FLO5、FLO8等基因调控,NewFLO型受Lg-FLO1基因调控,FLO1型和NewFLO型酵母其絮凝性及其生理性状存在较大差异(图1)。

图1 啤酒酵母的絮凝作用分类Fig.1 The classification of yeast flocculation
酿酒酵母携有5个FLO家族絮凝基因[2-3]:FLO1、FLO5、FLO9、FLO10和FLO11。其中FLO1、FLO5、FLO9和FLO10功能是实现细胞与细胞间的黏附,而FLO11使细胞与基质产生黏附;另外的Lg-FLO1基因主要负责甘露糖、葡萄糖和麦芽糖敏感的NewFLO类型的啤酒酵母菌株絮凝(表1)。

表1 絮凝基因的基因功能Table 1 The function of flocculation gene
絮凝性是酵母细胞本身一种固有的特性,菌株遗传背景的特异性起着决定性作用,其他各种因素作用都是通过影响絮凝基因的表达或者絮凝基因编码蛋白的活性来影响酵母最终的絮凝性能。
荧光定量PCR是近年来发展起来的一项新技术,可用于目标基因的表达、基因突变分析及多态性研究,是一种高效的基因表达量分析技术。该技术是在PCR反应体系中加入荧光基团,利用荧光信号积累实施检测整个PCR进程。它可以从复杂的样品中加测中微量的目标核酸,具有高准确性、高特异性[4-5]。张中保等[6]应用实时荧光定量PCR(real-time quantitative, PCR)技术,研究了玉米中10个水分胁迫诱导基因的相对表达量及表达模式,赵鲜仙[7]以酿酒酵母BY4742基因组DNA为模板, 采用PCR技术扩增获得ADH7启动子、CYC1终止子以及MSN2编码框序列, 采用荧光定量PCR技术检测MSN2基因及其调控代表基因的转录变化水平。
目前针对于酵母絮凝性基因方面的研究并不深入,本研究在前期实验中筛选获得1株弱絮凝性酿酒酵母,在此基础上,通过糖抑制实验确定了弱絮凝性和对照酵母菌株的絮凝表型,采用分子生物学技术测定酵母絮凝基因,从分子层面确定,弱凝聚性酿酒酵母与对照菌株在基因层面的差异性,确定酵母絮凝关键基因。
1 材料与方法
1.1 实验材料
1.1.1 实验菌株
弱凝聚性酵母菌株YJ085及对照菌株均由公司菌种保藏中心提供。
1.1.2 培养基
13°P麦汁培养基(糖化车间生产)、PDA培养基(北京陆桥)、YPD-Broth培养基(上海生工)。
1.1.3 主要试剂
糖类:甘露糖、麦芽糖、葡萄糖。
pH 3.9 50 mmol/L醋酸钠-醋酸-0.1% CaCl2的絮凝缓冲液:称取4.1 g醋酸钠、3 g醋酸和1 g CaCl2溶解后定容至1 L,用醋酸调节pH为3.9。
PCR反应相关试剂:Taq酶、dNTP、TaqReaction Buffer、Loading Buffer、QuantScript RT Kit、Real Master Mix(SYBR Green)(天根生化);Goldview核酸染料;柱型酵母RNA提取试剂盒(上海生工),RNAstore样本保存液。
1.1.4 主要仪器
恒温摇床培养箱(ZWY-C2112B型),上海智城分析仪器制造有限公司;可见光分光光度计(WFJ2100型,Unico);恒温水浴锅,美国polyscience;PCR仪(GeneAmp 9700 型),美国Applied Biosystems公司;荧光定量PCR仪(Light cycler2.0型),瑞士罗氏公司;水平电泳仪(DYY-8B型),北京市六一仪器厂;凝胶成像系统(JY04S-3C型),北京君意东方电泳设备有限公司;生化培养箱(INCUBATOR型),日本SANYO公司;超净工作台,苏州净化设备有限公司。
1.2 酵母絮凝表型的分型
1.2.1 酵母培养
挑取1环实验菌株,于25 ℃液体的麦汁培养基中活化1次,转接至三角瓶中,25 ℃,200 r/min摇床培养48 h。
1.2.2 絮凝水平的测定
参照STRATFORD[1]、张博润[8]、常玉广等[9]采用的分光光度法:取48 h培养物离心收集菌体,用250 mmol/L溶液及无菌水各洗涤2次,60 ℃处理5 min以杀死细胞(为避免代谢旺盛的酵母细胞消耗糖而降低其浓度,该处理不影响细胞的絮凝),然后测定絮凝水平离心收集菌体并悬于絮凝缓冲液中,立刻在600 nm处测OD值,然后细胞悬液转移至三角瓶中,25 ℃,120 r/min振荡2 h至絮凝完成,室温静置30 min,取上清在600 nm 处测OD值。以振荡处理后测定的OD值除以振荡处理前的OD值,再乘以100%(即自由细胞浓度)表示絮凝水平。
1.2.3 酵母菌的絮凝表型分型
分别将不同浓度的甘露糖、麦芽糖、葡萄糖溶于絮凝缓冲液中,浓度分别为0、0.2、0.4、0.6、0.8、1 mol/L,测定絮凝水平。
根据FLO1型酵母菌株絮凝仅受甘露糖抑制,而NewFLO型酵母菌株受多种糖抑制对酵母菌株进行絮凝表型分型。
1.3 酵母絮凝表型基因型的测定
1.3.1 特异性引物设计
针对酵母絮凝基因FLO1、FLO5、FLO8、FLO9、FLO10、FLO11、Lg-FLO1设计特异性引物,经过查阅相关文献,确定并设计引物见表2。
1.3.2 菌株培养
从原始琼脂斜面挑取菌苔1环,使用10 mL 13 °P麦汁培养基活化培养48 h,吸取1 mL转入10 mL 13 °P麦汁培养基活化培养24 h,挑取1环接种PDA培养基划线培养3 d。
1.3.3 酵母DNA的提取
酵母的DNA使用TIANGEN酵母基因组DNA提取试剂盒进行提取,并使用0.8%的琼脂糖凝胶电泳(80 V,30 min)检测所得DNA的质量。

表2 酵母絮凝基因特异性引物Table 2 Specific primers of yeast flocculation gene
1.3.4 PCR反应体系及反应条件
A:反应体系的优化
反应体系:10×Buffer 5 μL,2.5 mmol/L dNTP 1μL,10 μmol/L上游引物1 μL,10 μmol/L下游引物1 μL,Taq酶0.5 μL,模板3 μL,ddH2O 38.5 μL。
反应条件:94 ℃ 5 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,30个循环;72 ℃ 5 min。
1.3.5 酵母絮凝基因分型
对酵母的絮凝基因FLO1、FLO5、FLO8、FLO9、FLO10、FLO11、Lg-FLO1进行扩增,产物进行2%琼脂糖凝胶电泳,若目的基因扩增出现阳性条带,则代表酵母具有该基因,若无目的基因条带,则代表酵母不具有该基因。
1.4 酵母絮凝基因表达量分析
1.4.1 特异性引物设计
针对酵母絮凝基因FLO1、FLO5、FLO8、FLO9、FLO10、FLO11、Lg-FLO1设计特异性引物,并以ACT1管家基因作为内参基因,引物序列见表2。
1.4.2 菌株培养
将酵母菌用接种针从原始琼脂斜面无菌挑取菌苔1环到10 mL YPD-Broth培养基中,25 ℃恒温培养24 h;用移液管无菌移取1 mL菌液到10 mL YPD-Broth培养基中,25 ℃恒温培养24 h;将试管中的10 mL菌液全部转移到装有90 mL YPD-Broth的250 mL三角瓶中,25 ℃恒温培养,在固定时间段取发酵液1 mL,先离心收集菌体,用PBS缓冲液洗1次,再用100 μL PBS缓冲液悬浮细胞,加入2倍体积的RNAstore保存于4 ℃,待测。
1.4.3 酵母RNA提取
使用柱式酵母RNA提取试剂盒(上海生工)进行提取,得到的产物使用0.8%的琼脂糖进行凝胶电泳(120 V,20 min)检测是否成功提取RNA。
1.4.4 絮凝基因表达量分析
使用相对定量荧光PCR的方法进行分析,使用QuantScript RT Kit进行RNA的逆转录,使用Real Master Mix(SYBR Green)进行荧光定量PCR分析。
逆转录体系:10×RT Mix 2 μL,Super pure dNTPs 2 μL,Random(10 μmol/L) 2 μL,Quant Reverse Transcriptase 1 μL,模板RNA 50 ng-2 μg,RNase-Free水补充至20 μL。
逆转录反应条件:37 ℃孵育60 min。
荧光定量PCR反应体系:2.5×Real Master Mix(添加20×SYBR溶液)9 μL,10μmol/L正向引物 0.5 μL,10 μmol/L反向引物0.5 μL,DNA模板10~100 ng,ddH2O 补充至20 μL。
荧光定量PCR反应条件:95 ℃ 5 min;94 ℃ 30 s,55 ℃ 30 s(结束采集荧光信号),68 ℃ 30 s,45个循环。
按照反应体系及条件对各絮凝基因与内参基因进行荧光定量PCR,使用所得Cp值换算比率,根据比率的大小即可得出基因表达量变化的差异,按公式(1)计算:
比率=2-ΔΔCp
(1)
(ΔCp=Cp目的基因-Cp内参基因,ΔΔCp=ΔCp目的时间点-ΔCp对照时间点,用于生长过程中絮凝基因的表达变化分析)
根据得出的比率值,得知酵母絮凝基因的表达量差异,比率越高说明表达量越高,比率越低说明表达量越低。
1.4.5 酵母生长过程中的絮凝基因表达量变化
考察筛选菌株与对照菌株在发酵过程(YPD-Broth)中的基因表达量变化,每6 h取样1次(摇匀),进行荧光定量PCR分析絮凝基因表达量,同时跟踪各时间点的酵母数、出芽率、死亡率。
2 结果与讨论
2.1 酵母絮凝表型分型实验结果分析
2.1.1 甘露糖对酵母菌株絮凝性的影响
在0.2、0.4、0.6、0.8、1.0 mol/L不同浓度的甘露糖作用下,对筛选菌株YJ085及对照菌株进行絮凝水平的测定,其絮凝性见图2。

图2 甘露糖对酵母菌株絮凝性的影响Fig.2 The effect of mannose on the flocculation of yeast strains
由图2可知,YJ085菌株在0.2 mol/L甘露糖作用下,其絮凝抑制率达到50%左右,随着糖浓度的增加,其絮凝抑制作用略有增加。对照菌株在0.2 mol/L低浓度甘露糖作用下,其絮凝抑制率约为80%,浓度为0.6 mol/L时,对照菌株基本完全抑制絮凝,甘露糖对弱凝聚性酵母菌株YJ085的絮凝抑制性要低于对照酵母菌株。
2.1.2 麦芽糖对酵母菌株絮凝性的影响
在0.2、0.4、0.6、0.8、1.0 mol/L不同浓度的麦芽糖作用下,对筛选菌株YJ085及对照菌株进行絮凝水平的测定,其絮凝性见图3。
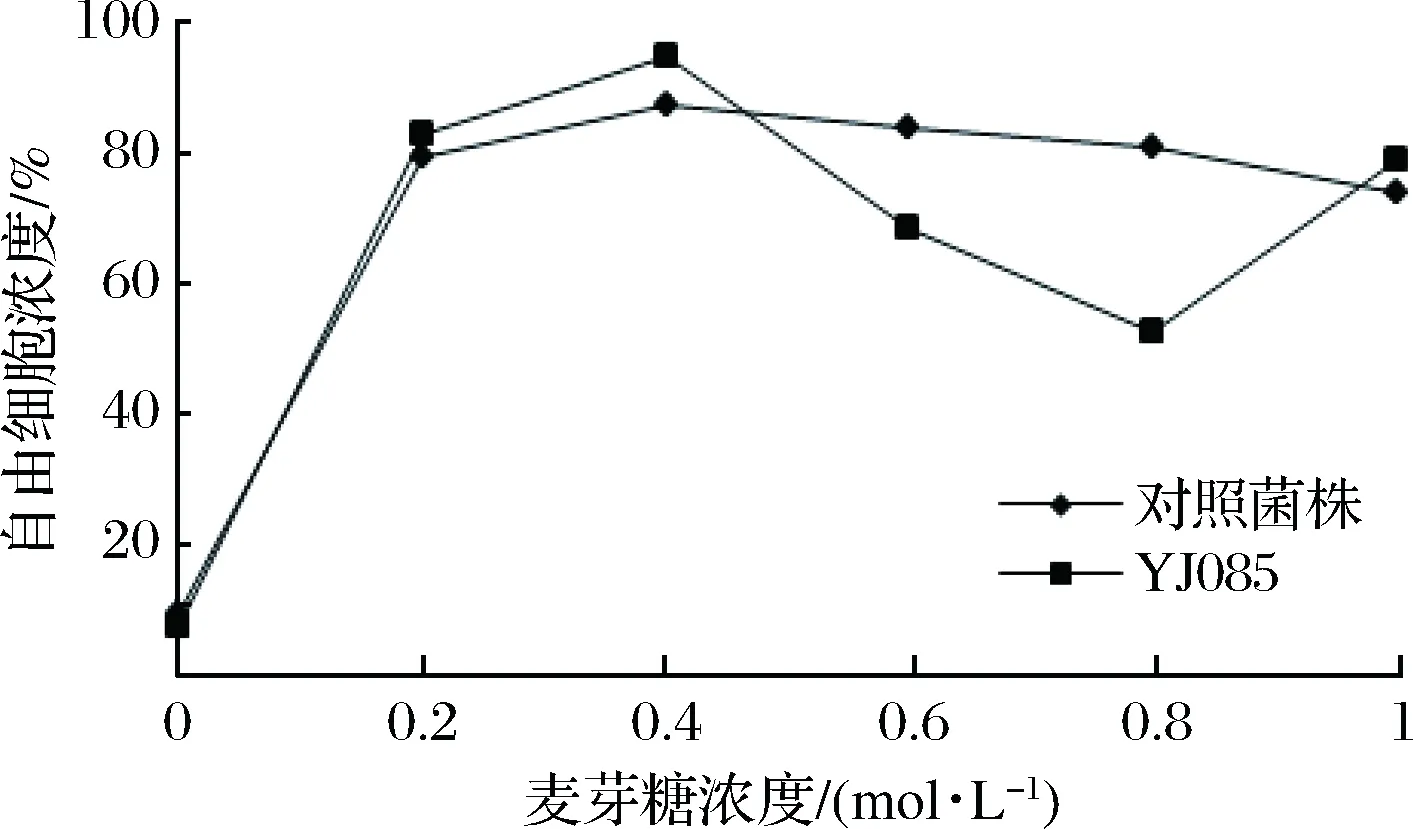
图3 麦芽糖对酵母菌株絮凝性的影响Fig.3 The effect of maltose on the flocculation of yeast strains
由图3可知,YJ085与对照菌株在麦芽糖浓度仅为0.2 mol/L时,絮凝抑制率高达到80%左右,随着糖浓度的增加,絮凝抑制率基本保持在该水平。与对照酵母菌株相比,麦芽糖对弱凝聚性酵母菌株YJ085的絮凝抑制无显著差异。
2.1.3 葡萄糖对酵母菌株絮凝性的影响
在0.2、0.4、0.6、0.8、1.0 mol/L不同浓度的葡萄糖作用下,对筛选菌株YJ085及对照菌株进行絮凝水平的测定,其絮凝性见图4。
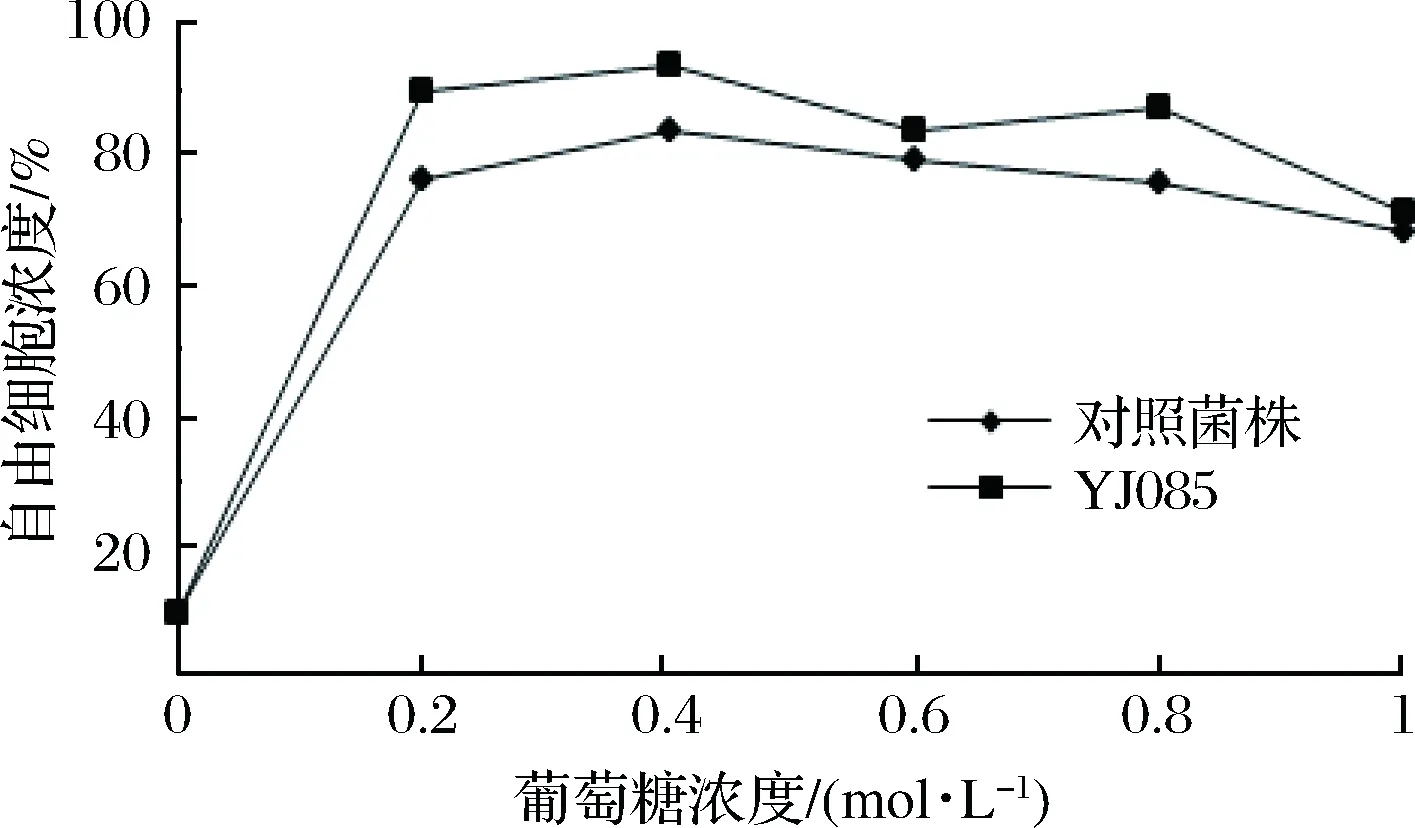
图4 葡萄糖对酵母菌株絮凝性的影响Fig.4 The effect of glucose on the flocculation of yeast strains
由图4可知,YJ085与对照菌株在葡萄糖浓度仅为0.2 mol/L时,絮凝抑制率达80%左右,随着糖浓度的增加,基本维持在该水平。与对照菌株相比,葡萄糖对弱凝聚性酵母菌株YJ085的絮凝抑制要高于对照酵母菌株。
虽然从絮凝表型上可分为2类,但不同糖类对酵母菌株絮凝抑制效果不同,具体分析结果见表3。

表3 酵母菌株的分型Table 3 The flocculation phenotype of yeast strains
按照根据FLO1型酵母菌株絮凝仅受甘露糖抑制,而NewFLO型酵母菌株受多种糖抑制对酵母菌株进行絮凝表型分型。YJ085与对照菌株均为不仅受甘露糖抑制,还受麦芽糖、葡萄糖的抑制,絮凝表型均为NewFLO型。
与对照相比,弱凝聚性酵母YJ085在甘露糖作用下絮凝性抑制低于对照酵母菌株。
2.2 酵母絮凝表型基因型的测定
对筛选获得的凝聚性酿酒酵母菌株YJ085与对照菌株进行絮凝基因分型,验证其絮凝表型的同时确定各絮凝基因的基因型见图5和图6。
从电泳图可以看出,筛选弱凝聚性菌株YJ085与对照菌株絮凝基因存在状况均一致,YJ085和对照酵母菌株中菌存在FLO1、FLO5、FLO8、FLO9、FLO11、Lg-FLO1基因,菌株的FLO10基因均不存在,均为NewFLO型。

图5 对照菌株絮凝基因分型Fig.5 The flocculation genotype of the control strain

图6 弱凝聚性酵母菌株YJ085基因分型Fig.6 The flocculation genotype of the YJ085 strain
2.3 酵母絮凝基因表达量分析
2.3.1 筛选菌株在生长过程中絮凝基因表达量的变化
以筛选菌株YJ085为研究对象,考察其在发酵过程(YPD-Broth)中的基因表达量变化,同时测定对照菌株基因表达量进行比较,每6 h,摇匀取样,进行荧光定量PCR分析絮凝基因表达量,同时跟踪各时间点的酵母数、出芽率、死亡率。在0~72 h发酵过程中,筛选菌株YJ085及对照菌株均在24 h达到高峰期,随后呈小幅下降直至基本稳定的趋势,出芽率均由发酵初期的13%~18%至发酵后期稳定在8%左右,死亡率在发酵过程中均维持在2%左右,生长曲线见图7。

表4 酵母菌株絮凝基因分型Table 4 The flocculation genotype of the yeast strains

图7 筛选菌株YJ085的生长曲线Fig.7 The growth curve of YJ085 strain
对不同时间点的发酵液取样,提取酵母RNA进行荧光定量PCR分析,筛选菌株YJ085与对照菌株在生长过程中絮凝基因的表达量变化见图8。

图8 菌株生长过程中絮凝基因的表达量变化Fig.8 The expression of flocculation gene in the growth of strain
从图8可以看出,筛选菌株YJ085各絮凝基因表达量显著低于对照菌株,筛选菌株YJ085的絮凝主要受FLO絮凝基因家族中的FLO5及Lg-FLO1基因调控,FLO1、FLO11表达量在发酵过程始终维持在较低表达量,FLO10基因不表达;在接种初期随着酵母的大量繁殖,FLO家族絮凝基因(FLO5、FLO8、FLO9、FLO11)表达出现抑制,随着酵母数在24 h左右达到高峰期,絮凝基因FLO5、FLO8、FLO9及Lg-FLO1表达量达到峰值;随后酵母数开始下降,絮凝基因的表达量也出现大幅下降;至36 h左右酵母数基本稳定,FLO8、FLO9基因开始维持在较低表达量,并呈现小幅波动趋势,而FLO5基因的表达量则呈现较大波动,并在42 h及54 h左右出现峰值,FLO11基因发酵后期表达量有小幅上涨趋势,Lg-FLO1基因表达量在48 h出现小幅升高后便稳定在较低水平。表明无论是筛选菌株或对照菌株,在下面酵母的发酵过程中,絮凝基因的表达量与酵母增殖数量呈负相关。
2.3.2 酵母菌株絮凝基因的表达量差异
荧光定量PCR分析筛选菌株YJ085与对照菌株在发酵过程酵母增值高峰期(24 h)的絮凝基因表达量,分析结果见图9。
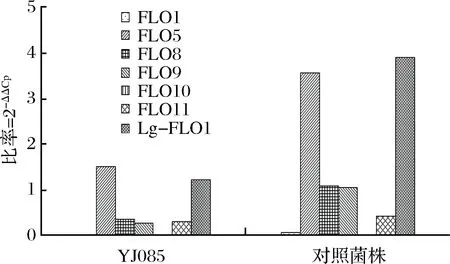
图9 不同酵母增殖高峰期的絮凝基因表达量Fig.9 The expression of flocculation gene in the proliferation peak of different yeast strains
经絮凝基因表达量分析发现,弱凝聚性酿酒酵母菌株YJ085絮凝基因FLO1、FLO5、FLO8、FLO10、FLO11、Lg-FLO1表达量均小于对照菌株,尤其是FLO5和Lg-FLO1絮凝基因表达量显著低于对照菌株。与对照菌株相比,YJ085絮凝家族基因整体表达量降低了63.98%,絮凝基因表达量的下降导致其凝聚性变化,从基因层面阐述了弱凝聚性酵母絮凝水平低于对照菌株的分子机制。
3 结论
通过甘露糖、麦芽糖、葡萄糖作用,确定弱凝聚性酿酒酵母絮凝表型为NewFLO型,与对照菌株相比,受甘露糖絮凝抑制更为明显。
通过PCR技术对筛选弱凝聚性酵母菌株和对照菌株的基因分型,确定YJ085和对照菌株中均存在FLO5、FLO8、FLO9、FLO11、Lg-FLO1基因,菌株的FLO10基因均不存在,均为NewFLO型。
基因表达分析方法——荧光定量PCR方法的测定结果表明,酿酒酵母发酵过程中,絮凝基因的表达量与酵母增殖数量呈负相关。通过对不同时间点的发酵液进行取样,提取酵母RNA分析得出YJ085与对照菌株在生长过程中絮凝基因的表达量变化主要受FLO絮凝基因家族中的FLO5及Lg-FLO1基因调控,FLO1、FLO11表达量在发酵过程始终维持在较低表达量,FLO10基因不表达,经絮凝基因表达量分析发现,YJ085各絮凝基因表达量均小于对照菌株,絮凝基因表达量的变化导致其凝聚性变化,降低了63.98%,从基因层面阐述了菌株凝聚性弱的原因。
[1] STRATFORD M, ASSINDER S.Yeast flocculation: Flo1 and NewFlophenotypes and receptor structure[J].Yeast, 1991, 7(6): 559-574.
[2] BARNEY M C, JANSEN G P, HELBERT J R.Use of genetic transformation for the introduction of flocculence into yeast[J].Journal of the American Society of Brewing Chemists, 1980, 102(4):S17-22.
[3] RUSSELL I, STEWART G G, READER H P, et al.Revised nomenclature of genes that control yeast flocculation[J].Journal of the Institute of Brewing, 1980, 86(3): 120-121.
[4] LIVAK K J, SCHMITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2(-ΔΔCp)Method[J].Methods, 2001, 25(4):402-408.
[5] MIKAEL K A, JOSÉ M A B, MARTIN B A, et al.The real-time polymerase chain reaction[J].Molecular Aspects of Medicine, 2006, 27(27):95-125.
[6] 张中保, 李会勇, 石云素,等.应用实时荧光定量PCR技术分析玉米水分胁迫诱导基因的表达模式[J].植物遗传资源学报, 2007, 8(4):421-425.
[7] 赵鲜仙, 周玚, 张思伟,等.ADH7启动子精细调控表达MSN2酿酒酵母菌株对糠醛耐受的研究[J].微生物学通报, 2015, 42(10):1 903-1 911.
[8] 张博润, 陈蔚, 铁翠娟, 等.酵母菌絮凝的分型及其生理生化特性的研究[J].微生物学报, 1999, 39(6): 527-532.
[9] 常玉广, 夏四清,马放,等.絮凝微生物 F2 的絮凝表型分析[J].同济大学学报: 自然科学版, 2009, 37(6): 801-804.
[10] GOVENDER P, DOMINGO J L, BESTER M C, et al.Controlled expression of the dominant flocculation genesFLO1,FLO5, andFLO11 inSaccharomycescerevisiae[J].Applied & Environmental Microbiology, 2008, 74(19):6 041-6 052.
[11] TOFALO R, PERPETUINI G, DI G P, et al.Genetic diversity ofFLO1 andFLO5 genes in wine flocculentSaccharomycescerevisiaestrains[J].International Journal of Food Microbiology, 2014, 191(2):45-52.
猜你喜欢
军事文摘·科学少年(2021年1期)2021-02-04 08:03:45
现代园艺(2017年21期)2018-01-03 06:41:32
中国调味品(2017年2期)2017-03-20 16:18:25
创新作文(小学版)(2016年16期)2016-11-11 05:47:54
现代检验医学杂志(2016年5期)2016-08-20 03:17:04
中国康复理论与实践(2015年10期)2015-12-24 05:42:44
中国科技信息(2015年2期)2015-11-16 08:18:32
医学研究杂志(2015年5期)2015-06-10 06:43:26
阅读与作文(小学低年级版)(2015年8期)2015-05-30 10:48:04
现代检验医学杂志(2015年5期)2015-02-06 01:42:20
