
蔗糖、硼、钙离子和γ-氨基丁酸对烟草花粉萌发的影响
2018-06-13 06:40周小利杨诗怡陈志芸廖菊够吕立堂陈穗云
生物学杂志 2018年3期
周小利, 杨诗怡, 陈志芸, 廖菊够, 顾 菁, 吕立堂, 陈穗云
(1.云南大学 云南省高校植物病虫害生物防控工程研究中心, 昆明 650091; 2.贵州大学 农业生物工程研究院山地植物资源保护与种质创新省部共建教育部重点实验室,贵阳 550025; 3.贵州大学 茶学院, 贵阳 550025)
花粉是高等开花植物的重要雄性生殖结构[1],花粉的正常萌发和花粉管生长是保证精子细胞顺利到达胚囊,并实现受精作用的前提,在植物生殖繁育过程中有极其重要的作用[2]。花粉管的快速生长使其穿入雌蕊组织输送精子完成受精作用[3]。花粉管生长具有极性,在其进行极性生长时,不仅需要花粉粒中储存的物质,还需从外界吸收糖分、硼酸和离子等物质,以提供花粉管正常生长和合成新的细胞壁结构。
糖类对植物花粉萌发及花粉管生长非常重要,蔗糖能为花粉的萌发提供能源物质,并且能够维持花粉与培养液之间渗透平衡,防止花粉和花粉管的损坏和破裂[4]。大多数植物花粉离体萌发的蔗糖浓度在10%~20%之间[5],如培养基中蔗糖浓度过低,提供花粉萌发的基础营养不够,花粉萌发就会受到抑制;但培养基中蔗糖浓度过高,则会造成花粉细胞原生质体脱水,抑制花粉萌发[6]。高质量分数蔗糖会抑制花粉萌发及花粉管生长,而蔗糖是光合作用主要产物,广泛分布于植物体内,它是植物储藏、积累和运输糖分主要形式,用蔗糖来培养花粉比其他多糖效果更明显,所以是最适合作为花粉萌发与花粉管生长的培养基[7]。硼酸在花粉管生长过程中也发挥了重要作用,据报道,细胞中98%的硼位于细胞壁,硼酸与果胶类物质RG2Ⅱ(鼠李半乳糖醛酸2Ⅱ)结合,使酸性果胶酯化[5]。花粉管生长过程中,酯化果胶和酸性果胶集中在花粉管的不同部位,其中酸性果胶在整个花粉管中积累,而酯化果胶主要集中在花粉管顶端,两者共同作用增强细胞壁机械强度[8-9],调节细胞壁的结构和性质,同时也阻止酚类物质的积累,更好地促进花粉管的生长[10];另外,硼酸能够使花粉对糖的吸收、转运和代谢增加,进而形成糖硼复合体,增加氧的吸收[11]。一般情况下,花粉中贮存的硼含量不足,当花粉在柱头上萌发时,柱头会渗出硼离子补偿花粉,满足花粉萌发的最适硼离子浓度[12-13]。Ca2+是细胞内的调节信使[14],具有调节胞内酶活性、促进细胞壁合成和调节细胞膨压等多重作用;Ca2+能影响细胞骨架的组装、分泌小泡的运输和融合、花粉管生长方向等[15]。通常花粉管的顶端生长要求细胞中游离钙的动态平衡,如平衡被打破,花粉管生长受抑制。适宜的γ-氨基丁酸(GABA)对烟草花粉的萌发也起着重要的作用。适宜浓度的GABA促进花粉的萌发和花粉管的生长,高浓度的GABA抑制花粉管的萌发[16]。研究表明,GABA能结合到细胞膜上,可能通过GABA-B受体来调节花粉管Ca2+的通道,导致Ca2+内流并调节花粉管的极性生长[17]。非损伤测微技术证明了外源GABA会使花粉管Ca2+内流,从而使花粉管内Ca2+浓度增加[18]。因此,GABA和Ca2+在影响花粉萌发和花粉管生长过程中有相互促进和协调的作用。
烟草花粉萌发和花粉管生长对烟草杂交育种过程至关重要。目前,有关烟草花粉的研究主要集中在花粉活力的测定[19]、花粉形态学[20]和花粉超微结构[21]等方面。课题组前期开展了栽培烟草K326(N.tabacumL.cv.K326)同野生烟草种N.repanda、N.stocktonii(残波烟草,斯托克通氏烟草,同属于残波烟草组)、N.alata(花烟草,碧冬烟亚属)[22]种间杂交亲和性及其生殖生理学基础研究,表明N.tabacumL.cv.K32×N.alata表现为花柱杂交不亲和障碍,而N.tabacumL.cv.K32×N.repanda、N.stocktonii则表现为柱头杂交不亲和[23-25]。本研究则主要对这几个种花粉离体萌发的条件进行探索,确定了不同烟草种花粉离体萌发最适的蔗糖、硼酸、钙离子和GABA浓度,以期丰富烟草花粉离体萌发影响因素的探究,并为研究上述烟草种间杂交亲和性差异的原因提供指导。
1 材料和方法
1.1 材料的收集与处理
供试材料分别是花烟草(N.alata)、栽培烟草K326(N.tabacumL.cv.K326)、斯托克通氏(N.stocktonii)和残波烟草(N.repanda),于2015年10月播种于云南大学生物馆东楼实验基地,3—5月后,取始开期的花,用镊子轻轻夹取雄蕊花药放进1.5 mL eppendorf离心管中,室温放置2 d,待其开裂后收集花粉。用一层滤纸包着大枪头尾端,然后将滤纸包着的枪头插进真空泵上的橡胶管中,打开抽真空按钮,将枪口尖的一端放入收集了花粉的离心管中,吸附花粉,花粉被抽到枪头中和滤纸上,将枪头中和滤纸上的花粉收集在一个新的1.5 mL Eppendorf离心管中,而花粉粒壳不会穿过枪头尖端,所以仍保留在原来的离心管中,通过这种方式吸附过滤后得到较纯净的花粉,干燥后于4℃下保存7 d待用。
1.2 实验方法
1.2.1 烟草花粉离体萌发培养基的设置
根据研究报道,本实验研究设置了4种不同的蔗糖浓度:8%、10%、12%及16%,5种不同的H3BO3浓度:0.03、0.05、0.1、0.2及0.3 g/L,7种不同的CaCl2·2H2O浓度:0.02、0.03、0.04、0.05、0.06、0.08和0.1 g/L,4种GABA浓度:0.7、1、3和5 mmol/L。每组不同浓度的培养基重复3次。
首先,仅用4种蔗糖浓度(8%、10%、12%和16%)的单因素培养基离体培养烟草花粉,确定每个烟草种花粉萌发的最适培养基;在此基础上,用最适蔗糖浓度+不同浓度的硼酸(0.03、0.05、0.1、0.2和0.3 g/L)确定每个种最适的蔗糖+硼酸培养基;之后,以最适的蔗糖+硼酸培养基加入不同浓度的Ca2+(0.02、0.03、0.04、0.05、0.06、0.08和0.1 g/L),确定每个种最适的蔗糖+硼酸+Ca2+培养基,并以该培养基为基础,寻找各个种最适的蔗糖+硼酸+Ca2++GABA培养基。
1.2.2 烟草花粉离体培养
用移液枪取200 μL的液体培养基于干净的96孔板中,将保存在1.5 mL的离心管中的花粉加入蒸馏水混匀后,然后用移液枪分别向每个孔中加入40 μL的花粉混匀液,用枪吹打混匀。将96孔板用湿润的滤纸遮盖,放入28℃、黑暗环境的恒温烘箱中培养3 h。
1.2.3 镜检
花粉在28℃黑暗恒温烘箱中培养3 h后,用移液枪从96孔板吸取花粉培养液置于载玻片上,轻轻盖上盖玻片,在Olympus BX51光学显微镜下观察花粉萌发状况并统计花粉萌发率[萌发率(%)=萌发的花粉数/花粉总数×100%],用测微尺测量花粉管的长度(如花粉管长度≥花粉直径为萌发,花粉管长度<花粉直径为未萌发)。每个样本统计150粒以上花粉,重复3次。
2 结果与分析
2.1 蔗糖浓度对烟草花粉萌发的影响
统计萌发3 h的烟草花粉萌发率,结果(图1)显示,蔗糖浓度在10%和12%时,花粉萌发率较高。N.alata和N.tabacumL.cv.K326花粉最适蔗糖浓度为10%,萌发率分别为46%和43% (图1),N.repanda和N.stocktonii在蔗糖浓度为12%时,花粉的萌发率分别达到最高值39%和36%。随着蔗糖浓度逐渐升高,烟草花粉的萌发逐渐下降,在蔗糖浓度高于16%时,花粉萌发率低于25%。而在蔗糖浓度低于8%时,仅有N.alata花粉萌发率达到了25%,其他3个烟草种的花粉萌发率低于25%。
2.2 硼酸对烟草花粉萌发的影响
在最适蔗糖浓度的培养基中,加入不同浓度的硼酸,花粉的萌发率提高。由图2可见,硼酸浓度达到0.05 g/L时,花粉萌发率最高,N.alata为72%,N.tabacumL.cv.K326为71%,N.stocktonii和N.repanda都为65%。硼酸浓度超过0.05 g/L 时,4个烟草种的花粉萌发率都呈现下降趋势。
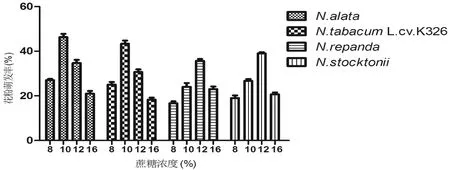
图1 蔗糖浓度对烟草花粉萌发生长的影响Fig 1 The effect of sucrose concentration on tobacco pollen germination

图2 硼酸浓度对烟草花粉萌发生长的影响Fig 2 The effect of boric acid concentration on tobacco pollen germination
2.3 钙离子对烟草花粉萌发的影响
在最适的蔗糖硼酸培养基中添加不同浓度的Ca2+,结果表明在一定范围内,随着Ca2+浓度的增加,烟草花粉的萌发率上升。由图3可见,当Ca2+浓度增加到0.05 g/L 时,花粉萌发率最高,N.alata和N.tabacumL.cv.K326萌发率分别为89%和90%,N.repanda和N.stocktonii花粉萌发率分别达到83%和86%。在最适0.05 g/L Ca2+浓度时,N.alata花粉萌发率比蔗糖硼酸培养基中萌发率增加20%,N.tabacumL.cv.K326和N.stocktonii增加19%,N.repanda增加21%。当Ca2+浓度≥0.1 g/L,N.repanda和N.Stocktonii、N.alata和N.tabacumL.cv.K326的花粉管生长受到明显抑制。
2.4 GABA对烟草花粉萌发的影响
在最适浓度的蔗糖+硼酸+Ca2+培养基中,加入不同浓度GABA培养4个烟草种的花粉。从图4可看出,培养基中加入0.7 mmol/L GABA后,供试的N.tabacumL.cv.K326、N.repanda和N.stocktonii花粉的萌发率分别为90%、83%和88%,GABA浓度达到1 mmol/L 时,3个种花粉萌发率分别为92%、 88%和89%(不加GABA时花粉萌发率分别为89%、83%和86%)。显著性差异分析表明:3个烟草种在加入0.7 mmol/L GABA和1 mmol/L GABA时,与对照组CK3(10%蔗糖+0.05 g/L硼酸+0.05 g/L Ca2+,图4)相比,花粉的萌发率差异不明显;而当GABA浓度为3 mmol/L时,GABA对3个烟草种的花粉萌发率起抑制作用;值得注意的是,N.alata花粉加入不同浓度的GABA后,花粉萌发率降低,加入0.7 mmol/L GABA 时,花粉萌发率为81%,加入1 mmol/L GABA 时,花粉萌发率为84%。显著性差异分析表明,这两种GABA浓度下,N.alata花粉的萌发率差异不明显,但均较对照组(图5,CK3,萌发率为89%)萌发率降低,伴随GABA浓度升高,其对花粉萌发率的抑制作用更明显。
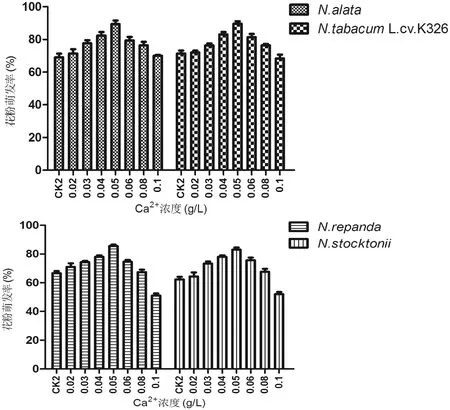
图3 Ca2+对烟草花粉萌发的影响Fig 3 The effect of calcium concentration on tobacco pollen germinate
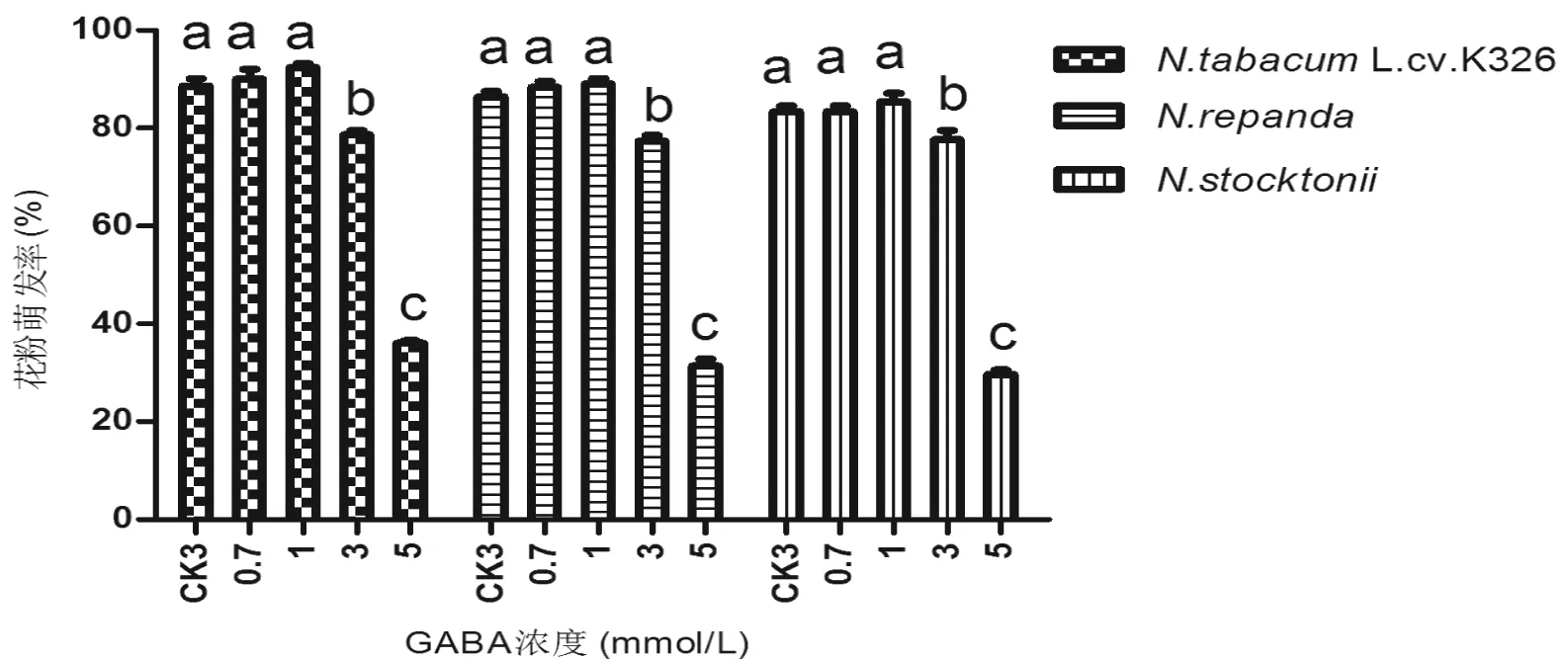
图4 GABA对烟草种花粉萌发的影响Fig 4 The effect of GABA concentration on tobacco pollen germinate
根据上述研究,本实验在不含Ca2+的培养基中检测了GABA对N.alata花粉离体萌发的影响,结果(如图6)表明:10%蔗糖+0.05 g/L 硼酸培养基中,浓度低于1 mmol/L 的GABA促进N.alata花粉离体萌发,最适GABA浓度为1 mmol/L,高于该浓度则抑制花粉萌发。在10%蔗糖+0.05 g/L 硼酸+0.05 g/L Ca2+培养基(图3)和10%蔗糖+0.05 g/L硼酸+1 mmol/L GABA培养基(图6)中,N.alata花粉萌发率分别为89%和91%,二者差异不显著,表明最适浓度的Ca2+和GABA促进花粉萌发的作用相当。
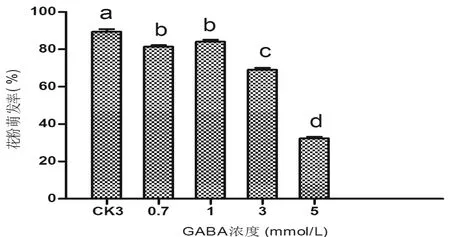
图5 GABA对烟草种N.alata花粉萌发的影响Fig 5 The effect of GABA concentration on N.alata pollen germinate
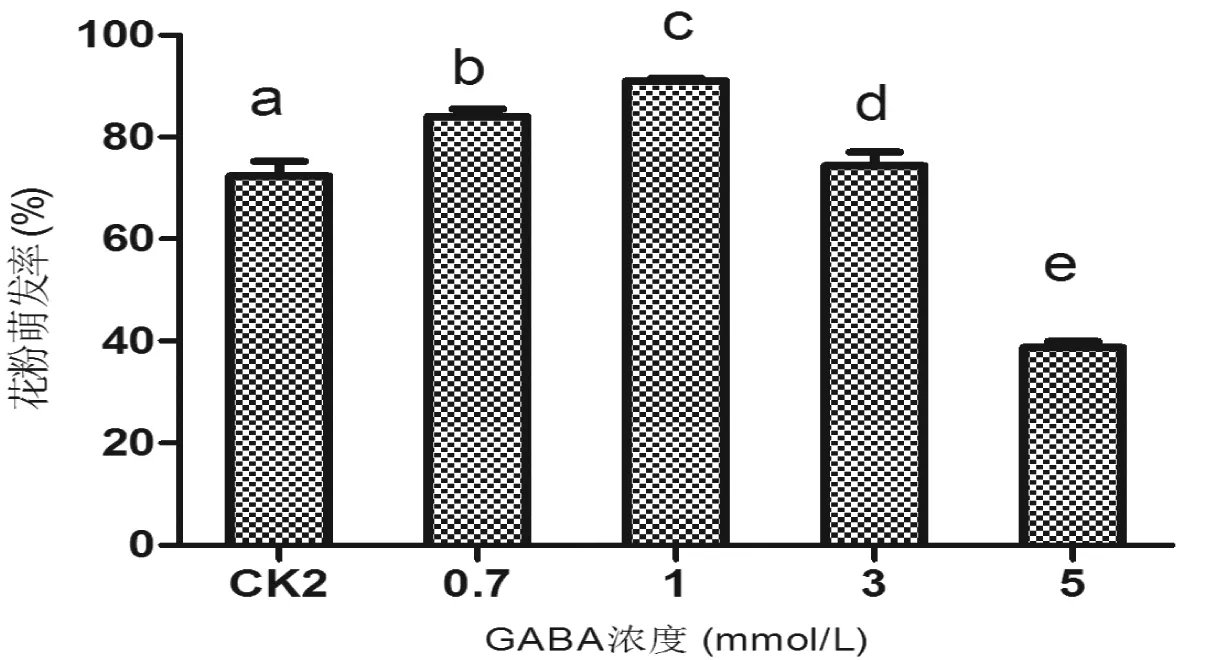
图6 GABA对烟草种N.alata花粉萌发的影响(不含Ca2+)Fig 6 The effect of GABA concentration on N.alata pollen germination (without Ca2+)
3 讨论
花粉离体萌发需要碳源(如蔗糖)、硼酸和矿物质(如钙离子)。蔗糖为花粉萌发及花粉管壁的合成提供能源物质,而硼酸可以加强花粉离体萌发时对糖的吸收和代谢,钙在花粉管顶端物质运输和引导花粉管生长等过程中具有重要作用[26-31]。GABA则可能通过调控胞内钙离子引导和调控花粉管生长[32]。已有研究表明,蔗糖对花粉萌发具有重要作用。添加适量的蔗糖对花粉的萌发有促进作用,但蔗糖浓度过高对花粉的萌发有抑制作用[33]。本实验研究得出N.alata、N.tabacumL.cv.K326、N.repanda和N.stocktonii花粉离体萌发时,随着蔗糖浓度的升高,花粉萌发率逐渐升高,但超过一定的量时花粉萌发受到抑制,而且N.alata和N.tabacumL.cv.K326花粉离体萌发比N.repanda和N.stocktonii需要的蔗糖浓度低,这可能是由不同烟草种之间内源糖含量差异引起,也可能是因为不同烟草种间花粉萌发的最适渗透压不同。硼酸能够增加花粉对糖的吸收、运转和代谢,促进花粉萌发[34]。硼对花药的花粉产生能力以及花粉粒生活力有重要的作用。硼能刺激花粉萌发和花粉管伸长[26]。4个烟草种离体萌发最适硼酸浓度都为0.05 g/L,超过该浓度则抑制花粉。最适蔗糖培养基中加入硼酸后,烟草花粉的萌发率有较大提高,这可能是由于硼离子与蔗糖容易形成络合物,更有助于糖在植物组织中运输,因此增加了蔗糖的吸收和代谢[35]。除此之外,硼还能参与果胶物质的合成,从而有利于花粉管壁的构造[13],促进花粉萌发。
钙是花粉萌发中非常重要的一个影响因子,Ca2+浓度过高将影响花粉管内细胞骨架的生理活动,会使花粉管顶端形成较厚的管壁,导致花粉管的生长受到抑制,适宜的Ca2+浓度能提高花粉的萌发率和促进花粉管的生长[26]。外源钙能促进花粉的萌发,一定浓度的钙还能使花粉管生长笔直,源钙浓度过高会使花粉管生长出现细弱、畸形,而且容易断落[36]。4个烟草种最适Ca2+浓度都为0.05 g/L,蔗糖+硼酸培养基中加入Ca2+极大促进了供试烟草种花粉离体萌发,表明烟草花粉离体萌发生长需要提供外源的Ca2+,这与花粉在柱头上萌发生长需要雌蕊提供Ca2+一致,可能原因是烟草花粉内源Ca2+不能满足花粉萌发生长所需。前人通过高效液相色谱(HPLC)测定GABA内源含量结果证明,烟草雌蕊组织中GABA从柱头到子房呈梯度分布,浓度介于0.75~4.2 mmol/L 之间[17],据此设计的外源GABA浓度为0.7~5 mmol/L。在最适的蔗糖+硼酸+Ca2+培养基中添加GABA表明,N.alata花粉萌发受抑制的GABA浓度较其他3个种低。由于前人报道指出,雌蕊来源的GABA可能通过促进花粉的钙内流而调控其萌发和生长[17-18],据此推测导致该结果的可能原因是,N.alata花粉内源钙离子可能较其他3个种高,导致其对GABA的耐受性较低。另外,在无Ca2+的培养基中添加GABA培养N.alata花粉,最适浓度的GABA促进N.alata花粉萌发的效果和Ca2+无显著差异,该结果表明,GABA也可能通过不依赖于促进外源钙离子内流的途径,在花粉萌发过程中发挥了作用。
4 小结
上述研究结果表明,外源提供的蔗糖、硼酸、Ca2+或GABA对于烟草花粉离体萌发具有重要作用,蔗糖、硼酸、GABA和Ca2+可能相互作用,共同促进花粉离体萌发。供试不同烟草种离体萌发所需最适外源物质浓度的差异,反映了烟草花粉的种间差异性,这种差异可能对烟草种间杂交亲和性的差异有影响。后续将进一步探究不同烟草种内源Ca2+或GABA等物质含量差异,及其对种间杂交亲和性差异的影响。
[1]陆焕海, 曹仲伟, 孙羿灵, 等.大气污染经雨水对植物花粉活性影响的初步分析[J].生物学通报, 2016, 51(7): 44-46.
[2]张 鹏, 张文超.花粉萌发与花粉管生长的信号调控机制[J].广东农业科学, 2013(5): 138-141.
[3]LORD E M, RUSSEELL S D.The mechanisms of pollination and fertilization in plants[J].Annu Rev Cell Dev Biol, 2002, 18(1): 81-105.
[4]YIN J L, ZHAO H E.Summary of influencial factors on pollen viability and its preservation methods[J].Chin Agri Sci Bull, 2005, 21(4): 110-113.
[5]许 珂, 古松枣, 江 莎.金银忍冬花粉离体萌发初探[J].热带亚热带植物学报, 2008, 16(2): 109-115.
[6]李桂云, 顾景梅, 王 峰.不同培养基对果树花粉发芽率影响的试验[J].山西果树, 2001(1): 4-5.
[7]张春玲, 张子平.蔗糖不同质量分数对杉松新鲜花粉萌发的影响[J].吉林林业科技, 2016, 45(2): 7-9.
[8]BLEVIN D G, LUKASZEWSKI K M.Boron in plant structure and function[J].Annual Review Plant Physiology and Plant Molecular Biology, 1998, 49: 481-500.
[9]LI Y Q, CHEN F, LINSKENS H F, et al.Distribution of unesterified and esterified pectins in cell walls of pollen tube of flowering plants[J].Sexual Plant Reproduction, 1994, 7(3): 145-152.
[10]金昌海, 水野雅史, 阚 娟, 等.不同品种苹果采后后熟软化过程中细胞壁多糖的降解[J].植物生理与分子生物学学报, 2006, 32(6): 617-626.
[11]董晓芬, 郑剑宇.硼酸和钙离子对红叶李花粉萌发的影响[J].大众科技, 2007(99): 134-135.
[12]GRAAF B, DERKSEN J W M, MARIANI C.Pollen and pistil in the progamic phase[J].Sex Plant Report, 2001, 14(1/2): 41-55.
[13]加藤幸雄, 志佐诚(译).植物生殖生理学[M].北京:科学出版社, 1987: 176-180.
[14]KONRAD K R, WUDICK M M, FEIJJ A.Calcium regulation of tip growth: new genes for old mechanisms[J].Curr Opin Plant Biol, 2011, 14(6): 721-730.
[15]胡适宜.被子植物胚胎学[M].北京:科学出版社, 2002: 29-58.
[16]MA H.Plant reproduction: GABA gradient guidance and growth[J].Current Biology, 2003, 13(21): R834-R836.
[17]赵 丽, 赵志龙, 龚汉雨, 等.GABA对烟草花粉管Ca2+和K+的动态调节表明离子通道参与了花粉管生长的调控[J].中国细胞生物学学报, 2013, 35(5): 668-675.
[18]YU G H, ZOU J, FENG J, et al.Exogenous γ-aminobutyric acid (GABA) affects pollen tube growth via modulating putative Ca2+-permeable membrane channels and is coupled to negative regulation on glutamate decarboxylase[J].Journal of Experimental Botany, 2014, 65(12): 3235-3248.
[19]李宗平, 张俊杰, 彭 灏, 等.烟草花粉活力鉴定方法筛选[J].中国烟草科学, 2013, 34(4): 80-82.
[20]李树美.几种烟草品种花粉形态的研究[J].安徽师范大学学报, 1998, 21(2): 128-131.
[21]黄腾波, 尤瑞麟.烟草花粉低温处理过程中超微结构变化的研究[J].北京大学学报(自然科学版), 2003, 39(6): 780-791.
[22]许美玲, 李永平.烟草种植资源图鉴[M].北京:科学出版社, 2009: 780-781.
[23]廖菊够, 康洪梅, 代瑾然, 等.温度对斯托克通氏烟草雄配子体形成和发育的影响[J].广西植物, 2013, 33(5): 614-619.
[24]廖菊够, 康洪梅, 朱 楠, 等.黄花烟草(Nicotianarustica)、迪勃纳氏烟草(Nicotianadebneyi)与NicotianatabacumK326互交亲和性的荧光鉴定[J].西北农业学报, 2014, 23(1): 53-59.
[25]廖菊够, 康洪梅, 代瑾然, 等.烟草大小孢子发生和雌雄配子体发育研究[J].西北植物学报, 2013, 33(1): 11-16.
[26]杨 瑞, 高 赛, 王金金, 等.外源硼和钙对“索邦”百合花粉萌发和花粉管生长的影响[J].电子显微学报, 2014, 33(4): 368-372.
[27]DEARNALEY J D, LEVINA N N, LEW R R,et al.Interrelationships between cytoplasmic Ca2+peaks pollen hydration and plasmam embrane conductance during compatible and incompatible pollinations ofBrasssiannapuspapilae[J].Plant Cell Physiol, 1997, 38(9): 985-999.
[28]FRANKLINR-TONG V E, HACKETT G, HEPLER P K.Ratio-imaging of Ca2+in the self-incompatibility response in pollen tubes ofPapaverrhoeas[J].Plant Joumal, 1997, 12(6): 1375-1386.
[29]FRANKLINR-TONG V E, RIDE J P, FRANKLIN F C H.Recombinant stigmatic self-incompatibility (S-)protein elicits a Ca2+transient in pollen ofPapaverrhoeas[J].P lant J, 1995, 8(2):299-307.
[30]SNOWMANA B N, KOVARBO D R, SHEVCHENKOA G, et al.Signal-mediated depolymerization of actin in pollen during the self-incompatibility response [J].Plant Cell, 2002, 14(10): 2613-2626.
[31]尤 扬, 贾文庆, 王 慧, 等.Ca2+对韭兰花粉萌发和花粉管生长的影响[J].亚热带植物科学, 2013, 39(2): 35-37.
[32]王晓华, 郝怀庆, 王钦丽, 等.花粉管细胞结构与生长机制研究进展[J].植物学通报, 2007, 24(3): 340-354.
[33]冯建文, 邹养军, 董 军, 等.‘嘎拉’苹果花粉萌发中培养基组分和培养条件优化[J].西北农业学报, 2016, 25(11):1643-1649.
[34]赖俊声, 龚榜初, 江锡兵, 等.锥栗花粉萌发率及生活力测定研究[J].西南林业大学学报, 2017, 37(1): 42-47.
[35]李佳荣, 杨鹏鸣.硼酸与蔗糖对百合花粉活力的影响[J].安徽农业科学, 2010, 38(18): 9494-9495.
[36]韩 霜.钙和硼对菊花品种金陵丰收花粉萌发及花粉管生长的影响[J].江苏农业科学, 2016, 44(5): 258-261.
猜你喜欢
中国果业信息(2022年1期)2022-11-28
三农资讯半月报(2020年10期)2020-06-08
设备管理与维修(2019年9期)2019-09-12
山东林业科技(2019年2期)2019-06-03
生物学杂志(2018年4期)2018-08-15
现代园艺(2017年11期)2017-06-28
中国果业信息(2017年8期)2017-01-13
奥秘(2016年10期)2016-12-17
大连工业大学学报(2015年4期)2015-12-11
中国塑料(2015年7期)2015-10-14
