
毛乌素沙地天然植物多样性组成及区系特征分析
2018-06-12 06:55:46段义忠杜忠毓亢福仁
西北植物学报 2018年4期
段义忠,李 娟,杜忠毓,亢福仁*
(1 榆林学院 生命科学学院,陕西榆林719000; 2 陕西省陕北生态修复重点实验室,陕西榆林 719000)
中国西北干旱地区分布着大量的沙漠、沙地,毛乌素沙地正是位于其中的中国四大沙地之一。该沙地为中国典型的农牧交错区,长期的放牧及农业生产对该区的生物多样性造成了巨大的威胁[1-2]。毛乌素沙地东南接壤黄土高原,西北紧靠库不齐沙漠,西南临近乌兰布和沙漠,因此该区正是湿润森林到干旱草原、荒漠的过渡区域。该区流动沙丘较少,植被覆盖率高,植物多样性较临近沙漠高[3-4]。植物区系研究是关于某一地区所有植物种类的组成、分布规律以及起源进化和演变历史的科学,开展植物区系研究是植物恢复的前提,也是植物资源开发利用的基础[5-7]。植物区系地理研究已经进入古植物、古地质及系统发育多学科融合交叉、精细尺度的起源与进化及空间格局形成机制阶段。植物区系分析方法从数据挖掘到大数据分析再到多方法的运用的全数据库构建大数据分析阶段,大数据使得植物区系地理研究由定性的现象描述进入到区系区划的定量研究及区系格局形成机制研究[8-10]。
目前关于毛乌素沙地植物研究多集中在植物资源、植物生理生态及植物群落特征方面[11-13],对该地区植物区系特征研究只限于低湿地维管植物等具体类群的植物区系研究[14],但对毛乌素沙地天然植物多样性组成及区系特征分析尚未见相关研究报道。从2014年到2016年,陕西省治沙研究所与榆林学院多名学者对毛乌素沙地及其边缘地区的天然植物进行了连续调查。本研究基于野外调查的基础上,运用植物区系地理学基本方法及原理,对毛乌素沙地天然植物多样性组成及区系特征进行研究,并与周围的沙地(沙漠)植物区系进行比较分析,本研究将为毛乌素沙地珍稀植物保护,沙生植物资源开发利用及生物多样性评价提供理论依据及实践指导。
1 研究区概况
毛乌素沙地位于鄂尔多斯高原与黄土高原之间的冲积平原凹地上,是一个多层次的生态过渡带,气候上处于干旱、半干旱区向半湿润区的过渡带,地理位置介于N37°30′~39°20′,E107°20′~111°30′,行政区划属于内蒙古自治区鄂尔多斯市、包头市南部、宁夏东北部和陕西榆林市北部,总面积4.22万km2,海拔大多在1 000~1 600 m(图1)[15]。毛乌素沙地气候属于中温带气候,年平均温度为6.0~8.5 ℃,年降水量为250~440 mm,集中在7~9月,占全年降水60%~75%,尤以8月为多;年平均蒸发量2 041~2 424 mm,气候干燥度在1.5~2.0之间,年平均风速为2.9 m/s,其中3~6月份很强,4~5月份最大风速超过17 m/s的日数很多。地表、5 cm处和10 cm处年平均地温分别是6.4 ℃、4.1 ℃和6.3 ℃,地表最高、最低地温的极值出现在每年7月(44.8 ℃)和1月(-20.5 ℃)。地带性土壤从东北向西南,土壤由淡栗钙土变成棕钙土,南部受局部地形与母质影响为沙黄土[16]。毛乌素沙地植被地带主要由西部边缘的荒漠草原亚地带、中东部的干草原地带以及东南边缘的干草原地带-森林草原过渡地带组成,植被类型主要有梁地上的草原与灌木植被,半固定、固定以及沙地上的沙生灌丛植被,滩地上的草甸、盐土与沼泽植被[17]。
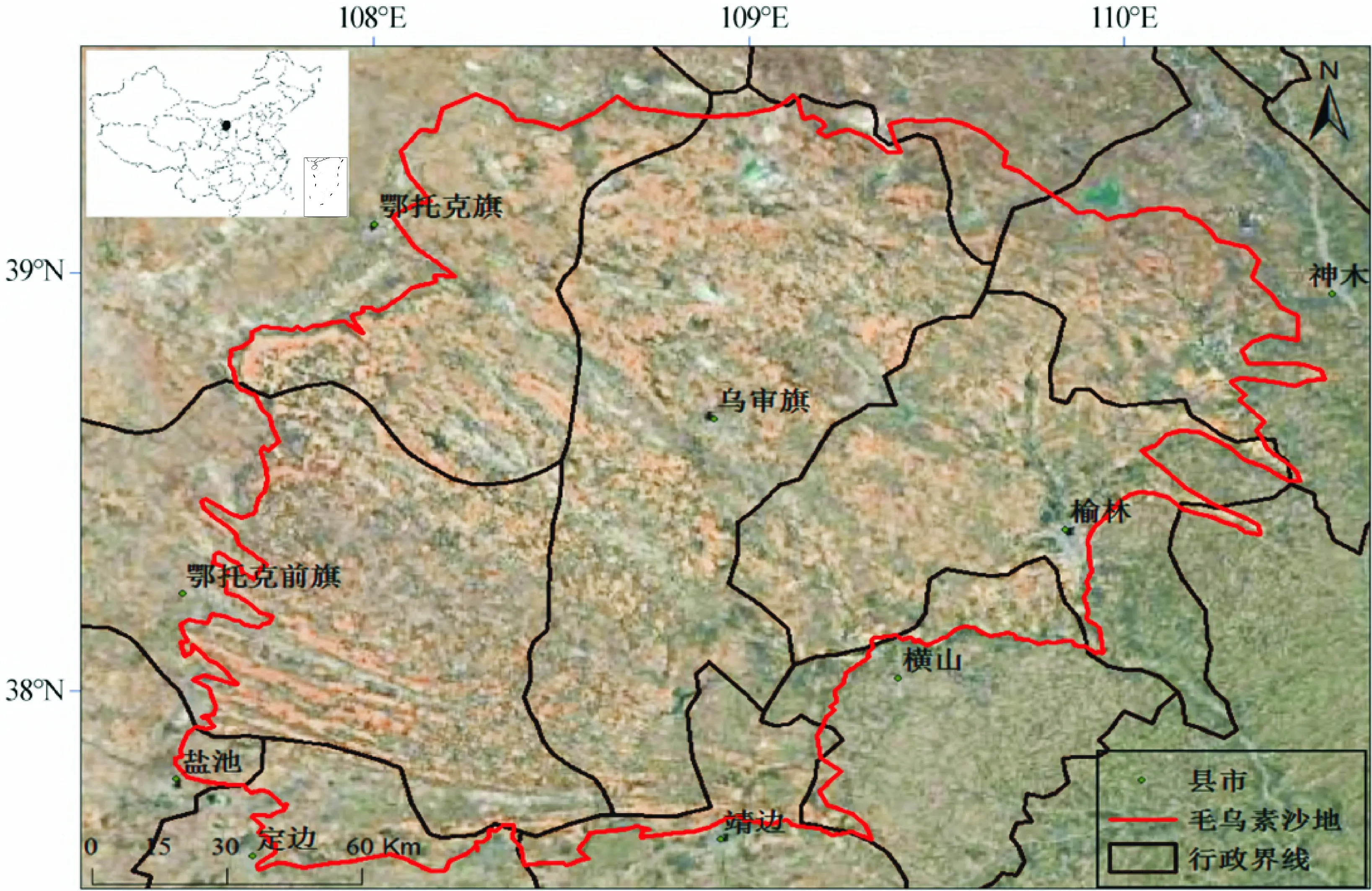
图1 毛乌素沙地地理位置Fig.1 Geographic location of Mu Us sandy land
2 研究方法
从2014年起,历经前后3年时间,联合考察组多次进入毛乌素沙地,对该区域的植物进行网格式标本采集工作(标本存放在榆林学院标本馆),共采集该区域植物标本3 142份。参照《中国植物志》、《内蒙古植物志》以及中国植物图像库等文献及图库对标本进行鉴定[18-19]。毛乌素沙地植物名录编制参照《FloraofChina》进行,植物区系分类类型参照吴征镒及李锡文等关于中国植物区系与植被地理划分方法进行归类[20-22]。
3 结果与分析
3.1 植物种类组成
通过野外调查统计并建立植物区系统计库,调查毛乌素沙地的主要植被类型,对植物资源进行详尽的分类(科、属、种)。根据初步调查和资料记载,共统计毛乌素沙地天然植物共有772种,分属于90科360属(表1)。其中,苔藓植物有1科1属1种,占毛乌素沙地植物总科数的1.11%,总属数的0.28%,总种数的0.13%;蕨类植物有7科7属12种,占毛乌素沙地植物总科数的7.78%,总属数的1.94%,总种数的1.55%;裸子植物有2科4属7种,占毛乌素沙地植物总科数的2.22%,总属数的1.11%,总种数的0.91%;单子叶植物有15科71属178种,占毛乌素沙地植物总科数的16.67%,总属数的19.72%,总种数的23.06%;双子叶植物有65科277属574种,占毛乌素沙地植物总科数的72.22%,总属数的76.94%,总种数的74.35%(表1)。
毛乌素沙地天然植物组成与内蒙古及全国的科、属、种的比较结果见表2,毛乌素沙地天然植物科、属、种组成占内蒙古自治区植物科、属、种的比例分别是47.12%、41.14%及27.75%,毛乌素沙地4.22万km2,约占内蒙古自治区面积的3.58%,上述比例说明毛乌素沙地科、属、种组成比较丰富;毛乌素沙地植物科、属、种组成占中国全境植物科、属、种比例分别是17.44%、9.45%及2.56%,说明毛乌素沙地植物是中国植物区系的重要组成部分,具有很大的科研与开发价值。
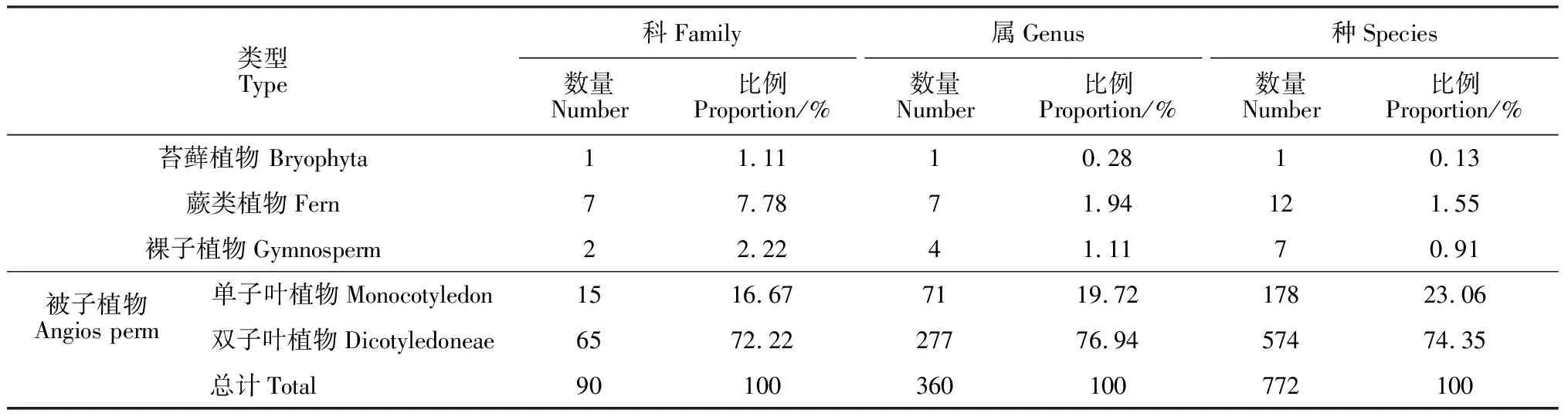
表1 毛乌素沙地天然植物科、属、种组成
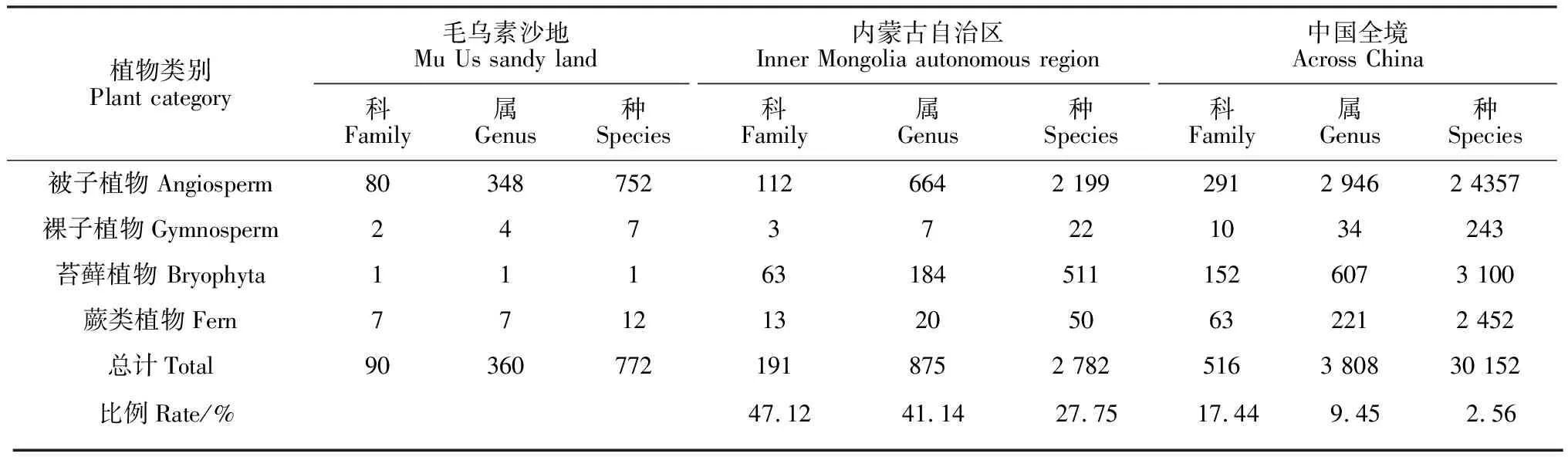
表2 毛乌素沙地天然植物科、属、种与其他区域的比较
3.2 科的多样性分析
根据初步调查结果(表3),毛乌素沙地植物科的物种数大于20种有7科,占比为7.78%,包括豆科(Leguminosae,113种)、菊科(Compositae,95种)、禾本科(Gramineae,90种)、藜科(Chenopodiaceae,35种)、莎草科(Cyperaceae,30种)、蔷薇科(Rosaceae,30种)及唇形科(Labiatae,24种);含种数10~19种的科有13科,占比为14.44%,包括毛茛科(Ranunculaceae,19种)、百合科(Liliaceae,17种)、杨柳科(Salicaceae,14种)、伞形科(Apiaceae,14种)、紫草科(Boraginaceae,13种)、眼子菜科(Potamogetonaceae,11种)、十字花科(Brassicaceae, 11种)、蓼科(Polygonaceae,11种)、萝藦科(Asclepiadaceae,11种)、玄参科(Scrophulariaceae,11种)、茄科(Solanaceae,10种)、大戟科(Euphorbiaceae,10种)及石竹科(Caryophyllaceae,10种);含种数5~9种的科有8科,占比为8.89%,包括旋花科(Convolvulaceae,9种)、龙胆科(Gentianaceae,7种)、香蒲科(Typhaceae,6种)、榆科(Ulmaceae,5种)、罂粟科(Papaveraceae、5种)、蒺藜科(Zygophyllaceae、5种)及葡萄科(Vitaceae,5种);含2~4种的有36科,占比为40.00%,代表性科包括桑科(Moraceae,4种)、柏科(Cupressaceae,4种)、苋科(Amaranthaceae,4种)、景天科(Crassulaceae,4种)、麻黄科(Ephedraceae,3种)、水龙骨科(Polypodiaceae,3种)、木贼科(Equisetaceae,3种)、马齿苋科(Portulacaceae,3种)、小檗科(Berberidaceae,3种)及牻牛儿苗科(Geraniaceae,3种);有26科仅含1个物种,具体科名见表4。毛乌素沙地≥20种的科含种数占总种数的比例是54.02%,10~19种的科含种数占总种数的比例是23.06%,5~9种的科含种数占总种数的比例是6.09%,2~4种的科含种数占总种数的比例是13.47%,单种科的含种数占总种数的比例是3.37%。由此可以看出,该区域大科占总科比例虽然很少,但却包含了一半以上的物种数,大科在该区域的植物区系组成和维持物种多样性起着重要作用。
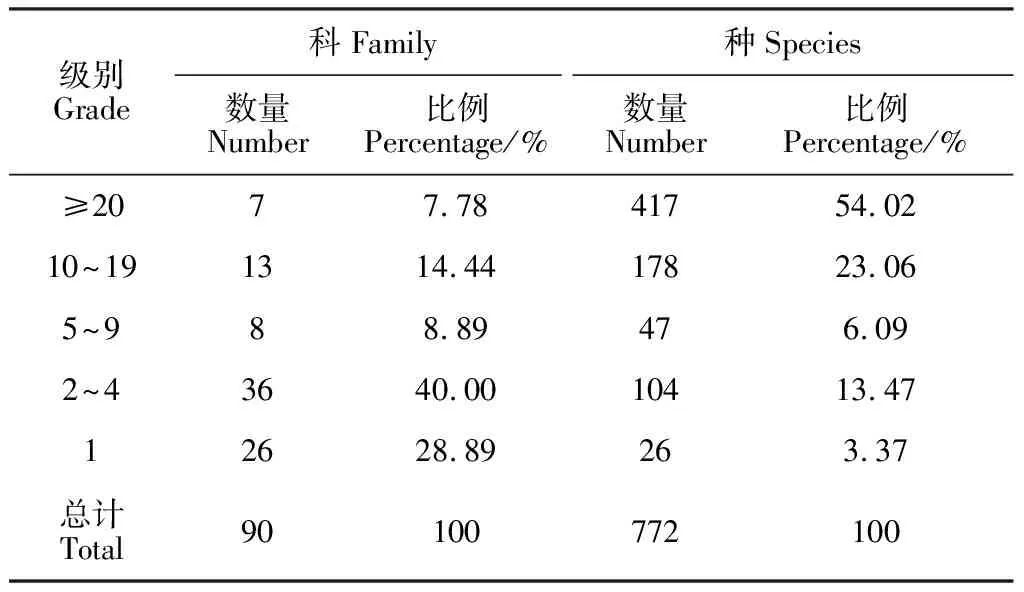
表3 毛乌素沙地天然植物科内种的组成数
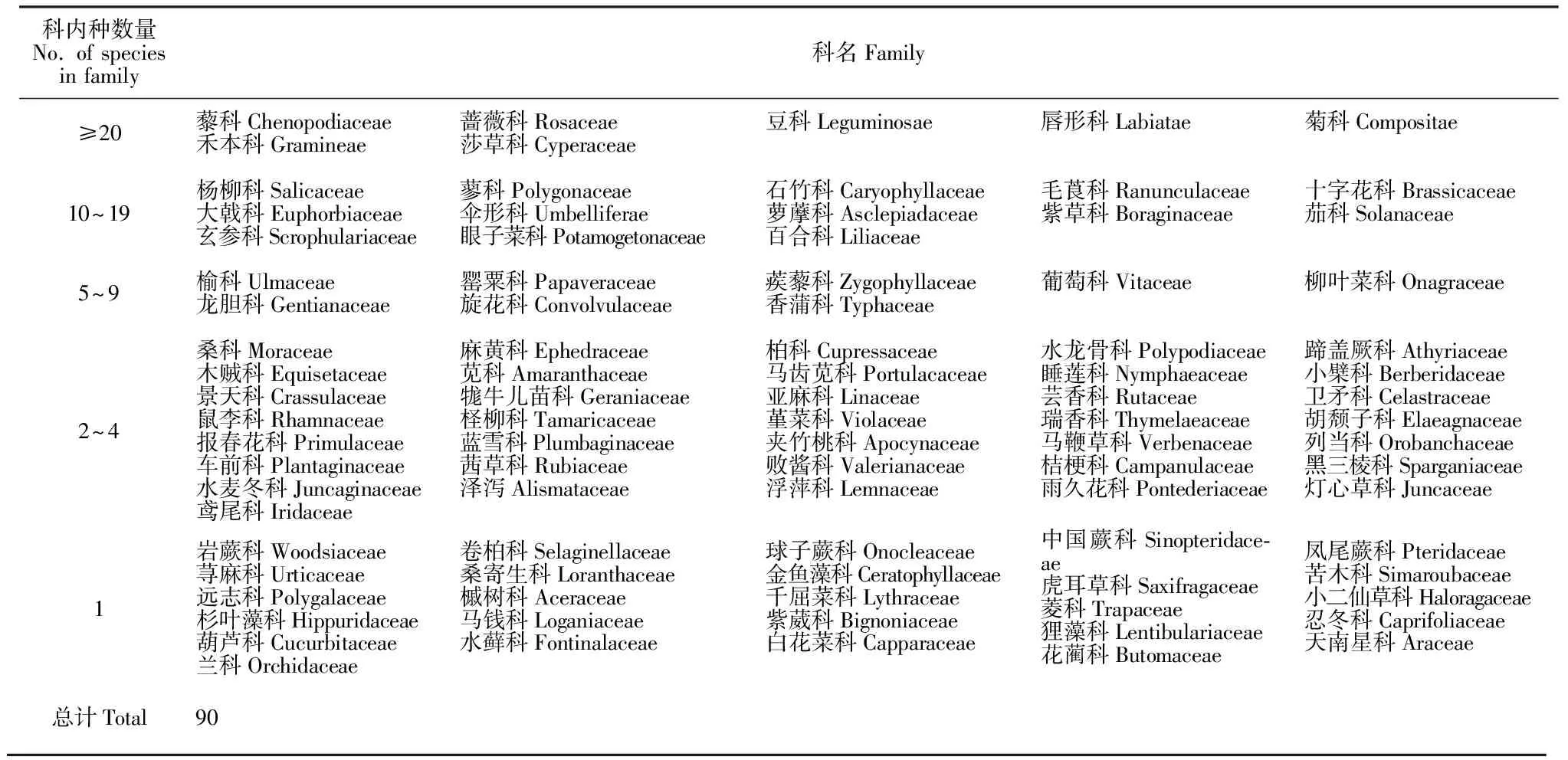
表4 毛乌素沙地天然植物科内物种数量大小排序
3.3 属的多样性分析
毛乌素沙地植物由360个属组成。统计该区域不同物种数等级的属数结果见表5,结果表明本区含11种以上的有7个属共106种,包括蒿属(Artemisia,30种)、黄耆属(Astragalus,15种)、柳属(Salix,13种)、野豌豆属(Vicia,13种)、棘豆属(Oxytropis,12种)、锦鸡儿属(Caragana,12种)、眼子菜属(Potamogeton,11种);含6~10种13属共99种,包括葱属(Allium,10种)、鹅绒藤属(Cynanchum,10种)、委陵菜属(Potentilla,9)、藜属(Chenopodium,8)、藨草属(Scirpus,7种)、大戟属(Euphorbia,7种)、胡枝子属(Lespedeza,7种)、莎草属(Cyperus,7种)、风毛菊属(Saussurea,6种)、香蒲属(Typha,6种)、岩黄耆属(Hedysarum,6种)、针茅属(Stipa,7种);含2~5种有104属331种,代表性属有铁线莲属(Clematis,5种)、柳叶菜属(Epilobium,5种)、天门冬属(Asparagus,5种)、独行菜属(Lepidium,5种)、甘草属(Glycyrrhiza,4种)、山黧豆属(Lathyrus,4种)、稗属(Echinochloa,4种)、车前属(Plantago,4种)、榆属(Ulmus,4种)、灯心草属(Juncus,4种)、芨芨草属(Achnatherum,3种)、地肤属(Kochia,3种)、苜蓿属(Medicago,3种)、车轴草属(Trifolium,2种)、扁穗草属(Blysmus,2种)等;含1种的单种属有236属236种,代表性属包括阿魏属(Ferula)、翠雀属(Delphinium)、短星菊属(Brachyactis)、白刺属(Nitraria)、荨麻属(Urtica)、鸡眼草属(Kummerowia)、盐生草属(Halogeton)、白头翁属(Pulsatilla)、针枝蓼属(Atraphaxia),其中鹅肠菜属(Myosoton)、桔梗属(Platycodon)、防风属(Saposhnikoia)等为世界单型属。含11种以上的属占毛乌素沙地天然植物总种数的13.73%,含6~10种的属占比为12.82%,含2~5种的属占比为42.88%,单属种占比为30.57%,单种属及含2~5种的属占到的比例73.45%,占该区域植物物种数的绝大部分,而大属包含的种数相对较少,而单种属数量较高,由此可以看出毛乌素沙地植物物种多样性相对较高。
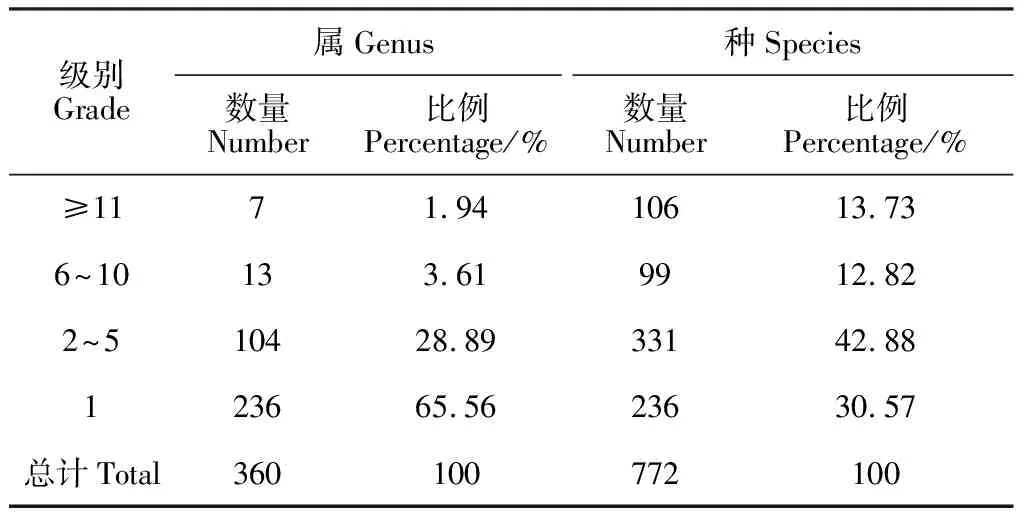
表5 毛乌素沙地天然植物属内种的组成
3.4 科的区系成分分析
植物科的分布反映了一个地区植物区系的组成及其性质,科作为一个自然的分类群,由不同发源地和分布格局的属组成,因此比属具有更为漫长的历史,科主要强调类群之间演化的共有属性和阶段性。毛乌素沙地天然植物90科划分为8个分布类型及4个变型(表6)。其中,世界分布38科,占毛乌素沙地天然植被总科数的42.22%,包括藜科、蔷薇科、唇形科、菊科、禾本科、莎草科、石竹科、玄参科、眼子菜科、百合科、旋花科、香蒲科(Typhaceae)、金鱼藻科(Ceratophyllaceae)、远志科(Polygalaceae)、千屈菜科(Lythraceae)、小二仙草科(Haloragaceae)、杉叶藻科(Hippuridaceae)等;泛热带分布20科,占比为22.22%,包括大戟科(Euphorbiaceae)、萝藦科(Asclepiadaceae)、茄科(Solanaceae)、榆科(Ulmaceae)、蒺藜科(Zygophyllaceae)、荨麻科(Urticaceae)、桑寄生科(Loranthaceae)、马钱科(Loganiaceae)、葫芦科(Cucurbitaceae)、天南星科(Araceae)、兰科(Orchidaceae)等;热带亚洲、大洋洲(至新西兰) 和中、南美(或墨西哥) 间断分布1科,占比为1.11%,即鸢尾科;旧世界热带分布1科,占比为1.11%,即紫葳科(Bignoniaceae);温带分布18科,占比为17.78%,包括蓼科、毛茛科、十字花科、紫草科(Boraginaceae)、罂粟科、龙胆科、虎耳草科(Saxifragaceae)、忍冬科(Caprifoliaceae)、岩蕨科(Woodsiaceae)、球子蕨科(Onocleaceae)、胡颓子科(Elaeagnaceae)等;环北极分布1科,占比为1.11%,即槭树科(Aceraceae);北温带和南温带(全温带)间断分布2科,占比为2.22%,包括柏科(Cupressaceae)及杨柳科(Salicaceae);东亚和北美洲间断分布1科,即小檗科(Berberidaceae);旧世界温带分布2科,占比为2.22%,包括柽柳科(Tamaricaceae)及花蔺科(Butomaceae);欧亚和南部非洲(有时也在大洋洲)间断分布1科,占比为1.11%,即菱科;温带亚洲分布1科,占比为1.11%,即卷柏科(Selaginellaceae);地中海区、西亚至中亚分布6科,占比为6.67%,包括蹄盖蕨科(Athyriaceaea)、水龙骨科(Polypodiaceae)、中国蕨科(Sinopteridaceae)、凤尾蕨科(Pteridaceae)、水藓科(Fontinalaceae)、蓝雪科(Plumbaginaceae)。上述结果表明毛乌素沙地植物区系科的类型组成中以世界分布、泛热带分布及温带分布为主,带有温带性质。

表6 毛乌素沙地天然植物科的分布区类型统计
3.5 属的区系成分分析
吴征镒认为地理成分是按各分类单位(特别是种)的现代分布区来分析的,可以归为若干分布区类型,然而它并不反映植物区系的发生。他在对中国种子植物的3 116个属进行了详细的研究后,把中国种子植物属的分布区归纳为15个类型和31个变型(表7)。现在对属的分布区划分多采用这一方案。根据吴征镒先生对中国种子植物属的分布区类型的划分,现将毛乌素沙地天然植物区系的360属划分为15个分布区类型及10个变型,结果见表7。可以看出毛乌素沙地植物区系属的地理成分中北温带分布类型及其亚类型最多,共101属,占比达到了28.06%,代表性属包括刺柏属(Juniperus)、冰草属(Agropyron)、播娘蒿属(Descurainia)、发草属(Deschampsia)、紫草属(Lithospermum)、紫荆属(Cercis);世界分布类型共48属,占比达到了13.33%,代表性属包括苍耳属(Xanthium)、车前属(Plantago)、扁豆属(Lablab)、补血草属(Limonium)、百金花属(Centaurium)、灯心草属(Juncus);泛热带分布类型29属,占比达到8.05%,代表性属包括刀豆属(Canavalia)、鹅绒藤属(Cynanchum)、锋芒草属(Tragus)、扁莎属(Pycreus)、打碗花属(Calystegia)、醉鱼草属(Buddleja)、泽兰属(Eupatorium);旧世界温带分布类型45属,占比达到12.50%,代表性属包括益母草属(Leonurus)、隐花草属(Crypsis)、隐子草属(Cleistogenes)、沙棘属(Hippophae)、青兰属(Dracocephalum)、荆芥属(Nepeta)等;地中海区、西亚至中亚分布类型28属,占比达到7.77%,代表性属包括燥原荠属(Ptilotricum)、盐爪爪属(Kalidium)、梭梭属(Haloxylon)、亚麻荠属(Camelina)、木贼属(Equisetum)、念珠芥属(Torularia);东亚(东喜马拉雅-日本)分布类型27属,占比达到7.50%,代表属包括马鞍树属(Maackia)、党参属(Codonopsis)、地黄属(Rehmannia)、败酱属(Patrinia)、斑种草属(Bothriospermum);中国特有分布类型10属,占比达到2.78%,代表性属包括知母属(Anemarrhena)、菊三七属(Gynura)、文冠果属(Xanthoceras)、百花蒿属(Stilpnolepis)等,此外该区域还存在一些热带分布类型,代表性属包括栝楼属(Trichosanthes)、大戟属(Euphorbia)、粉背蕨属(Aleuritopteris)、杠柳属(Periploca)、荩草属(Arthraxon)、木兰属(Magnolia)、桃属(Amygdalus)、葛属(Pueraria)、漏芦属(Stemmacantha)、蓼属(Polygonum)。上述结果说明毛乌素沙地植物区系属的类型组成具有明显的温带性质, 且向地中海区、西亚至中亚分布类型过渡性质,但与热带植物区系也存在一定的亲缘关系。
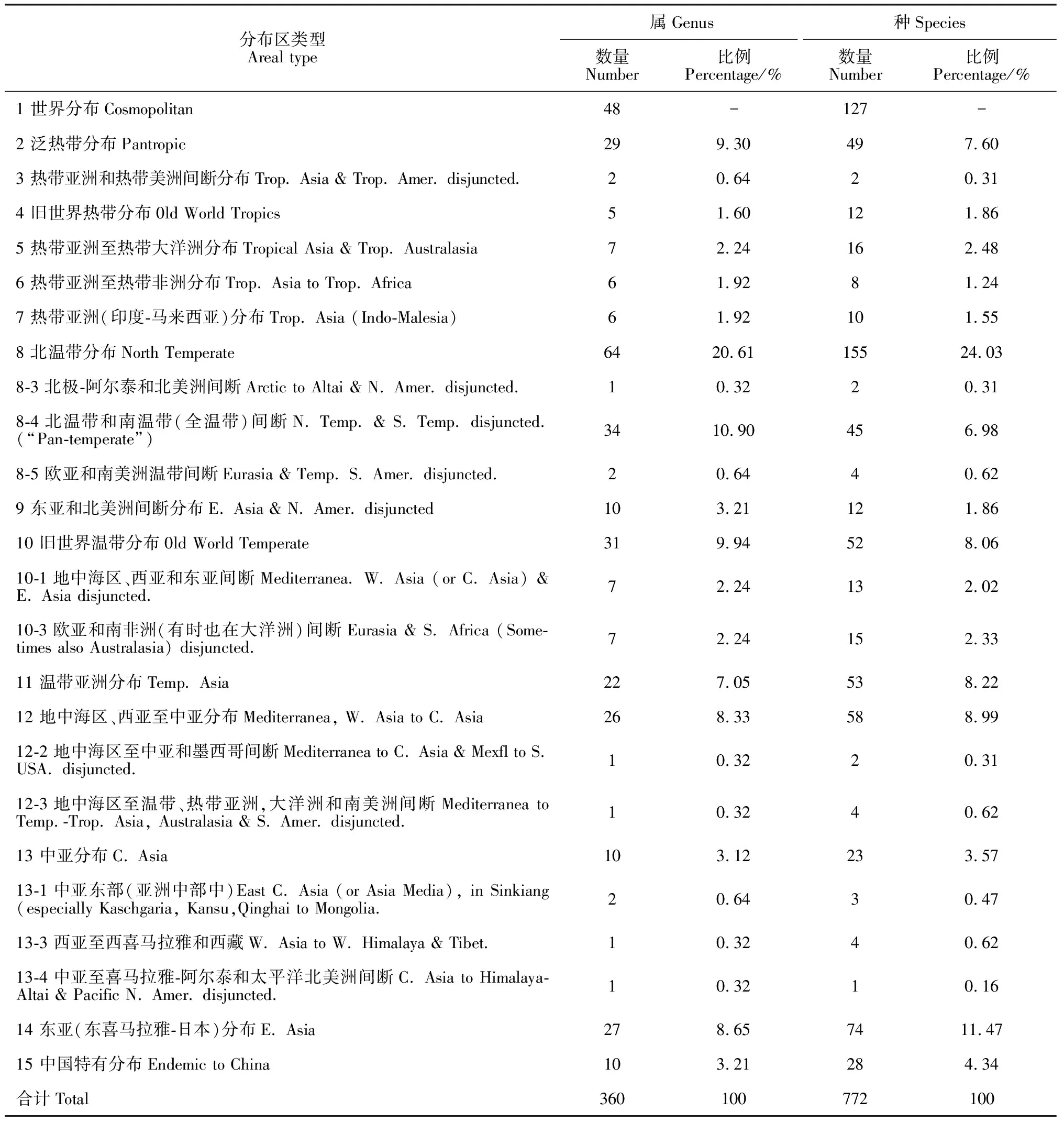
表7 毛乌素沙地天然植物属和种的分布区类型统计
3.6 毛乌素沙地及周围沙地(沙漠)植物属的分布区类型比较
属是由种所组成的,它在分类学上所包含的种通常具有同一起源和相似的进化趋势,属的分类特征也相对稳定,占有比较稳定的分布区,随着地理环境的变化发生分异,因而具有比较明显的自然保护区性差异,因此植物属的分布型比科更能反映植物的演化扩展过程、区域分异及地理特征[24-25]。为对比分析毛乌素沙地植物区系分布类型与周围沙地(沙漠)的关系,在已有文献数据的基础上[26-30],选取浑善达克沙地、乌兰布和沙漠、库布齐沙漠及库姆塔格沙漠属的植物区系做对比分析,结果见表8。从表8各区域属的总数来看,从多到少顺序依次是毛乌素沙地、浑善达克沙地、库布齐沙漠、乌兰布和沙漠及库姆塔格沙漠,表明随着5个区域由东向西变化,生物多样性从高到底。其中,浑善达克沙地属的分布区类型以温带成分占绝对优势,表明该植物区系具有明显的温带性质;库布齐沙漠属的植物区系分布类型虽仍以温带成分为主,但不占绝对优势,地中海区、西亚至中亚分布成分的比例逐渐增大;乌兰布和沙漠温带成分比例继续降低,地中海区、西亚至中亚分布的比例继续增大,单种属比例较大,说明该区域植物区系整体多样性较高;库姆塔格沙漠植物区系属于典型的荒漠植物区系,该区域植物区系温带成分,地中海区、西亚至中亚分布的比例几乎持平,较大科属已不占优势,单种属、寡种属构成库姆塔格沙漠植物区系的主体。
应用软件IBM SPSS.20,利用植物区系属的分布类型数据[31],对毛乌素沙地及相邻的沙地(沙漠)植物区系进行聚类分析,结果见图2。其中,5个沙地(沙漠)植物区系被分为2大类型,第一大类将毛乌素沙地及浑善达克沙地聚类在一起,表明这两个区域都处于湿润森林向干旱草原、荒漠的过渡地带,地形地貌及气候环境特征相似,因此两个区域的植物区系相似性较高;第二大类将库布齐沙漠及乌兰布和沙漠聚类,再和库姆塔格沙漠聚类在一起组成第二大类,主要原因在于库布齐沙漠及乌兰布和沙漠由于地理位置更为接近,两个区域共同属的比例很高,因此植物区系类型相似度很高,库姆塔格沙漠位于5个沙地(沙漠)的最西端,物种数最少,温带分布及地中海分布是该区植物区系的主要类型,是典型的荒漠植物区系,与库布齐沙漠及乌兰布和沙漠有很多相同的属,因此与两个区域聚集在一类。
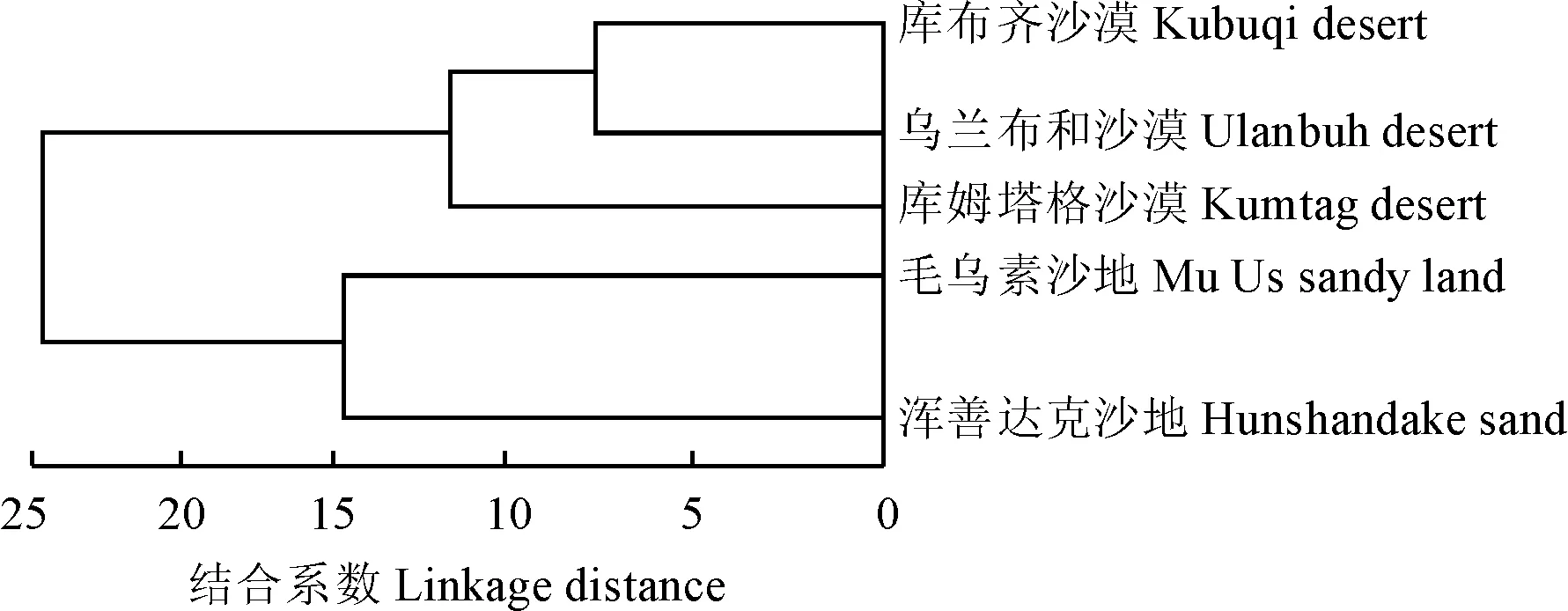
图2 毛乌素沙地与周边4个沙地(沙漠)植物区系聚类分析Fig.2 Cluster analysis of the plant flora relationship between Mu Us sandy land and other 4 sandy lands (desert)
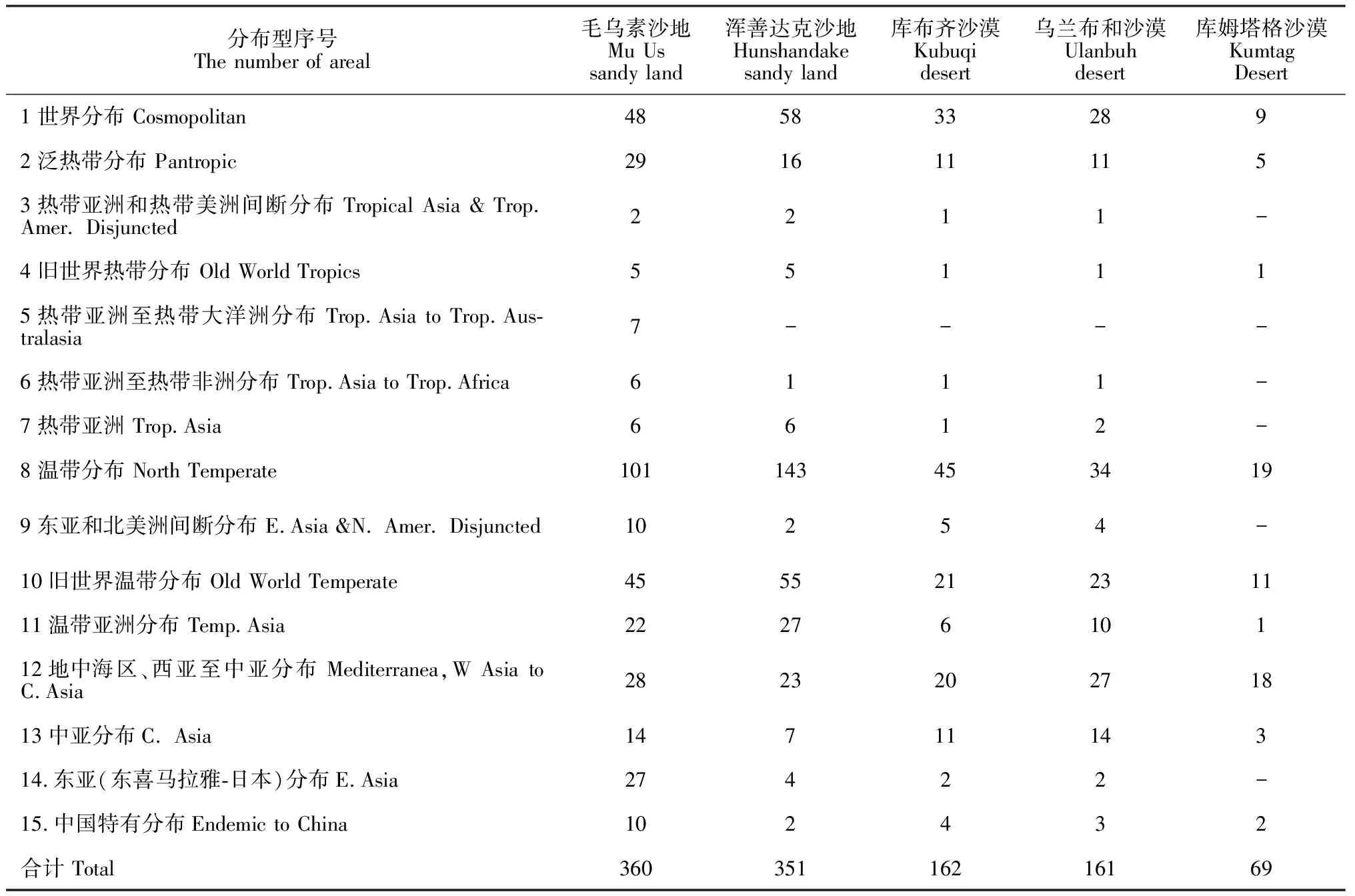
分布型序号The number of areal毛乌素沙地Mu Us sandy land浑善达克沙地Hunshandake sandy land库布齐沙漠Kubuqidesert乌兰布和沙漠Ulanbuh desert库姆塔格沙漠Kumtag Desert1世界分布 Cosmopolitan4858332892泛热带分布 Pantropic2916111153热带亚洲和热带美洲间断分布 Tropical Asia & Trop. Amer. Disjuncted2211-4旧世界热带分布 Old World Tropics551115热带亚洲至热带大洋洲分布 Trop.Asia to Trop.Aus-tralasia7----6热带亚洲至热带非洲分布 Trop.Asia to Trop.Africa6111-7热带亚洲 Trop.Asia6612-8温带分布 North Temperate1011434534199东亚和北美洲间断分布 E.Asia &N. Amer. Disjuncted10254-10旧世界温带分布 Old World Temperate455521231111温带亚洲分布 Temp.Asia2227610112地中海区、西亚至中亚分布 Mediterranea,W Asia to C.Asia282320271813中亚分布C. Asia1471114314.东亚(东喜马拉雅-日本)分布E.Asia27422-15.中国特有分布Endemic to China102432合计 Total36035116216169
4 讨 论
毛乌素沙地地处半湿润区向干旱、半干旱区的过渡地带,毛乌素沙地天然植物共有772种,分属于90科360属,占内蒙古自治区植物科、属、种的比例分别是47.12%、41.14%及27.75%。单种属、寡种属占到的比例达到73.45%,而大属包含的种数相对较少,因此毛乌素沙地植物物种多样性相对丰富,是中国植物区系的重要组成部分,该区域的优势科为豆科、菊科及禾本科,优势属为蒿属。
毛乌素沙地天然植物90科划分为10个分布区类型及4个变型,360属划分为15个分布区类型及10个变型。科的植物区系类型以世界分布、泛热带分布及温带分布为主,还包含了一定比例的地中海区、西亚至中亚分布类型,表明毛乌素沙地植物区系存在明显的过渡性,优势科几乎全部由世界分布科组成,反映了毛乌素沙地植物区系与所在气候带干旱、半干旱特征。从毛乌素沙地植物区系的组成来看,该区包含一些古老的科属,有些为孑遗物种,例如蒺藜科的霸王和白刺,麻黄属的草麻黄、中麻黄及木贼麻黄等,同时还包括了一些间断分布的科属,例如温带和南温带(全温带)间断分布的柏科及杨柳科;东亚和北美洲间断分布1科的小檗科;欧亚和南部非洲(有时也在大洋洲)间断分布1科的菱科,说明毛乌素沙地包含一些起源比较古老,带有孑遗成分的植物。属的植物区系类型以温带分布为主,具有明显的温带性质, 但与热带植物区系也存在一定的亲缘关系,具有过渡性。毛乌素沙地科的植物区系组成以世界分布为主,属的植物区系组成以温带分布为主,反映了该区的植物区系与所处的地理位置和气候具有相适应的特性[32-33]。
毛乌素沙地及相邻的沙地(沙漠)植物区系存在明显的过渡性质,毛乌素沙地与浑善达克沙地以温带分布为主,但也存在一定比例的热带分布成分及地中海区、西亚至中亚分布类型,但随着经度不断西移,热带分布类型比例逐渐降低,温带分布及地中海分布比例升高,到库姆塔格沙漠热带分布属的类型基本消失,而温带分布及地中海分布比例升至一半以上,且二者比例接近。在野外调查中发现,由于毛乌素沙地处于农牧交错带,过度放牧及人类大规模的推地开荒,致使该区植被遭到了严重破坏,而且由于该区处于干旱、半干旱地带,植被一旦遭到破坏,就会发生根本性的变化,自然修复能力很差,最后导致沙地裸露,沙地逐渐沙漠化,很多中生植物将无法存活,导致旱生、超旱生植物的大量出现,对该区植物多样性及植物区系多样性造成严重破坏。因此本研究结果将对该区的植物资源保护、合理开发利用提供重要的理论依据和现实意义。
参考文献:
[1] 赵 爽,高福元,贾 佳,等. 毛乌素沙地风沙沉积物磁学特征及其古环境意义[J]. 地球物理学报, 2015,58(10): 3 706-3 718.
ZHAO S, GAO F Y, JIA J,etal. Magnetic Properties of eolian sand sediments in the Mu Us Desert and their paleoen vironmental significance[J].ChineseJournalofGeophysics, 2015,58(10): 3 706-3 718.
[2] 刘媖心. 试论中国沙漠地区植物区系的发生与形成[J].植物分类学报,1995,(2):131-143.
LIU Y X. A study on origin and formation of the Chinese desert floras[J].ActaPhytotaxonomicaSinica, 1995,(2): 131-143.
[3] 张 楠,刘翠英. 毛乌素沙地豆科植物资源调查研究[J]. 中国林副特产, 2009,(4): 69-70.
ZHANG N, LIU C Y. Study on the Resources of Legumes in Mu Us sandy land[J].ForestBy-ProductandSpecialityinChina, 2009,(4): 69-70.
[4] 白岗栓,李志熙,张占山. 毛乌素沙地高等植物资源[J]. 草地学报, 2006,(2): 170-180.
BAI G S, LI Z X, ZHANG Z S. Investigation of the Resources of Higher Plants in the Mu Us Sandy Land[J].ActaAgresiteSinica, 2006,(2): 170-180.
[5] 王荷生. 植物区系地理[M]. 北京:科学出版社, 1992: 1-155.
[6] 王荷生. 华北植物区系地理[M]. 北京:科学出版社, 1997: 1-149.
[7] 中国科学院《中国自然地理》编辑委员会. 中国自然地理——植物地理(上册)[M]. 北京:科学出版社, 1995: 29-89.
[8] 唐 瑶,张 震,王 瑞,等. 安徽大别山区种子植物区系的初步研究[J]. 西北植物学报, 2017,37(7): 1 438-1 446.
TANG Y, ZHANG Z, WANG R,etal. Preliminary study on seed plants flora of Dabie Mountain in Anhui Province[J].ActaBot.Boreal.-Occident.Sin, 2017,37(7): 1 438-1 446.
[9] 邓元德,何宗明,庄崇洋. 福建梁野山国家级自然保护区种子植物区系研究[J]. 西北植物学报, 2015,35(4): 830-836.
DENG Y D, HE Z M, ZHUANG C Y. Seed plant flora of Liangyeshan National Nature Reserve in Fujian Province[J].ActaBot.Boreal.-Occident.Sin, 2015,35(4): 830-836.
[10] 黄 升,吕世安,洪建峰,等. 湖北星斗山国家级自然保护区种子植物区系研究[J]. 植物科学学报, 2016,34(5): 684-694.
HUANG S, LÜ S, HONG J F,etal. Study on the floristics of seed plants in Xingdoushan Nature Reserve, Hubei[J].PlantScienceJournal, 2016,34(5): 684-694.
[11] 李志熙. 毛乌素沙地高等植被调查与研究[D]. 陕西杨陵:西北农林科技大学, 2005.
[12] 刘翠英. 毛乌素沙地植物资源调查及开发利用研究[J]. 干旱地区农业研究, 2006,(2): 183-188.
LIU C Y. Investigation and exploitation of plant resources in Maowusu Desert[J].AgriculturalResearchintheAridAreas, 2006,(2): 183-188.
[13] 朱媛君,张璞进,牛明丽,等. 毛乌素沙地丘间低地主要植物群落土壤酶活性[J]. 生态学杂志, 2016,35(8): 2 014-2 021.
ZHU Y J, ZHANG P J, NIU M L,etal. Soil enzyme activities of the main plant communities in inter-dune lowland of Mu Us Sandy Land[J].ChineseJournalofEcology, 2016,35(8): 2 014-2 021.
[14] 呼格吉勒图,杨 劼,张 磊,等. 毛乌素沙地低湿地维管植物区系特征[J]. 中国沙漠, 2011,31(5): 1 189-1 194.
HU G J L T, YANG J, ZHANG L,etal. Study on the Flora of Vascular Plants in Low Wetland of the Mu Us Sandy Land[J].JournalofDesertResearch, 2011,31(5): 1 189-1 194.
[15] 赵 爽,高福元,贾 佳,等. 毛乌素沙地风沙沉积物磁学特征及其古环境意义[J]. 地球物理学报, 2015,58(10): 3 706-3 718.
ZHAO S, GAO F Y, JIA J,etal. Magnetic properties of eolian sand sediments in the Mu Us Desert and their paleoen vironmental significance[J].ChineseJournalofGeophysics, 2015,58(10): 3 706-3 718.
[16] 杨梅焕,曹明明,朱志梅. 毛乌素沙地东南缘沙漠化过程中植被的退化和稳定性[J]. 水土保持通报,2017,37(5): 10-15.
YANG M H, CAO M M, ZHU Z M. Vegetation Degradation and Its Stability in Desertification at Southeastern Edge of Mu Us Sandy Land[J].BulletinofSoilandWaterConservation, 2017,37(5): 10-15.
[17] 周淑琴,荆耀栋,张青峰,等. 毛乌素沙地植被空间自相关分布模式及变化特征[J]. 应用基础与工程科学学报,2015,23(2): 318-330.
ZHOU S Q, JING Y D, ZHANG Q F,etal. Vegetation Spatial Autocorrelation Distribution Pattern and Variation Characteristics in Mu Us Sandy Land[J].JournalofBasicScienceandEngineering, 2015,23(2): 318-330.
[18] 中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京:科学出版社,2004.
[19] 内蒙古植物志编辑委员会. 内蒙古植物志(第二版)第四卷[M]. 呼和浩特:内蒙古人民出版社, 1992.
[20] 吴征镒,周浙昆,李德铸,等. 世界种子植物科的分布区类型系统[J]. 云南植物研究,2003,25(3): 245-257.
WU Z Y, ZHOU Z K, LI D Z,etal. The areal-types of the world families of seed plants[J].ActaBotanicaYunnanica, 2003,25(3): 245-257.
[21] 吴征镒,周浙昆,李德铸,彭 华,孙 航. 种子植物分布区类型及其起源和分化[M]. 昆明:云南科技出版社.
[22] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991, (增刊Ⅳ); 1-139.
WU Z Y. The areal-types of Chinese genera of seed plants[J].ActaBotanicaYunnanica, 1991, (Supp.Ⅳ): 1-139.
[23] 李锡文. 中国种子植物区系统计分析[J]. 云南植物研究, 1996,(4): 3-24.
LI X W. Floriatic Statistics and Analyses of Seed Plants from China[J].ActaBotanicaYunnanica, 1996,(4): 3-24.
[24] 王荷生.华北植物区系的演变和来源[J].地理学报,1999,54(3):213-223.
WANG H S. The Evolution and Sources of North China's Flora[J].ActaAgrestiaSinica, 1999,54(3): 213-223.
[25] 布海丽且姆·阿卜杜热合曼,刘会良,刘 娜,等. 天山野果林种子植物区系分析[J]. 干旱区地理, 2016,39(4): 828-834.
BUHLQM ABDRHM, LIU H L, LIU N,etal. Spermatophyte floras in the wild fruit forest in Tianshan Mountains[J].AridLandGeography, 2016,39(4): 828-834.
[26] 王晓莉,吴新宏,师尚礼,等. 浑善达克沙地植物区系分析[J]. 草原与草坪, 2008,(4): 14-19.
WANG X L, WU X H, SHI S L,etal. Analysis of flora in Otindag Sand Land[J].GrasslandandTurf, 2008,(4): 14-19.
[27] 张存厚,刘果厚. 浑善达克沙地种子植物区系分析[J]. 应用生态学报, 2005,(4): 610-614.
ZHANG C H, LIU G H. Spermatophyte flora in Hunshandake sand[J].ChineseJournalofApplifedEcology, 2005,(4): 610-614.
[28] 王继和,袁宏波,张锦春,等. 库姆塔格沙漠植物区系组成及地理成分[J]. 中国沙漠, 2008,(5): 860-867+1 001.
WANG J H, YUAN H B, ZHANG J C,etal. Composition and geographical elements of the flora in Kumtag Desert[J].JournalofDesertResearch, 2008,(5): 860-867+1 001.
[29] 姜春义. 内蒙古库布齐沙漠地区植物多样性及其保护研究[D]. 呼和浩特:内蒙古师范大学, 2016.
[30] 张锦春,刘长仲,姚 拓,等. 乌兰布和沙漠天然植物区系特征及地理成分分析[J]. 草原与草坪, 2013,33(1): 29-33+39.
ZHANG J C, LIU C Z, YAO T,etal. Analysis of floristic characteristics and the geographical elements of natural plants in Ulanbuh Desert[J].GrasslandandTure, 2013,33(1): 29-33+39.
[31] 张巧明,王得祥,龚明贵. 秦岭火地塘种子植物区系及其与毗邻地区的关系分析[J]. 西北植物学报, 2012,32(3): 589-595.
ZHANG Q M, WANG D X, GONG M G. Flora of seed plant in huoditang and its relationship with floras in other areas of Qinling Mountains[J].ActaBot.Boreal.-Occident.Sin, 2012,32(3): 589-595.
[32] 潘晓玲,左家哺. 中国西北地区种子植物区系相似性的定量研究[J]. 西北植物学报, 1997,17(1): 94-102.
PAN X L, ZUO J P. Quantitative study on seed flora resemblance in Northwestern China[J].ActaBot.Boreal.-Occident.Sin, 1997,17(1): 94-102.
[33] 应俊生. 中国种子植物特有属的分布区学研究[J]. 植物分类学报, 1996,340(5): 179-185.
YING J S. Areography of the endemic genera of seed plants in China[J].ActaPhytotaxonomicaSinica, 1996,34(5): 179-185.
猜你喜欢
《学习方法报》地理商务星球七年级(2023年32期)2023-03-13 13:43:26
疯狂英语·新悦读(2022年7期)2022-11-23 00:33:39
金秋(2021年8期)2021-07-27 02:10:48
音乐天地(音乐创作版)(2020年12期)2021-01-11 01:13:46
中国水土保持科学(2019年6期)2019-04-26 05:14:04
世界热带农业信息(2019年11期)2019-01-05 12:18:59
华东师范大学学报(自然科学版)(2017年1期)2017-02-27 13:41:16
安徽农业科学(2015年30期)2015-12-25 07:51:42
南方农业·下旬(2015年9期)2015-10-26 20:27:21
中国粮油学报(2015年5期)2015-02-06 01:47:29
