
不同氮浓度对一株产油绿球藻生长、脂类积累及脂肪酸分布的影响
2018-06-11 06:40:28李涛许瑾吴华莲王铭向文洲
生物技术通报 2018年5期
李涛 许瑾 吴华莲 王铭 向文洲
(1. 中国科学院南海海洋研究所 中国科学院热带海洋生物资源与生态重点实验室,广州 510301;2. 中国科学院广州能源研究所 中国科学院可再生能源与天然气水合物重点实验室,广州 510640;3. 贵州民族大学生态环境工程学院,贵阳550025)
许多门类的微藻在逆境条件下可以大量积累油脂,近10年来,利用微藻油脂生产生物柴油的研究受到国内外广泛关注[1-2]。目前科学家普遍认为微藻生物柴油前景广阔,但现阶段与石化柴油相比缺乏市场竞争力。因此,越来越多的科研工作者将关注点转向微藻油脂的高值化利用,如食品原料、饲料添加剂及化妆品原料等。微藻油脂可以进行高值化开发的依据源于其特殊的脂肪酸组成,微藻可以合成多种具有活性的多不饱和脂肪酸(PUFAs),如α-亚麻酸(C18∶3,ALA),它们在预防心血管疾病、抗癌、促进大脑发育、免疫调节等方面发挥了重要作用[3-4]。
微藻合成的脂类主要有储藏性脂(主要是中性脂类)和膜脂(糖脂和磷脂)两种,PUFAs主要以脂质的形式存在于藻细胞内。膜脂是细胞结构性脂类,含量相对稳定,约占干重的10%;储藏性脂是产油微藻在逆境条件下大量积累的脂类,是细胞能量和碳源的主要储藏形式,可占干重的60%以上[5]。从油脂开发角度分析,储藏性脂相对膜脂更具有优势,原因包括:(1)储藏性脂含量相对于膜脂更高,细胞经过诱导处理后,储藏性脂含量可达到膜脂的3-7倍[6];(2)储藏性脂的流动性好、色泽纯净(橙黄色或浅黄色),而膜脂流动性差、常与叶绿素等混合,色泽暗黑,限制了膜脂产品的开发[7]。因此,微藻油脂高值化开发应该筛选PUFAs更多分布于储藏性脂类中的藻株,或者通过代谢调控的方式,使微藻合成的PUFAs更多用于储藏性脂类合成。
微藻合成的脂酰CoA主要有两种去向:一是,通过“Kennedy途径”合成储藏性脂类;二是,通过“原核途径”和“真核途径”合成膜脂[8]。目前对产油微藻如何分配脂酰CoA用于储藏性脂类和膜脂合成的机制尚不清楚。但已有研究表明,PUFAs在不同脂类分子间的分布受到严格调控。首先,微藻遗传特性影响PUFAs的分布,Reis等[9]发现自养三角褐指藻(Phaeodactylum tricornutum)有84%的二十碳五烯酸(C20:5,EPA)存在于糖脂中,而仅有11%的EPA存在于TAGs(三酰甘油脂)中。Khozin-Goldberg等[10]发现紫球藻(Porphyridium cruentum)可以积累占总脂肪酸47%的EPA,但分布在TAGs中的EPA,仅占2%。Pal等[11]研究表明拟微绿球藻(Nannochloropsissp.)的EPA在TAGs中分布较低。其次,培养条件对PUFAs的分布存在影响。Lu等[12]报道盐度可以影响不同雪衣藻(Chlamydomonas nivalis)不同脂类分子的比例;Řezanka等[13]报道氮磷饥饿可以影响Trachydiscus minutus的EPA在不同脂类分子之间的分配。第三,细胞生长状态影响PUFAs的分布,Tonon等[14]报道眼点拟微绿球藻(Nannochloropsis oculata)、三角褐指藻(P.tricornutum)、假微型海链藻(Thalassiosira pseudonana)和巴夫藻(Pavlova lutheri)的脂肪酸分布不仅受藻种背景影响,而且培养周期对其影响也较大。阐明微藻PUFAs的分布规律,提高PUFAs在中性脂中的比例,对于微藻油脂的高值化开发具有重要的指导作用。
本实验室分离纯化获得了一株产油绿球藻(Chlorococcumsp.),通过前期研究发现,该藻具有生长速率快、油脂含量高、可以积累高含量的α-亚麻酸等特性,具有较高的开发潜力。本研究以产油绿球藻为研究对象,通过测定不同氮浓度条件下的生物质浓度、总脂含量、脂类分级、脂肪酸分布等指标,研究其生长、脂类积累和脂肪酸分布规律,研究结果为高值化开发产油绿球藻油脂提供依据,同时为进一步深入开展产油微藻PUFAs分布调控规律提供基础数据。
1 材料与方法
1.1 材料
1.1.1 藻种 产油绿球藻由中国科学院南海海洋研究所海藻资源与生物技术实验室分离并保藏。
1.1.2 培养方法 微藻培养在光径为3.0 cm、长度为60 cm、有效培养体积为320 mL的柱式光生物反应器中,培养温度为(25±1)℃,荧光灯提供单侧光源,培养光强为300 μmol photons/m2s(微摩尔 光量子/平方米 秒),连续鼓入二氧化碳加富的压缩空气提供碳源和搅拌(CO2∶Air,1∶99)。
1.2 方法
以BG-11培养基为基础培养基[15],设置3个硝酸钠浓度组,分别为3.5 mmol/L、5.9 mmol/L和17.6 mmol/L,每个氮浓度处理组设置3个生物学重复,实验周期为14 d。
将藻种接入硝酸钠浓度为8.8 mmol/L的BG-11培养基中进行扩种培养,待藻细胞生长至对数期(7 d左右),离心收集藻细胞,用无氮的BG-11培养基反复冲洗2次,然后重悬浮于装有不同氮浓度培养基的柱状光生物反应器中,初始接种OD750约为0.5,每2 d测定生物质浓度,第0天、第5天、第10天、第12天、第14天离心收集藻细胞,冻干后测定总脂含量、脂类组成和脂肪酸分布。
1.2.1 生物质浓度 每2 d取藻液V mL,将其过滤到预先烘好的混合纤维滤膜(重量为m1,g)上,将带有藻细胞的滤膜置于80℃烘箱24 h,放置干燥器中冷却至室温后称重(重量为m2,g)。
生物质浓度(g/L)=1000×(m2- m1)/V
1.2.2 总脂含量测定 80 mg藻粉加入3 mL 10% 二甲亚砜-甲醇溶液,分别于50℃抽提30 min、冰浴抽提30 min后,离心收集上清液于预先烘干的玻璃小瓶中,藻渣加入乙醚/正己烷6 mL(1∶1,V∶V),冰浴抽提1 h,离心收集上清液至同一玻璃小瓶中,重复上述过程直到藻渣变白。在合并的抽提液中加入纯水,使二甲亚砜-甲醇、水、乙醚、正己烷的比例为1∶1∶1∶1(V∶V),震荡分相,移取有机相至另一小玻璃瓶中,在通风橱中用氮气吹至较小体积,将浓缩液转移至预先称重过的1.5 mL 塑料离心管中,用氮气吹干至恒重,即得总脂含量[16]。
1.2.3 总脂产量的计算 总脂产量(g/L)=mt× Lt。其中:mt为收获时微藻的生物质浓度(g/L),Lt为收获时微藻的总脂含量(% DW,%干重)。
1.2.4 脂类分级 以一定体积的氯仿-甲醇(1∶1,V∶V)溶解总脂,利用硅胶层析柱(500 mg Sep-PakTMcartridge of silicagel,Waters)进行总脂分级。首先用10 mL的氯仿洗脱得到中性脂(主要为三酰甘油,Triacylglycerol,TAG),然后用10 mL丙酮洗脱得到粗糖脂(Glycolipids,GLs),最后用10 mL甲醇洗脱得到磷脂(Phospholipids,PLs),收集每一组份于小玻璃瓶中,在通风橱中用氮气吹至较小体积,将浓缩液转移至预先称重的1.5 mL 塑料离心管中,用氮气吹干至恒重,得到不同脂类组分的重量[17]。
1.2.5 脂肪酸组成 秤取25 mg藻粉,加入2 mL 2%H2SO4无水甲醇:甲苯(90∶10,V∶V),同时加入25 μL 1%的C17∶0,充氩气后,置于80℃的水浴加热搅拌1.5 h,加入1 mL纯水和正己烷震荡分层,将上层有机相转移到1.5 mL样品瓶中,用氮气吹干,再加入100 μL正己烷密封,利用气相色谱进行脂肪酸含量的测定。气相色谱测定条件为:DB-5毛细管柱(30 m × 0.25 mm),以高纯氮气为载气;检测器为FID检测器;进样口温度260℃,程序升温(60℃保留2 min→30℃/min升温至120℃→1.5℃/min升温至 250℃保留 2 min)[16]。
1.2.6 数据统计与分析 本研究中所有图表所示的平均值和标准偏差均由3个生物学重复和3个测定重复计算获得;利用SPSS 18.0进行数据分析(Analysis of Variance,ANOVA);采用S-N-K方法对不同处理组进行多重比较;样品均值之间的差异用最小显著性差异(Least Significant Difference,LSD)进行分析,置信度为0.05。
2 结果
2.1 氮浓度对产油绿球藻生长的影响
产油绿球藻在3.5 mmol/L、5.9 mmol/L和17.6 mmol/L三种氮浓度培养条件下,生物质浓度随时间的变化曲线如图1所示,产油绿球藻生物质浓度的时相变化呈现延滞期(0-2 d)、对数期(2-10 d)、平台期(10-14 d)3个阶段。在延滞期(0-2 d)和对数期前期(2-4 d),3个氮浓度处理组生物质浓度的变化无明显差异(P>0.05)。从第4天起,3个氮浓度组的生物质浓度出现显著差异(P<0.01),5.9 mmol/L氮浓度组的生物质浓度增长最快,其次为3.5 mmol/L氮浓度组,最慢为17.6 mmol/L氮浓度组,至培养结束时,5.9 mmol/L氮浓度组、3.5 mmol/L氮浓度组和17.6 mmol/L氮浓度组的生物质浓度分别为5.94 g/L、5.85 g/L和4.73 g/L。此外,培养结束时,不同氮浓度组藻液颜色出现明显不同,3.5 mmol/L氮浓度组为橙黄色,5.9 mmol/L氮浓度组为橙色略带绿色,而17.6 mmol/L氮浓度组为黄绿色,藻液颜色的不同表示藻细胞色素组成和含量的差异,色素与细胞光合作用存在直接相关,从而间接反映微藻生物质的积累情况。
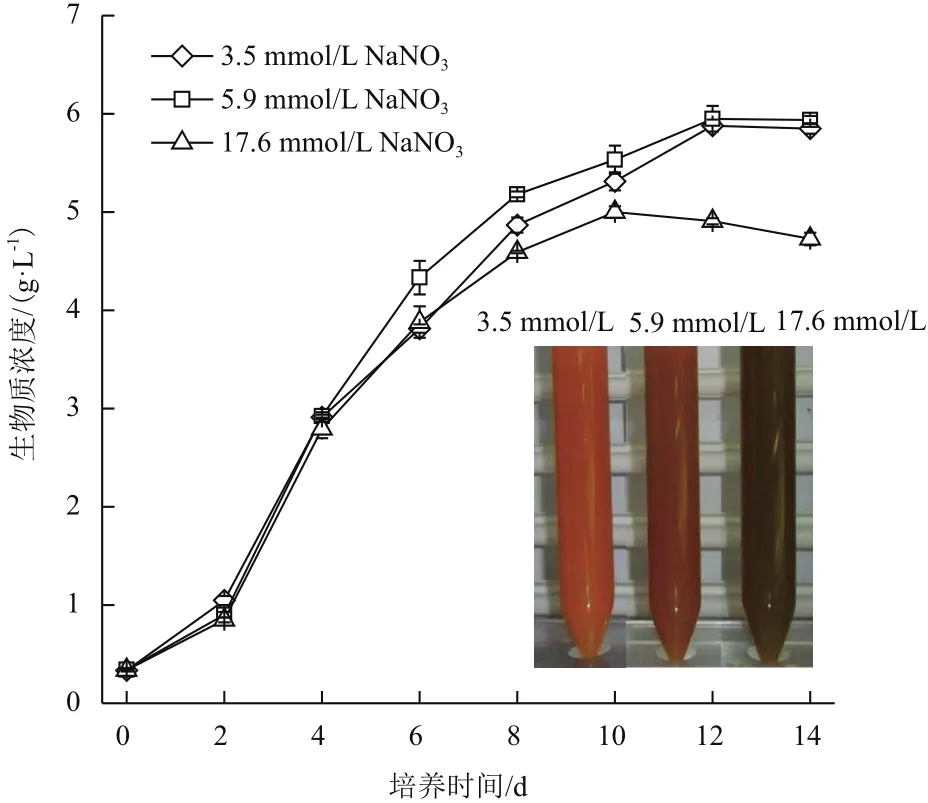
图1 不同氮浓度条件下产油绿球藻(Chlorococcum sp.)生物质浓度的时相变化
2.2 氮浓度对产油绿球藻总脂积累的影响
产油绿球藻在3种氮浓度条件下总脂含量随时间的变化关系如图2所示,随着培养时间的延长,3个氮浓度组的总脂含量均呈现逐渐增加的趋势,总脂积累最快的阶段为第5至第10天。但3个氮浓度组总脂的积累速率存在明显差异(P<0.01),总脂积累最快的组为3.5 mmol/L氮浓度组;其次为5.9 mmol/L氮浓度组,最慢为17.6 mmol/L氮浓度组,至培养结束,3.5 mmol/L氮浓度组的总脂含量达到43.5% DW,中氮组达到42.2% DW,17.6 mmol/L氮浓度组仅有30.3% DW。上述结果说明低氮浓度更有利于微藻油脂的积累。
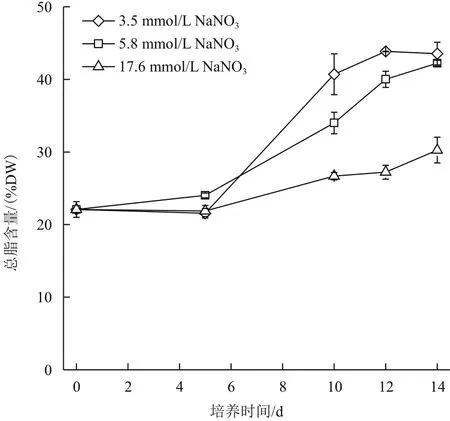
图2 不同氮浓度条件下产油绿球藻总脂含量的时相变化
总脂产量(Total lipid yield)是生物质浓度(g/L)与总脂含量(% DW)的乘积,反映微藻油脂积累的能力。图3为3种氮浓度处理组的总脂产量随时间的变化关系,3种氮浓度处理组的总脂产量随时间延长均表现为增加趋势。至14 d培养结束,3.5 mmol/L氮浓度组和5.9 mmol/L氮浓度组分别取得了2.55 g/L和2.51 g/L的总脂产量(P>0.05),而17.6 mmol/L氮浓度组的总脂收获量仅为1.43 g/L,远低于3.5 mmol/L氮浓度组和5.9 mmol/L氮浓度组(P<0.01)。以上结果说明,培养过程中,过多的氮并不能获得高的总脂产量,而适量的氮会取得更高的总脂产量。
2.3 氮浓度对产油绿球藻脂类组成的影响
微藻脂类可以分为中性脂、糖脂和磷脂3大类,在环境和营养适宜的条件下,微藻合成的脂肪酸主要用于糖脂和磷脂的合成,在逆境条件下,微藻合成的脂肪酸主要用于中性脂的合成,微藻响应不同氮浓度,调整自身脂类组成以适应环境。产油绿球藻在3种氮浓度条件下脂类组成的时相变化如图4所示,培养前期,糖脂和磷脂占较大比例(占总脂的50%以上),随着培养时间的延长,糖脂和磷脂的比例逐渐减少,而中性脂比例逐渐增加。3.5 mmol/L氮浓度组的中性脂积累速率明显高于5.9 mmol/L氮浓度组和17.6 mmol/L氮浓度组,至培养结束,3.5 mmol/L氮浓度组的中性脂比例达到88.6%TL,5.9 mmol/L氮浓度组和17.6 mmol/L氮浓度组分别为86.3% TL和80.5% TL,以上结果说明,低氮浓度有利于中性脂的积累,而对膜脂的合成起抑制作用。
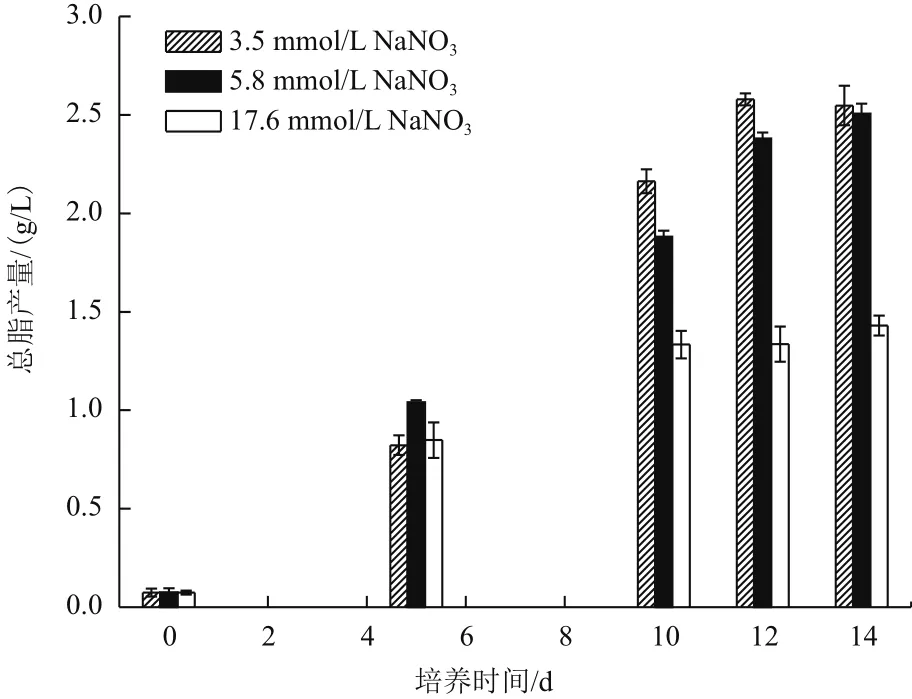
图3 不同氮浓度条件下产油绿球藻总脂产量的时相变化
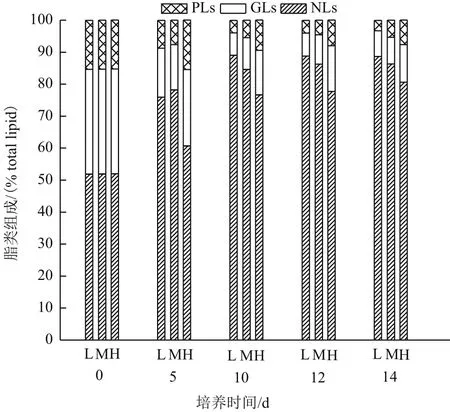
图4 不同氮浓度条件下产油绿球藻脂类组成的时相变化
2.4 氮浓度对产油绿球藻脂肪酸分布的影响
利用气相色谱仪测定了产油绿球藻的脂肪酸组成,其主要脂肪酸包括以下6种,它们分别为C16∶0、C16∶1、C18∶0、C18∶1、C18∶2 和C18∶3(ω-3)。随后测定了上述6种脂肪酸在中性脂、糖脂和磷脂中的分布情况。结果表明,6种主要脂肪酸在中性脂、糖脂和磷脂中均有分布,但在中性脂、糖脂和磷脂中的比例各不相同。以培养第0天为例,C16∶0、C16∶1、C18∶0、C18∶1 和 C18∶2 在中性脂中的比例分别为52.6%、62.1%、52.0%、62.4%和53.7%,均超过50%以上,而C18∶3(ω-3)仅有33.9%分布于中性脂中。随着培养时间的延续,6种主要脂肪酸在中性脂和膜脂中的分布比例发生明显变化,均呈现出在中性脂中比例增加,而在膜脂中比例减少的趋势,至培养结束,低氮组C16∶0、C16∶1、C18∶0、C18∶1、C18∶2 和 C18∶3(ω-3)在中性脂中的比例分别提高到85.6%、90.8%、84.7%、95.2%、88.3%和86.5%。
氮浓度对6种脂肪酸的分布具有一定影响,随着氮浓度降低,C16∶0、C16∶1、C18∶1、C18∶2和C18∶3在中性脂中的比例显著增加,而在膜脂中的比例逐渐减少,仅有C18∶0在中性脂和膜脂中的比例不受氮浓度影响。
3 讨论
一些门类的微藻可以合成具有特殊生物学活性的PUFAs,是开发高值微藻油的理想原料[18]。微藻合成的脂类主要有储藏性脂和膜脂两种[2],PUFAs存在于上述两种脂类中。储藏性脂具有流动性好、色泽清澈、含量高等特点,相对于膜脂应用范围更广、开发优势更显著[7]。因此,筛选PUFAs更多分布于储藏性脂类中的藻株是微藻油高值化开发的关键环节。本实验室分离纯化获得了一株产油绿球藻,该藻具有生长速率快、生物量高、可积累高含量油脂和α-亚麻酸等特性,存在潜在的开发利用价值。本研究以该株产油绿球藻为对象,研究不同氮浓度对其生长、脂类积累和脂肪酸分布规律的影响,为高值微藻油的开发奠定基础。
在诸多影响微藻油脂积累的因素中,氮元素对微藻油脂积累的影响最为明显,大量研究表明,低氮限制可以提高微藻油脂含量,但对微藻正常分裂生长存在严重抑制[19]。本研究设置17.6 mmol/L、5.9 mmol/L和3.5 mmol/L三种氮浓度,其中,17.6mmol/L为正常BG-11培养基中硝酸钠的浓度。前期研究发现,产油绿球藻(Chlorococcumsp.)在17.6 mmol/L氮浓度条件下可以保持较快分裂速率,对藻细胞的正常生长并未产生抑制,因此设置17.6 mmol/L为氮充足组。而当浓度降低为5.9 mmol/L和3.5 mmol/L后,培养前期藻细胞数量有一定增加,但随着氮元素的快速消耗,藻细胞很快进入氮限制状态(4-6 d左右),细胞停止分裂并开始大量合成油脂,故设置5.9 mmol/L和3.5 mmol/L为氮限制组。此外,藻细胞在3.5 mmol/L氮浓度条件下较5.9 mmol/L更早进入氮限制状态,因此,设置两种氮限制处理组,深入探讨不同低氮限制浓度对微藻脂类积累、脂类组成和脂肪酸分布的潜在影响。结果显示,3.5 mmol/L和5.9 mmol/L氮浓度条件下,微藻的生物质浓度明显高于17.6 mmol/L氮浓度组,这一结果与已有的文献报道相矛盾[19-21]。从上述结果是否可以得出高氮浓度限制微藻正常生长这一结论,答案是否定的。藻细胞分裂情况显示,中氮和低氮条件下,藻细胞分别在第4天和第6天已停止分裂,而高氮条件第12天仍然观察到细胞分裂现象(研究结果未在本文中列出),说明高氮并未抑制微藻正常分裂生长。总脂积累情况显示:3.5 mmol/L和5.9 mmol/L氮浓度组的总脂积累速率明显高于17.6 mmol/L氮浓度组。由此推测:微藻细胞在3.5
mmol/L和5.9 mmol/L氮浓度条件下更早进入氮限制,氮限制抑制细胞正常分裂,但氮限制却激活细胞能量和碳源的快速储存途径,使微藻生物质快速积累,而在17.6 mmol/L氮浓度条件下,细胞将更多的能量和碳源用于其他非储存型代谢活动,如细胞分裂等,并未用于储藏物的快速积累。此外,观察到培养过程中17.6 mmol/L氮浓度条件下藻液颜色始终比3.5 mmol/L和5.9 mmol/L氮浓度组更绿,颜色绿表明叶绿素浓度高。叶绿素含量通常与光合效率呈正相关[22],17.6 mmol/L氮浓度组的光合效率高于3.5 mmol/L和5.9 mmol/L氮浓度组,但有趣的是17.6 mmol/L氮浓度组的生物质积累量却远低于3.5 mmol/L和5.9 mmol/L氮浓度组,这一结果同样印证了前述推测“3.5 mmol/L和5.9 mmol/L氮浓度组进入低氮限制后,在碳代谢和能量代谢途径上可能存在较大的改变,使细胞为适应逆境而进行能量和碳源的储存”。本文作者在2016年发表的文章对这一现象进行了详细的解释,并提出通过初始氮调控的方式可以获得更高油脂产量的培养模式[6]。
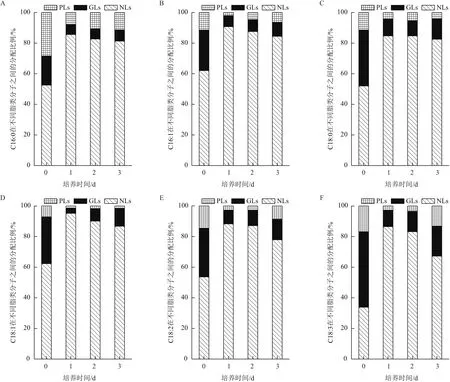
图5 产油绿球藻主要脂肪酸在不同脂类中的分布
总脂产量是生物质浓度与总脂含量的乘积,反映微藻油脂积累的实际能力[5],本研究中3.5 mmol/L和5.9 mmol/L氮浓度组分别取得了2.55 g/L和2.51 g/L的总脂产量,比17.6 mmol/L氮浓度组提高了78.3%和75.5%。深入比较发现3.5 mmol/L和5.9 mmol/L氮浓度组的生物质浓度和总脂含量均高于17.6 mmol/L氮浓度组。产生这一结果的原因是给予细胞少量的初始氮,微藻利用少量的氮源进行分裂和生长,达到一定的细胞密度,当培养基中氮源耗尽后,细胞进入氮限制状态,开始大量合成储藏性脂类,从而到收获时达到较高的总脂产量(自然两步法)。如果延长17.6 mmol/L氮浓度组的培养时间,是否也可以获得更高的总脂产量?或者给予一个更加合适的氮浓度,是否可以将总脂产量得到更大幅度的提高,这部分优化内容本文作者将在后续的工作中开展。
微藻油脂组成对其下游开发应用极为重要,膜脂通常与叶绿素混合,使油呈现黑褐色,将严重影响油的品质,大幅增加微藻油脱色和精炼的成本[7]。因此,了解微藻脂类组成的变化规律,改变培养条件,使微藻通过自身的代谢调控,减少膜脂比例,增加中性脂比例,具有非常重要生产意义[23]。本研究结果显示,随着培养时间的延长,中性脂比例呈现增加的趋势,低氮条件下的中性脂增速更快、比例更高。上述结果与已有研究结果一致。王金娜[24]报道假微型海链藻(T.pseudonana)、蛋白核小球藻(Chlorellapyrenoidosa)和绿色巴夫藻(P.viridis)的中性脂在指数生长后期或平台期开始大量积累。周芷薇[25]报道低氮可以大幅提到魏氏藻中性脂的比例。此外,藻细胞降解的膜脂可以为中性脂的合成提供碳源,以磷脂作为酯酰供体,通过磷脂:二酰甘油酰基转移酶(PDAT)催化形成三酰甘油,以糖脂作为酯酰供体,通过半乳糖脂:半乳糖脂半乳糖基转移酶(GGGT)催化单半乳糖甘油二酯中的半乳糖基转移到单半乳糖甘油二酯或双半乳糖甘油二酯上,在此过程中产生的二酰甘油是三酰甘油合成的前体[26-27]。叶绿体的类囊体膜是许多色素蛋白复合物的结合位点,如果膜脂大量降解,细胞光合固碳工厂将会停止工作,导致细胞最终衰亡。
微藻合成的PUFAs可以分布于中性脂类中或膜脂中,判断一株产油藻株是否具有高值化微藻油的开发潜力,需要分析其PUFAs在储藏性脂类中或膜脂分布情况。大量研究表明,培养条件(温度、光强、盐度、氮元素及磷元素)对微藻的脂肪酸组成存在一定影响,那么培养条件是对PUFAs的分布是否也存在影响,Lu等[12]年报道盐度可以影响雪衣藻(C.nivalis)不同脂类分子的比例;Řezanka 等[13]报道氮磷饥饿可以影响T.minutus影响EPA在不同脂类分子之间的分配。本研究所用藻株产油绿球藻的脂肪酸组成为 C16∶0、C16∶1、C18∶0、C18∶1、C18∶2和C18∶3(ω-3),随着培养时间的延续,脂肪酸在中性脂中比例增加,而在膜脂中比例减少。此外,随着氮浓度降低,上述6种脂肪酸在中性脂的比例也显著增加,上述结果与中性脂的变化规律保持一致。推测:中性脂的快速合成,需要更多的脂肪酸作为供体,微藻调控合成的脂肪酸进入中性脂合成途径,膜脂降解产生的脂肪酸也可能为中性脂的合成提供供体[26-27]。有研究报道这种分配调控机制可能与不同脂类在细胞中合成位点有关,中性脂的合成主要在内质网,而膜脂的合成主要发生在叶绿体中[8],当氮缺乏导致叶绿体膜大量降解后,其上的脂类合成酶也会随之分解,导致膜脂无法合成,脂肪酸转而被分配至中性脂合成途径[8]。产油绿球藻可以合成具有生物学活性的α-亚麻酸,本研究表明,在氮充足的条件下,α-亚麻酸分布在膜脂中,而在氮胁迫后,α-亚麻酸会更多的分布于中性脂中,中性脂含量增加,α-亚麻酸的含量会相应增加,上述特性使该藻株具有高值化油脂开发潜力。此外,一些具有开发价值的多不饱和脂肪酸,如二十碳五烯酸(EPA)主要分布于膜脂上,如拟微绿球藻、三角褐指藻、紫球藻等[9-11],如果阐明EPA的分布调控机制,在后续的工作中利用基因改造或者定向诱导的方式,使EPA倾向于分布于储藏性脂类中,这对于EPA微藻油开发具有非常重要的价值。
4 结论
适宜氮浓度对于提高产油绿球藻总脂产量、中性脂的比例、多不饱和脂肪酸分布具有重要作用。产油绿球藻积累的α-亚麻酸在低氮条件下倾向于分布在中性脂中,因此它是一株具有高值化微藻油开发价值的藻株。
[1] Wijffels RH, Barbosa MJ. An outlook on microalgal biofuels[J].Sci, 2010, 329(5993):796-799.
[2] Hu Q, Sommerfeld M, Jarvis E, et al. Microalgal triacylglycerols as feedstocks for biofuel production :perspectives and advances[J].Plant J, 2008, 54(4):621-639.
[3] Spolaore P, Joannis-Cassan C, Duran E, et al. Commercial applications of microalgae[J]. J Biosci Bioeng, 2006, 101(2):87-96.
[4] 李春颖, 仇雪梅. 海洋微藻脂肪酸组成的研究进展[J]. 生物技术通报, 2008(4):63-65.
[5] Rodolfi L, Chini Zittelli G, Bassi N, et al. Microalgae for oil:strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor[J]. Biotechnol Bioeng, 2009, 102(1):100-112.
[6] Li T, Xu J, Gao B, et al. Morphology, growth, biochemical composition and photosynthetic performance ofChlorella vulgaris(Trebouxiophyceae)under low and high nitrogen supplies[J].Algal Research, 2016, 16:481-491.
[7] Li T, Xu J, Wu H, et al. A saponification method for chlorophyll removal from microalgae biomass as oil feedstock[J]. Marine Drugs, 2016, 14(9):162-181.
[8] Heldt HW. Plant Biochemistry 3th edition[M]. San Diego:Elsevier Academic Press, 2005.
[9] Reis A, Gouveia L, Veloso V, et al. Eicosapentaenoic acid-rich biomass production by the microalgaPhaeodactylum tricornutumin a continuous-flow reactorv[J]. Bioresource Technol, 1996, 55(1):83-88.
[10] Khozin-Goldberg I, Yu H Z, Adlerstein D, et al. Triacylglycerols of the red microalgaPorphyridium cruentumcan contribute to the biosynthesis of eukaryotic galactolipids[J]. Lipids, 2000, 35(8):881-889.
[11] Pal D, Khozin-Goldberg I, Cohen Z, et al. The effect of light,salinity, and nitrogen availability on lipid production byNannochloropsissp. [J]. Apply Microbiol Biotechnol, 2011, 90(4):1429-1441.
[12] Lu N, Wei D, Chen F, et al. Lipidomic profiling and discovery of lipid biomarkers in snow algaChlamydomonas nivalisunder salt stress[J]. Eur J Lipid Sci Technol, 2012, 114(3):253-265.
[13] Řezanka T, Lukavský J, Nedbalová L, et al. Effect of nitrogen and phosphorus starvation on the polyunsaturated triacylglycerol composition, including positional isomer distribution, in the algaTrachydiscus minutus[J]. Phytochemistry, 2011, 72(18):2342-2351.
[14] Tonon T, Harvey D, Larson T R, et al. Long chain polyunsaturated fatty acid production and partitioning to triacylglycerols in four microalgae[J]. Phytochemistry, 2002, 61(1):15-24.
[15] Andersen R A. Algal culturing techniques[M]. London:Elsevier Academic Press, 2005.
[16] Khozin-Goldberg I, Shrestha P, Cohen Z. Mobilization of arachidonyl moieties from triacylglycerols into chloroplastic lipids following recovery from nitrogen starvation of the microalgaParietochloris incise[J]. BBA-Mol Cell Biol Lipids, 2005, 1738(1-3):63-71.
[17] Christie WW. Lipid analysis:Isolation, separation, identification and structural analysis of lipids[M]. Oxford:Pergamon Press,1982.
[18] Lang I, Hodac L, Friedl T, et al. Fatty acid profiles and their distribution patterns in microalgae:a comprehensive analysis ofmore than 2000 strains from the SAG culture collection[J]. BMC Plant Biology, 2011, 11(1):124-140.
[19] Breuer G, Lamers PP, Martens DE, et al. The impact of nitrogen starvation on the dynamics of triacylglycerol accumulation in nine microalgae strains[J]. Bioresource Technol, 2012, 124:217-226.
[20] Dean AP, Sigee DC, Estrada B, et al. Using FTIR spectroscopy for rapid determination of lipid accumulation in response to nitrogen limitation in freshwater microalgae[J]. Bioresource Technol,2010, 101:4499-4507.
[21] Adams C, Godfrey V, Wahlen B, et al. Understanding precision nitrogen stress to optimize the growth and lipid content tradeoff in oleaginous green microalgae[J]. Bioresource Technol, 2013,131:188-194.
[22] 韩博平, 韩志国, 付祥. 藻类光合作用机理与模型[M]. 北京:科学技术出版社, 2003.
[23] 姜悦, 陈峰. 利用海洋微藻培养生产 ω-3 多不饱和脂肪酸[J].海洋科学, 1997, 21(6):18-20.
[24] 王金娜, 严小军, 周成旭, 等. 产油微藻的筛选及中性脂动态积累过程的检测[J]. 生物物理学报, 2010, 26(6):472-480.
[25] 周芷薇, 高保燕, 雷学青, 等. 低氮胁迫对两种魏氏藻生长和油脂积累的影响[J]. 可再生能源, 2015, 33(5):777-783.
[26] Yoon K, Han D, Li Y, et al. Phospholipid:Diacylglycerolacyltransferase is a multifunctional enzyme involved in membrane lipid turnover and degradation while synthesizing triacylglycerol in the unicellular green microalgaChlamydomonas reinhardtii[J].Plant Cell, 2012, 24(9):3708-3724.
[27] Benning C, Ohta H. Three enzyme systems for galactoglycerolipid biosynthesis are coordinately regulated in plants[J]. J Biol Chem, 2005, 280(4):2397-2400.
猜你喜欢
现代临床医学(2023年1期)2023-03-24 08:30:20
粮食与食品工业(2021年3期)2021-06-18 07:50:48
化工设计通讯(2021年2期)2021-03-15 07:03:30
中国石油石化(2020年12期)2020-07-30 05:10:24
水生生物学报(2019年3期)2019-05-29 06:58:48
广东茶业(2019年1期)2019-04-28 08:32:30
中国油脂(2017年7期)2017-09-16 04:28:48
东方考古(2016年0期)2016-07-31 17:45:44
中国医药科学(2015年15期)2015-02-27 12:32:15
水生生物学报(2014年2期)2014-05-27 08:07:30
