
不同北美海棠品种对低温胁迫的生理响应
2018-06-05 05:56阿地力衣克木巴哈尔古丽阿尤甫哈尔肯叶尔江阿依古丽铁木儿
西南农业学报 2018年4期
阿地力·衣克木,巴哈尔古丽·阿尤甫,哈尔肯·叶尔江,阿依古丽·铁木儿
(新疆林业科学院,新疆 乌鲁木齐 830063)
【研究意义】海棠(Malusspp.)属蔷薇科苹果属(MalusMill.)观赏树种,品种众多,集叶、花、果、树形共赏。海棠起源于我国,具有品种资源丰富、观赏价值高、环境适应性强、地里分布广泛等特性[1-3]。18 世纪以来,欧美等国家的园艺工作者在从中国大量引种和杂交选育的基础上,培育出一些观花、观果的海棠品种,一般称为北美海棠[4]。该树种以其优美的树形,丰富多彩的观赏效果,成为当前国内各城市绿化的流行树种。【前人研究进展】2008年以来,新疆也引进了一系列优良品种,并开展了北美海棠的引种、驯化[5-6]等工作。不同地区间引种时,常会遇到对气候不适应的现象。北疆地区冬天比较寒冷,低温胁迫是经常遇到的一种灾害,在低温下植物会产生不同程度的伤害,寒冷作用于机体,所致损伤或死亡[7]。为了避免北美海棠“南树北引”过程中出现盲目性。【本研究切入点】本试验通过研究不同北美海棠品种对低温胁迫的生理响应,从而了解其抗寒生理特性,评价抗寒能力,筛选抗寒品种。【拟解决的关键问题】以期为北美海棠的引种驯化和抗寒育种实践提供依据。
1 材料与方法
1.1 试验材料
2014年,从辽宁省铁岭市西丰县云竹林场引进北美海棠优良品种,高原之火(M.‘Prairifire’)、印度安魔力(M.‘Indian Magic’)、粉手帕(M.‘fenshoupa Begonia’)、亚当(M.‘Adams’)、金色海棠(M.‘gold Begonia’)、牡丹海棠(M.‘peony Begonia’)、红叶海棠(M. ‘micro maluscv’)、绚丽(M.‘Radiant’)、草莓果冻(M.‘Strawberry Parfait’)、红宝石(M.‘Red Jew elberry’) 等 10 个品种。10个供试材料的均保存于新疆林业科学院玛纳斯试验基地。该基地位于玛纳斯县,玛纳斯县位于新疆维吾尔自治区中北部,昌吉州最西部,总面积1.1×104km2。地理坐标为东经85°34′~86°43′,北纬43°28′~45°38′,地处内陆,远距海洋,气候干旱,降雨稀少,属暖温带极端干旱荒漠气候。降雨量稀少,蒸发量大,昼夜温差悬殊。年均气温10.01 ℃,极端最高气温40.9 ℃,极端最低气温-37.4 ℃,太阳辐射年均总量140 kcal /cm2,年均日照2727.2 h,年均降水量65.4 mm,年均蒸发量2002.2 mm,年均无霜期185 d。全年盛行西风,最大风力八级,多出现在3-5月,平均大风日数为 32 d,每年春季大风多拌有沙尘及寒潮天气。
1.2 试验方法
1.2.1 取样方法 于2017年2月8日取样,每品种选取生长健壮,树冠外围发育正常、长势良好、粗细均匀一致的1年生休眠枝条50 条,剪成 30 cm 左右的小段,用报纸包好,带回实验室。
1.2.2 低温胁迫处理 将选取好的枝条洗净后剪成长短相对一致(长30 cm 左右)的短枝,先用自来水冲洗干净,然后用去离子水冲洗 2 遍,用吸水纸吸干水分,然后将每种枝条粗细均匀一致地分成5份,包上报纸,打湿后用塑料膜包裹,放在0 ℃冰箱里备用。将其中4份分别在-25、-30、-35和-40 ℃的低温冰箱进行24 h 低温处理。对照为在室外自然越冬的枝条,试验地当时最低温度为-20 ℃。低温处理后在室内进行各指标的测定,重复 3 次。
1.2.3 相关指标的测定方法 ①参照电导率法[8]。将低温冷冻处理后的枝条剪成2~3 mm 的小段,然后称取 2 g 试样放入三角瓶,并加入50 mL 蒸馏水,浸泡24 h 后,测定浸出液的电导率(重复3次)。然后放在水浴锅中煮沸1 h,静止冷却后测定其电导度。相对电导率Y( %)=初电导值/终电导值×100。②生理指标的测定。可溶性蛋白测定参照考马斯亮蓝G-250染色法[9],可溶性糖测定参照蒽酮比色法[9],丙二醛(MDA)含量测定参照硫代巴比妥酸(TBA)法[10]。
1.3 数据分析
采用统计软件SPSS16.0和Excel2003进行数据处理和统计分析。
不同北美海棠品种抗寒性综合评价。应用隶属函数法对不同北美海棠品种抗寒性进行综合评判。①与抗寒性呈正相关的参数以下公式计算:U(X)=(Xij-Xmin)/(Xmax-Xmin)。②与抗寒性呈负相关的参数以下公式计算:U(X)=1-(Xij-Xmin)/(Xmax-Xmin),公式中:U(X)为抗寒指标的隶属度,且U(X)∈[0,1];Xij为各指标的测定值;Xmax、Xmin为各指标的最大值和最小值。以每一种各项指标隶属度的平均值作为品种抗寒能力综合评判标准,进行比较。
2 结果与分析
2.1 低温胁迫对不同北美海棠品种相对电导率的影响
由表1可以看出,随着处理温度的降低,不同北美海棠品种枝条相对电导率均呈现“S”型上升趋势。其中,印度安魔力枝条相对电导率上升最快,为82.572 %,比对照(-20 ℃)上升了20.808 %,并与对照(-20 ℃)呈极显著差异(P≤0.01)。高原之火、亚当、牡丹海棠、绚丽、草莓果冻、红宝石枝条相对电导率上升较快,分别为86.624 %、84.439 %、79.818 %、82.333 %、86.206 %、83.026 %,比对照(-20 ℃)上升了17.789 %、18.484 %、16.504 %、18.273 %、18.586 %、19.258 %,并与对照(-20 ℃)呈极显著差异(P≤0.01) 粉手帕、金色海棠、红叶海棠枝条相对电导率上升较缓慢。为81.542 %、80.957 %、79.122 %,比对照(-20 ℃)上升了14.613 %、14.950 %、11.826 %,并与对照(-20 ℃)呈极显著差异(P≤0.01)。
2.2 低温胁迫对不同北美海棠品种枝条可溶性糖含量的影响
由表2可以看出,随着处理温度的降低,不同北美海棠品种枝条可溶性糖含量均呈现先上升后下降趋势,并在-20~-35 ℃范围内可溶性糖含量的变化一定程度上体现了各品种抗寒能力,其中,粉手帕、亚当、金色海棠、绚丽、红宝石可溶性糖含量在-30℃的敏感性较高,其中粉手帕和亚当与-35 ℃不存在差异,与其它处理呈极显著差异(P≤0.01);金色海棠、绚丽、红宝石呈极显著差异(P≤0.01)。高原之火、印度安魔力、牡丹海棠、红叶海棠、草莓果冻可溶性糖含量在-35 ℃的敏感性较高,其中高原之火、印度安魔力、牡丹海棠、红叶海棠与其它处理呈极显著差异(P≤0.01);草莓果冻与-30 ℃不存在差异,与其它处理呈极显著差异(P≤0.01)。
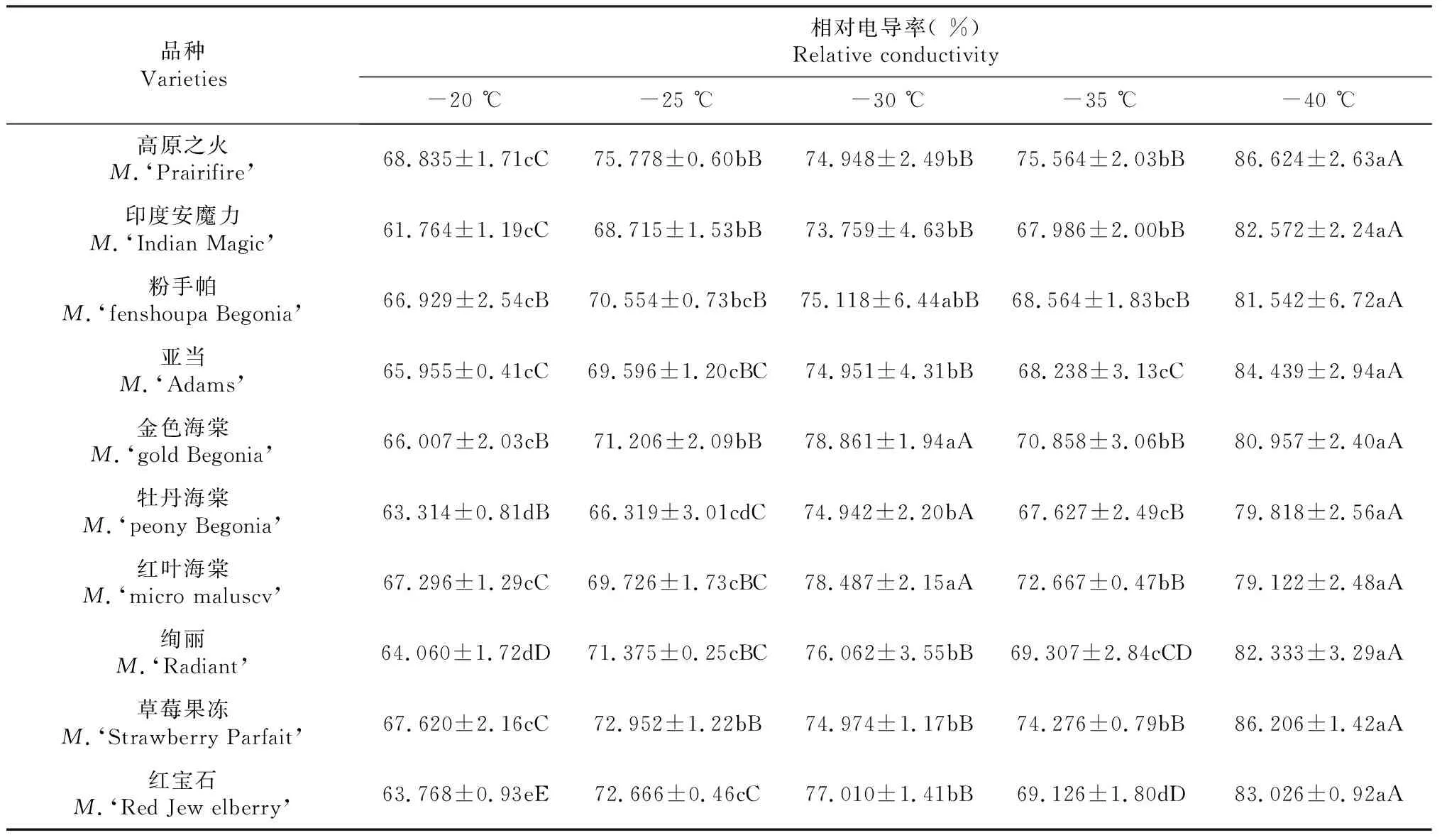
表1 低温胁迫对不同北美海棠品种相对电导率的影响
注:同行不同小写字母表示(P≤0.05)差异显著性;同行不同大写字母表示(P≤0.01)差异极显著性。下同。
Note: The small letter means a significant difference at 0.05 level; The capital means a significant difference at 0.01 level. The same as below.
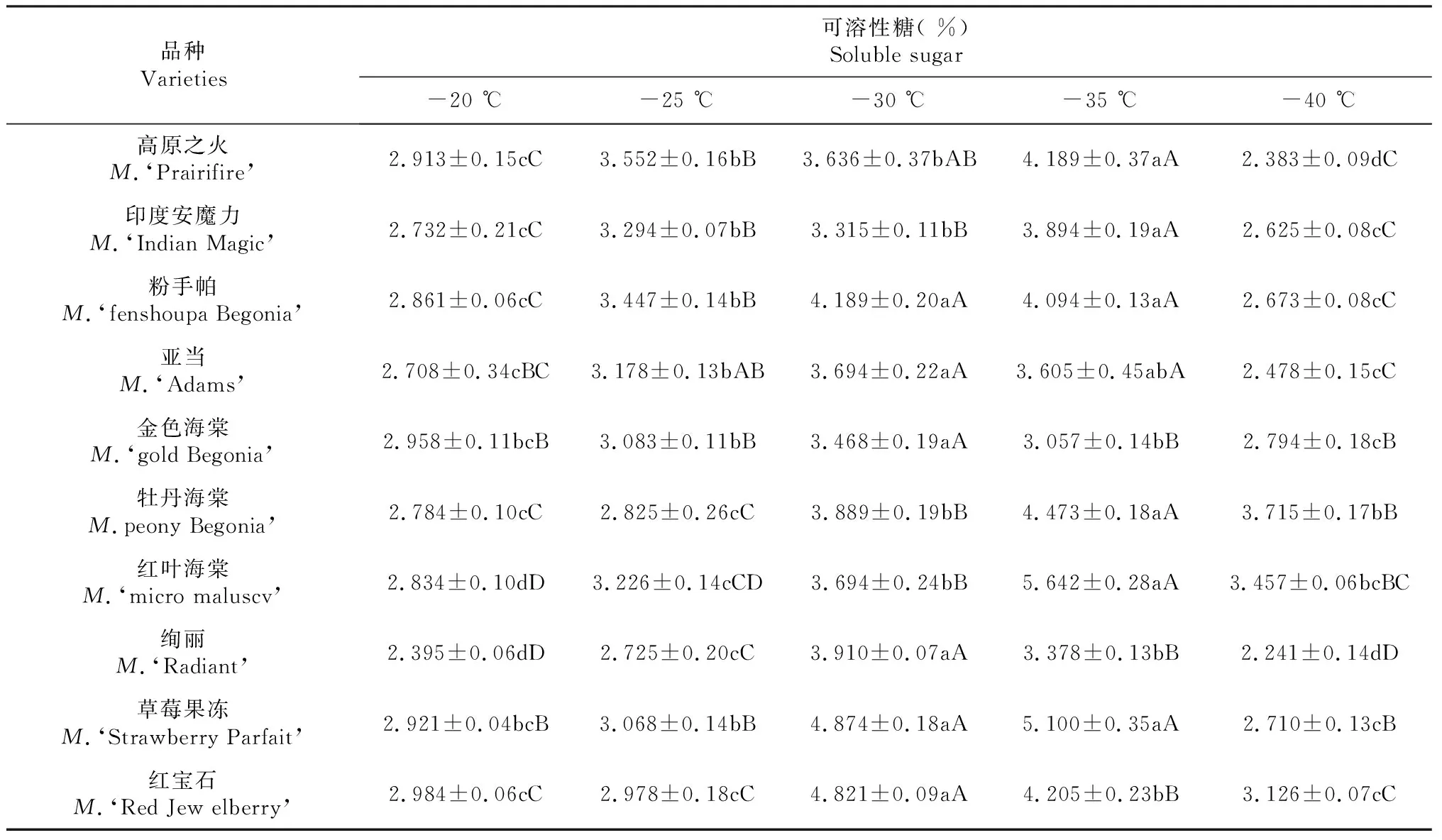
表2 低温胁迫对不同北美海棠品种枝条可溶性糖含量的影响
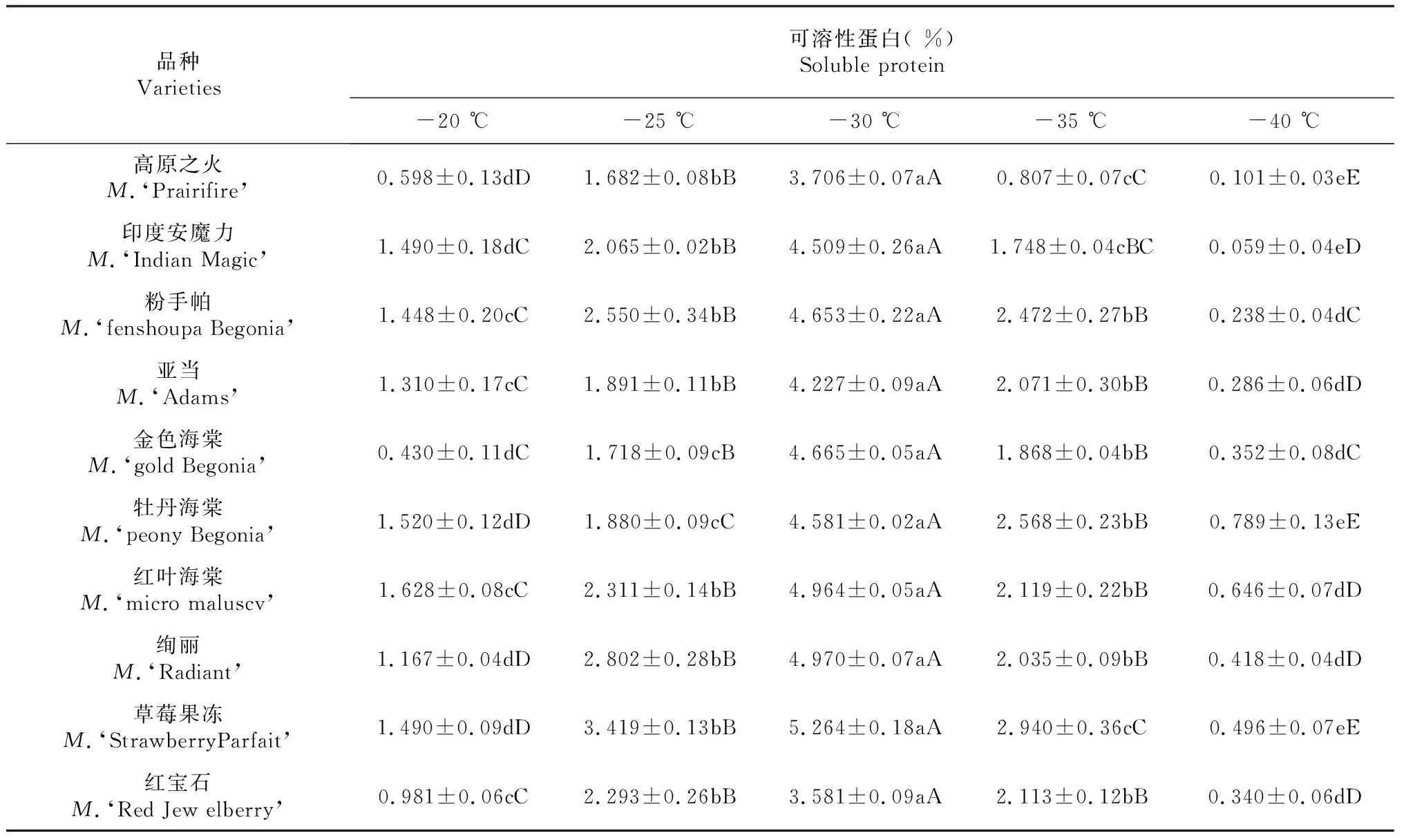
表3 低温胁迫对不同北美海棠品种可溶性蛋白含量的影响
2.3 低温胁迫对不同北美海棠品种枝条可溶性蛋白的影响
由表3可以看出,随着处理温度的降低,不同北美海棠品种枝条可溶性蛋白含量均呈现先上升后下降趋势。在-20~-30 ℃范围内可溶性蛋白的变化较准确的体现了各品种抗寒性,并在-30 ℃可溶性蛋白的敏感性较高,并与其它处理呈极显著差异(P≤0.01)。
2.4 低温胁迫对不同北美海棠品种枝条丙二醛的影响
由表4可以看出,随着处理温度的降低,不同北美海棠品种枝条丙二醛含量呈先上升后下降趋势。高原之火和亚当在-30~-35 ℃,其它品种都在-20~-35 ℃范围内丙二醛的变化一定程度上体现了各品种抗寒能力。其中,牡丹海棠和红叶海棠在-25 ℃的敏感度较高并与其他处理呈极显著差异(P≤0.01);粉手帕、绚丽、红宝石在-30 ℃的敏感度较高,其中粉手帕与对照(-20 ℃)呈极显著差异(P≤0.01),与其他处理不存在差异;绚丽和红宝石与-25 ℃不存在差异,其他处理呈极显著差异(P≤0.01)。高原之火、印度安魔力、亚当、金色海棠、草莓果冻在-35 ℃的敏感度较高,其中,高原之火与对照(-20 ℃)呈显著差异(P≤0.05),与-25 ℃不存在差异,与-30和-40 ℃呈极显著差异(P≤0.01);印度安魔力和粉手帕与对照(-20 ℃)呈极显著差异(P≤0.01),与-25、-30 ℃不存在差异,印度安魔力在-40 ℃呈极显著差异(P≤0.01),粉手帕在-40 ℃不存在差异;亚当与-20、-25 ℃之间不存在差异,与-30、-40 ℃之间存在极显著差异(P≤0.01);金色海棠与对照(-20 ℃)呈极显著差异(P≤0.01),与-25 ℃之间不存在差异,与-30、-40 ℃之间存在显著差异(P≤0.05);草莓果冻与对照(-20 ℃)、-25 ℃之间存在极显著差异(P≤0.01),与-30 ℃之间存在显著差异(P≤0.05),与-40 ℃之间不存在差异。
2.5 不同北美海棠品种抗寒性的综合评判
由表5可以看出,不同北美海棠品种的平均隶属度0.441~0.564,红叶海棠的平均隶属度为最高,为0.564,粉手帕的平均隶属度最低,为0.441。平均隶属度显示,不同北美海棠品种的抗寒性排序为:红叶海棠>金色海棠>绚丽>红宝石>高原之火>印度安魔力>牡丹海棠>亚当>草莓果冻>粉手帕。
3 讨 论
低温胁迫是影响植物生长、发育和地理分布的重要环境限制因素之一。低温胁迫对植物的影响主要体现在酶活性、膜系统、细胞失水等,导致细胞代谢紊乱,甚至是细胞死亡[11]。温度的改变则引起植物的各种生理生化指标的改变来抵抗环境温度的胁迫。主要抗寒性指标有相对电导率、可溶性糖、可溶性蛋白、脯氨酸、丙二醛、酶活性等,这些指标虽然不能直接判断品种间抗寒性强弱,但都与抗寒性有密切的关系[12-13]。植物受低温伤害时,膜的选择透性减弱,膜内大量溶质外渗,可引发植物代谢失调。具体反映在植物浸出液的电导率增加[14-15]。植物在低温锻炼的过程中,淀粉水解为可溶性糖,这是细胞原生质的重要保护物质。由于糖含量的增加,冰点降低,缓解原生质的脱水过程,保护了原生质胶体免于凝聚[16]。可溶性蛋白含量随低温锻炼过程抗寒性的提高而增加,可溶性蛋白质含量的增加,可显著加强细胞的保水力,同时增加细胞内束缚水的含量,对发展高水平的抗冻性有重要作用[17-18]。人们发现植物在逆境下遭受伤害时,往往发生膜脂过氧化作用,丙二醛(MDA)是膜脂过氧化的最终产物,对质膜有毒害作用,丙二醛的积累可能对膜和细胞造成一定的伤害[19]。其含量可以反映植物遭受逆境伤害的程度。
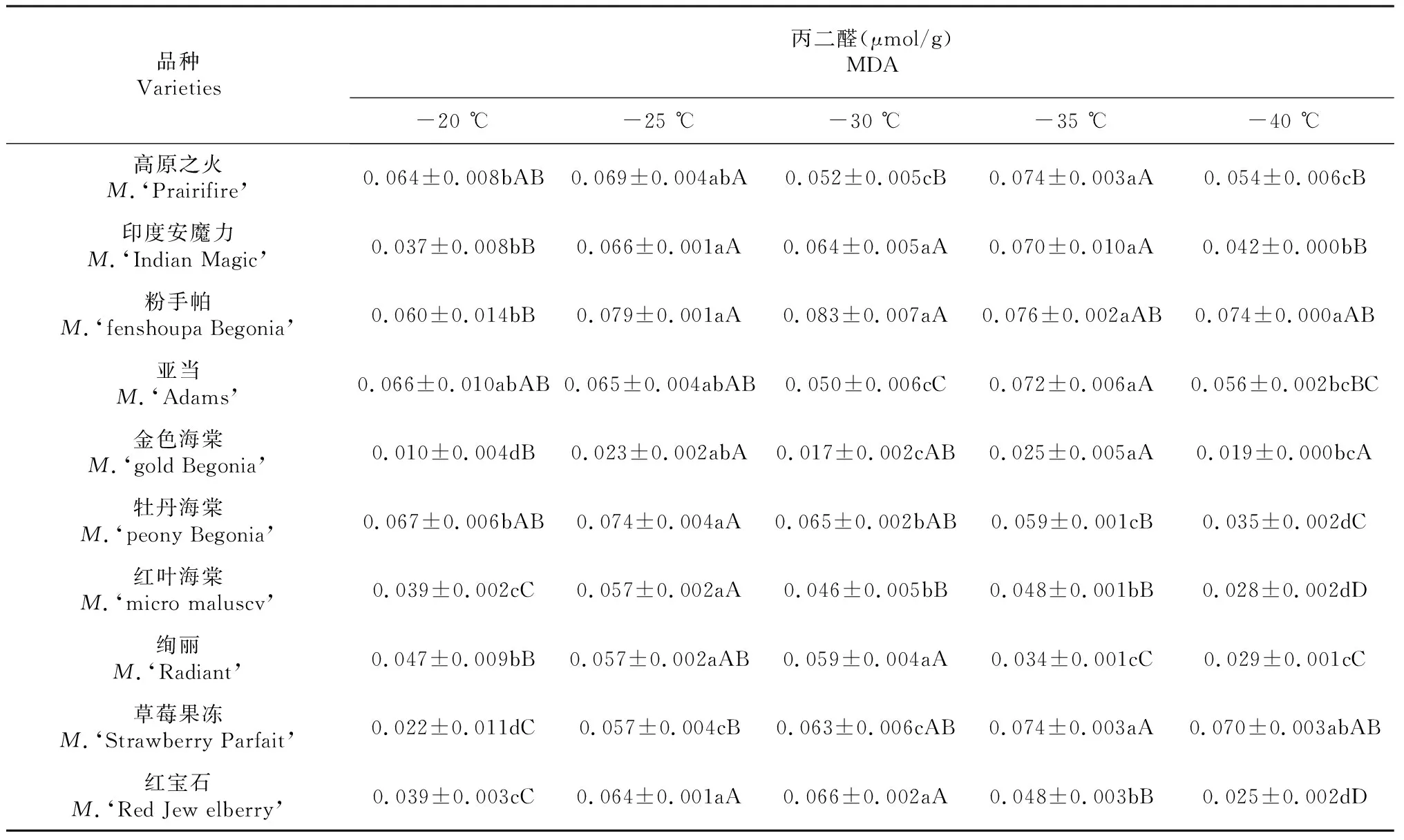
表4 低温胁迫对不同北美海棠品种丙二醛含量的影响
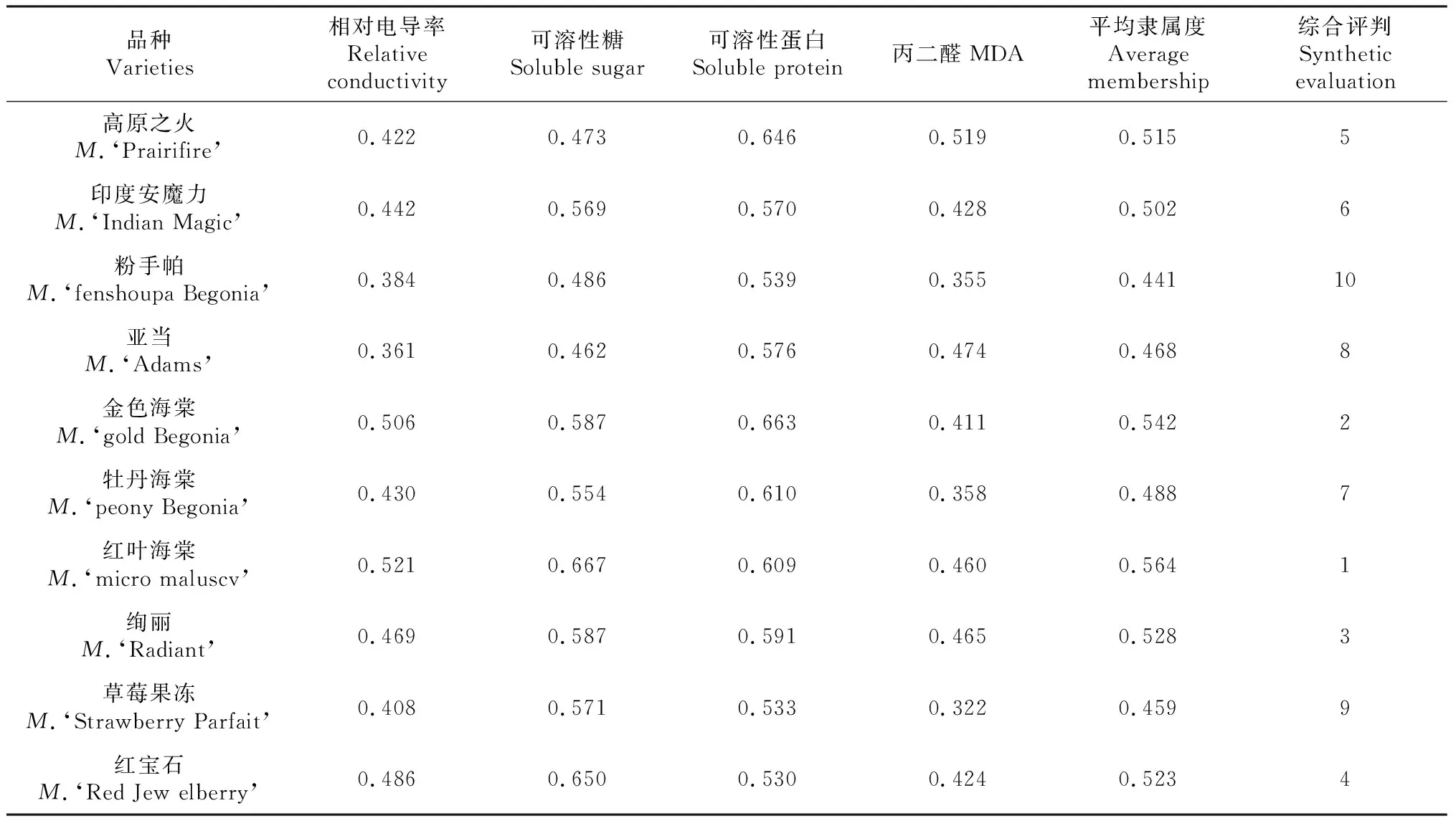
表5 不同北美海棠品种抗寒性的综合评判
本试验通过测定各低温处理后相对电导率、可溶性糖、可溶性蛋白、丙二醛含量等4个相关指标发现,在-20~-30 ℃范围内可溶性蛋白的变化,在-20~-35 ℃范围内可溶性糖和MDA的变化一定程度上体现了各品种抗寒能力。随着低温胁迫的加剧,相对电导率逐渐增大,在-30~-40 ℃范围内相对电导率的变化量较准确的反应北美海棠的抗寒性,根据相对电导率在-30~-40 ℃范围内的变化量,可知北美海棠各品种抗寒性的强弱为红叶海棠>金色海棠>牡丹海棠>红宝石>绚丽>粉手帕>印度安魔力>亚当>草莓果冻>高原之火。
果树是多年生木本植物,其受到的逆境胁迫而引起植物生理生化指标变化的因素是错综复杂的[20],因此,运用单一抗寒指标难于判断果树对寒冷的综合适应能力,也不能充分揭示抗寒的本质,为了全面准确的鉴定北美海棠的抗寒性,克服单一指标在北美海棠抗寒鉴定中的不足,本文通过研究低温处理后各项抗寒性生理指标的变化,在多个指标测定的基础上,运用隶属函数法进行综合评价。不同北美海棠品种抗寒能力的强弱排序为:红叶海棠>金色海棠>绚丽>红宝石>高原之火>印度安魔力>牡丹海棠>亚当>草莓果冻>粉手帕,该排序结果与相对电导率的排序结果基本一致。本研究结果为今后北美海棠的抗寒性选育和生产发展提供了理论依据。但树龄、采集时间的不同,北美海棠的各个生理指标可能发生变化,因此,进行抗寒评价时,需要建立新的综合评价方法进行评价。植物体抗寒机制非常复杂,影响其抗寒性的因素较多,因此今后还需要不断地深入研究北美海棠抗寒性机理,并同时结合田间观察,才能使结果更加准确、科学。
4 结 论
本研究发现,随着低温胁迫的加剧,不同北美海棠枝条的相对电导率均呈现上升趋势。可溶性糖、可溶性蛋白、丙二醛含量呈先上升后降低趋势。综合评价显示,不同北美海棠品种抗寒性的强弱排序为:红叶海棠>金色海棠>绚丽>红宝石>高原之火>印度安魔力>牡丹海棠>亚当>草莓果冻>粉手帕。
参考文献:
[1]刘志强,汤庚国. 海棠在园林中的应用研究[J].苏州科技学院学报(工程技术版),2004,17(3):75-80.
[2]李 鹏,王志忠,沈红香. 风景园林中的瑰宝一观赏海棠[J]. 中国花卉园艺,2006(12):44-46.
[3]林 娜,姜卫兵,翁忙玲. 海棠树种资源的园林特性及其开发利用[J]. 中国农学通报,2006,22(10):242-247.
[4]郑 杨,曲晓玲,郭 翎,等. 观赏海棠资源谱系分析及育种研究进展[J]. 山东农业大学学报(自然科学版), 2008,39(1):152-160.
[5]李 伟,杨 彬. 北美海棠系列在昌吉地区的引种栽培试验[J].新疆林业,2012(1):21-24.
[6]徐基平,李艳红,胡秀琴. 克拉玛依地区北美海棠的引种栽培及推广应用研究[J].园艺与种苗,2011(1):51-53.
[7]沈贤辉,刘 刚. 植物抗寒生理研究进展[J]. 长江大学学报(自科版),2014(17):40-42.
[8]沈德绪. 果树育种实验技术[M]. 北京:中国农业出版社,1992.
[9]邹 琦. 植物生理学实验指导[M]. 北京:中国农业出版社,2004.
[10]刘家尧. 植物生理学实验教程[M]. 北京:高等教育出版社,2010.
[11]潘瑞炽. 植物生理学[M]. 北京:高等教育出版社,2008.
[12]谢丽芬. 葡萄砧木抗寒性鉴定及抗寒机理的研究[D]. 内蒙古农业大学,2007.
[13]杨凤翔,金 芳,颜 霞.不同草莓品种抗寒性综合评价[J]. 果树学报,2010,27(3):368-372.
[14]李合生. 现代植物生理学[M]. 北京:高等教育出版社,2002.
[15]LYONS J M. Chilling injury in plants[J]. Ann Rev Plant Physical, 1975, 24: 445-446.
[16]孟繁静. 植物生理学[M]. 北京:华中理工大学出版社,2000.
[17]Mohapatra S S, Poole R J, Dhindsa R S. Cold acclimation freezing resistance and protein synthesis in alfalfa[J]. Exp. Bot,1987,38(10): 1697-1703.
[18]潘 杰,简令成,钱迎倩. 小麦抗寒力诱导过程中特异性蛋白质的合成[J]. 植物学集刊,1994(7):144-157.
[19]曾超西,王以柔,刘鸿先. 低温下黄瓜幼苗子叶硫氯基(SH)含量变化与膜脂过氧化[J]. 植物学报,1991,33(1):50-54.
[20]王丽华,李 西,刘 尉,等. 四种暖季型草坪草对SO2的抗性及净化能力的比较[J]. 草业学报,2013,22(1):225-233.
猜你喜欢
蔬菜(2022年10期)2022-11-10
小哥白尼(野生动物)(2022年3期)2022-06-16
热带作物学报(2022年5期)2022-06-01
今日农业(2021年20期)2021-11-26
河北果树(2020年4期)2020-11-26
山西农业科学(2020年10期)2020-10-16
河北果树(2020年1期)2020-02-09
浙江农业科学(2017年8期)2017-08-30
学苑创造·A版(2016年8期)2016-07-06
海峡姐妹(2015年7期)2015-02-27
