
四川小麦品种籽粒硬度和穗发芽抗性相关基因的分子标记鉴定
2018-06-05 05:56李式昭郑建敏万洪深杨漫宇罗江陶刘廷辉杨开俊蒲宗君
西南农业学报 2018年4期
李式昭,郑建敏,伍 玲,李 俊,万洪深,杨漫宇,罗江陶,刘廷辉,杨开俊,蒲宗君*
(1. 四川省农业科学院作物研究所/农业部西南地区小麦生物学与遗传育种重点实验室, 四川 成都 610066; 2. 四川省甘孜州农科所, 四川 康定 626000)
【研究意义】籽粒硬度是决定小麦品质的关键性状之一,对面粉颗粒大小、出粉率、润麦加水量、破损淀粉含量和最终食品加工品质都有重要影响。而穗发芽(Pre-harvest sprouting,PHS)则指小麦收获前遇到阴雨天气时籽粒在穗上发芽的现象,不仅降低产量,而且严重劣化品质。穗发芽后的小麦α-淀粉酶活性提高,导致淀粉水解,面粉的降落值降低,烘烤品质变差[1]。分析四川小麦籽粒硬度和穗发芽抗性相关基因分布,对四川小麦的品质改良具有重要意义。【前人研究进展】小麦籽粒硬度主要受位于5D染色体短臂上的1对主效基因Ha调控,由puroindoline蛋白控制,该蛋白的2种组分puroindoline a(PINA)和puroindoline b(PINB)是形成籽粒硬度的基础,分别由基因Pina和Pinb编码[2-3]。小麦籽粒硬度的差异主要源于Pina和Pinb基因的不同变异类型,当Pina和Pinb基因都处于野生(功能)状态时,籽粒表现为软质;但当其中任一基因缺失或发生突变时,籽粒表现为硬质,但也有例外[4]。软质麦适宜制作饼干和糕点等甜食,而硬质麦则更适合制作面包和面条等食品。不同puroindoline等位变异类型对磨粉和加工品质的影响也不尽相同,目前已命名17个Pina位点和25个Pinb位点的等位变异[5]。相较于Pina位点,对Pinb-D1b基因的研究则更为深入,该基因的氨基酸序列中第46位点的甘氨酸(Glycine,Gly)突变为丝氨酸(Serine,Ser),造成小麦质地变硬。相关研究[6-8]表明,Pinb-D1b较Pina-D1b出粉率高、灰份低,磨粉品质更好,此外,前者的馒头、面包和面条加工品质也略优于PINA蛋白缺失和野生类型。小麦穗发芽则与种子的休眠期长短显著相关,休眠期越长,穗发芽抗性越强;休眠期短或无休眠期的品种更易发生穗发芽[9]。Vp-1基因是种子成熟、干燥及休眠的主要转录调节因子,能促进胚成熟,加速休眠并抑制其萌发[10],该基因位于小麦第3同源群染色体的长臂上[11]。研究[12-14]表明,等位基因类型Vp-1Bb和Vp-1Bc与抗穗发芽相关,而Vp-1Ba则与感穗发芽相关。【本研究切入点】目前,针对上述品质性状,国内、外已开发出多种鉴定上述基因内部等位变异的功能标记[15-16],方便进行快速检测。【拟解决的关键问题】本研究利用2个籽粒硬度相关分子标记Pinb-D1a、Pinb-D1b和1个穗发芽抗性等位基因分子标记Vp1B3,对2000年以来四川育成的105份小麦品种进行鉴定,旨在深入了解四川小麦品种籽粒硬度和穗发芽抗性相关等位基因的变异类型和分布规律,为选配有价值的品质育种亲本,加速四川小麦品质育种进程提供理论依据。
1 材料与方法
1.1 材料
试验选用自2000年以来四川育成的105份优良小麦新品种,其中包括:川麦系列32份、绵麦系列(含绵阳、绵麦、国豪麦)19份、川农系列(含川农、良麦、蜀麦)18份、川育系列(含川育、科成麦)10份、西科麦系列(含绵农、西科麦)8份和其他单位品种18份。2个籽粒硬度和1个穗发芽抗性等位基因分子标记的引物序列信息见表1。
1.2 基因组DNA提取
每份品种采用5株幼苗混合取样,参照Murray等[17]的微量CTAB法提取小麦基因组DNA,用于基因位点检测,并确定105份品种的基因型。
1.3 PCR扩增及电泳检测
全部3对引物均由生工(上海)生物工程股份有限公司(http://www.sangon.com)合成。PCR反应在AB Veriti型PCR扩增仪中进行,各分子标记的PCR反应体系、程序及扩增产物的检测均采用表1参考文献方法进行。
2 结果与分析
2.1 籽粒硬度相关基因的分子标记检测
在Pinb-D1位点,105份四川小麦品种中,有95份品种扩增出标记Pinb-D1a显示非Pinb-D1b等位基因类型的250 bp特征带(图1A),频率为89.5 %;11份品种扩增出标记Pinb-D1b显示Pinb-D1b等位基因类型(Pinb突变)的250 bp特征带(图1B),频率为10.5 %。
2.2 穗发芽抗性相关基因的分子标记检测
在Vp-1B位点,105份四川小麦品种中,有88份品种扩增出共显性标记Vp1B3显示Vp-1Bc(抗穗发芽)的569 bp特征带,频率为83.8 %;17份品种扩增出显示Vp-1Ba(感穗发芽)的652 bp特征带,频率为16.2 %(图2);未能扩增出含有Vp-1Bb(抗穗发芽)等位基因类型的品种。
表1 分子标记PCR引物及其相关信息
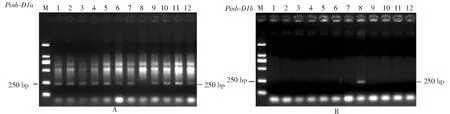
M:DL2000;1:川麦39;2:川麦44;3:川麦50;4:川麦56;5:川麦65;6:川麦107;7:川农23;8:绵阳35;9:绵麦46;10:川育20;11:西科麦4号;12:杏麦2号M: DL2000; 1: Chuanmai 39; 2: Chuanmai 44; 3: Chuanmai 50; 4: Chuanmai 56; 5: Chuanmai 65; 6: Chuanmai 107; 7: Chuannong 23; 8: Mianyang 35; 9: Mianmai 46; 10: Chuanyu 20; 11: Xikemai 4; 12: Xingmai 2图1 特异性籽粒硬度标记Pinb-D1a(A)、Pinb-D1b(B)对部分供试品种的PCR扩增结果Fig.1 PCR products amplified by specific grain hardness markers Pinb-D1a (A) and Pinb-D1b (B) in several tested cultivars
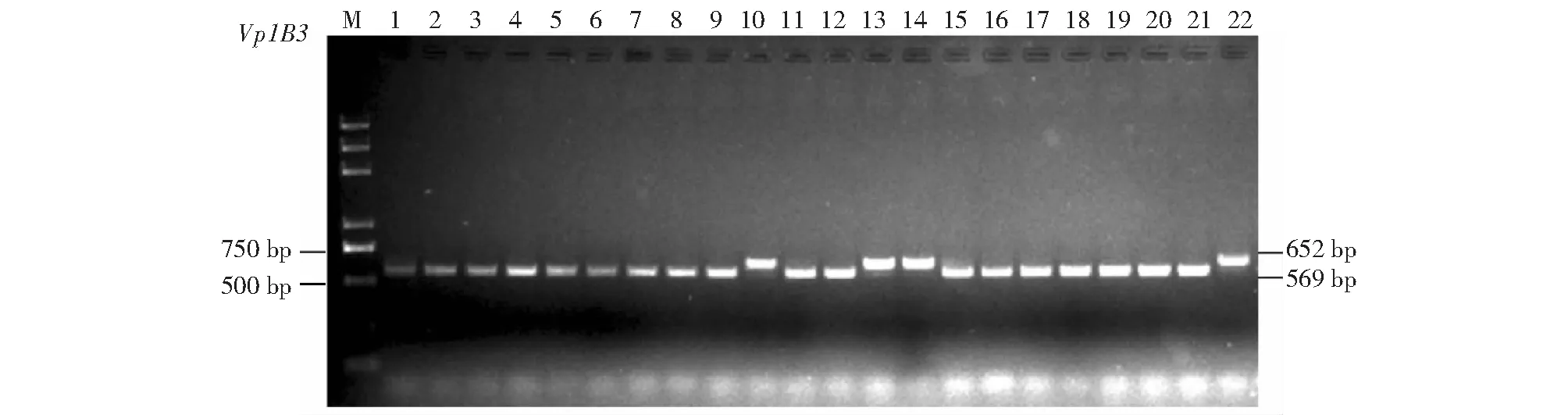
M:Trans2K plus DNA marker;1:川麦32;2:川麦37;3:川麦39;4:川麦42;5:川麦44;6:川麦48;7:川麦51;8:川麦55;9:川麦58;10:川麦62;11:川麦64;12:川麦104;13:川麦65;14:川麦107;15:川农23;16:川农27;17:绵阳30;18:绵麦37;19:绵麦51;20:川育20;21:西科麦4号;22:蜀麦482M: Trans2K plus DNA marker; 1: Chuanmai 32; 2: Chuanmai 37; 3: Chuanmai 39; 4: Chuanmai 42; 5: Chuanmai 44; 6: Chuanmai 48; 7: Chuanmai 51; 8: Chuanmai 55; 9: Chuanmai 58; 10: Chuanmai 62; 11: Chuanmai 64; 12: Chuanmai 104; 13: Chuanmai 65; 14: Chuanmai 107; 15: Chuannong 23; 16: Chuannong 27; 17: Mianyang 30; 18: Mianmai 37; 19: Mianmai 51; 20: Chuanyu 20; 21: Xikemai 4; 22: Shumai 482 图2 特异性穗发芽抗性标记Vp1B3对部分供试材料的PCR扩增结果Fig.2 PCR products amplified by specific resistance to PHS markers Vp1B3 in several tested cultivars
2.3 四川小麦籽粒硬度和穗发芽抗性基因的分布
105份四川小麦品种籽粒硬度和穗发芽抗性相关基因的分布如表2所示,聚合2项优质基因(同时含有Pinb-D1b和Vp-1Bc)的四川小麦品种有11份,分别为川麦55、川农12、川农18、绵阳35、绵麦43、国豪麦15、蓉麦2号、良麦4号、蜀麦375、渝麦7号和南麦302,频率占全部品种的10.5 %。
3 讨 论
3.1 优质籽粒硬度基因在四川小麦中的分布与组成形式
Giroux等[15]研究表明,硬质小麦Cheyenne的质地变化是由于其Pinb基因的第46位氨基酸由甘氨酸(G)突变为丝氨酸(S)造成的。本研究显示,四川小麦品种含Pinb-D1b(Pinb突变)的频率较低,仅占10.5 %。其中,32份川麦、19份绵麦、18份川农、10份川育、8份西科麦系列品种中含Pinb-D1b的材料分别仅有1、3、4、0、0份,频率为3.1 %、15.8 %、22.2 %、0.0 %和0.0 %。陈锋等[7]研究认为,中国小麦品种Pinb-D1b变异类型较多,占54.5 %,但西南区频率较低,仅占19.2 %。不同地区的Pinb-D1b分布也不尽相同,陕西占57.4 %[18],青海占15.2 %[19]。本研究中所采用的105份小麦品种基本涵盖了2000年以来四川省的育成品种,能够代表该省小麦的遗传特征,检测表明四川小麦品种Pinb-D1b频率较低,与陈锋等[7]结论一致,这不仅与自然地理环境和当地的饮食习惯有关,也与选育品种所选用的亲本资源相关。在实际育种过程中,籽粒硬度已可通过硬度仪(单籽粒谷物特性测试仪,Single Kernel Characterization System,SKCS)等仪器快速进行表型选择,今后四川小麦的籽粒硬度品质育种,可多考虑引入含Pinb-D1b类型的材料,改善Puroindoline基因的遗传多样性,适度提升硬质麦的比例。

表2 105份四川小麦籽粒硬度和穗发芽抗性的分子标记检测结果
注:下划线表示正向品质效应基因。
Note: The gene names underlined mean genes with positive effect to noodle quality.
3.2 优质穗发芽抗性基因在四川小麦中的分布与组成形式
四川因小麦收获期与雨季相接,近年来已成为小麦穗发芽的重灾区,从本研究结果来看,抗穗发芽基因资源已经在四川得到良好应用,含Vp-1Bc(抗穗发芽)的品种频率占83.8 %,高于中国推广小麦品种抗穗发芽的平均频率(71.7 %)[14],接近优质面条商品小麦澳白麦的水平(88.9 %)[20]。川麦、绵麦、川农、川育、西科麦系列品种含Vp-1Bc的材料分别有23、17、17、10、7份,频率为71.9 %、89.5 %、94.4 %、100.0 %和87.5 %。究其原因,可能是四川阴雨潮湿气候较多,生产上种植的小麦品种在较大的选择压力下得以保留丰富的抗穗发芽基因资源,但也应看到,四川还缺少含Vp-1Bb(高抗穗发芽)基因资源的材料;同时,现有的抗穗发芽品种若遭遇连续遇雨的自然环境,其抗性表现也会出现很大差异,部分含Vp-1Bc(抗穗发芽)的品种依然大量出现穗发芽现象,这就要求育种家尽早开拓出新的穗发芽抗源。另一方面,红粒[12]、白粒[13]小麦的抗穗发芽能力也不尽相同,一般来说,红粒小麦较白粒休眠期更长,更抗穗发芽,但白粒小麦更受农民亲睐,充分挖掘现有的白粒抗穗发芽小麦材料就显得更为重要。
3.3 聚合优质籽粒硬度和穗发芽抗性基因品种在四川小麦中的分布
聚合优质籽粒硬度和穗发芽抗性基因的四川小麦品种有川麦55等11份,频率为10.5 %;但其中白粒品种仅有4份,分别为川麦55、绵阳35、国豪麦15和渝麦7号,仅占全部品种的3.8 %。上述品种可成为改良籽粒硬度和穗发芽抗性的宝贵资源材料,直接利用它们作为优质基因导入的亲本,可能是改良四川小麦品质的捷径。同时,也应看到,部分品种的实际磨粉品质与基因检测结果并不十分吻合,如川麦36和川麦39,是四川省审定的为数不多的强筋小麦品种,硬度值较高,但检测结果是非Pinb-D1b类型的;在穗发芽抗性上,含抗穗发芽Vp-1Bc基因的绵麦367在实际生产中反而较感穗发芽。这充分说明籽粒硬度和穗发芽抗性均是由多基因位点控制的,实际品质表现是多种基因共同作用的结果,继续发掘其他相关基因位点的功能,才能更好地为分子标记辅助育种工作服务。
4 结 论
本研究检测的105份四川小麦品种的籽粒硬度和穗发芽抗性基因组成具有丰富的多态性,在四川小麦品质育种中逐步导入优质基因Pinb-D1b和Vp-1Bc,是综合提高四川小麦品质的有效途径。
参考文献:
[1]Humphreys D G, Noll J. Methods for characterization of preharvest sprouting resistance in a wheat breeding program[J]. Euphytica, 2002, 126:61-65.
[2]Morris C F. Puroindolines: the molecular genetic basis of wheat grain hardness[J]. Plant Molecular Biology, 2002, 48:633-647.
[3]Chen F, Beecher B, Morris C F. Physical mapping and a new variant ofPuroindolineb-2 genes in wheat[J]. Theoretical and Applied Genetics, 2010, 120:745-751.
[4]夏兰芹,何中虎,陈新民,等. 小麦硬度主效基因Pina和Pinb的克隆和序列分析[J]. 作物学报, 2003, 29(1):25-30.
[5]Morris C F, Bhave M. Reconciliation of D-genome puroindoline allele designations with current DNA sequence data[J]. Journal of Cereal Science, 2008, 48:277-287.
[6]Martin J M, Frohberg R C, Morris C F, et al. Milling and bread baking traits associated with puroindoline sequence type in hard red spring wheat[J]. Crop Science, 2001, 41:228-234.
[7]陈 锋,钱森和,张 艳,等. 中国冬小麦puroindoline类型分布及其对溶剂保持力的影响[J]. 中国农业科学, 2005, 38(11):2173-2181.
[8]陈 锋,陈东升,钱森和,等. Puroindoline基因对春小麦磨粉及馒头、面条品质的影响[J]. 作物学报, 2006, 32(7):980-986.
[9]Li C D, Ni P X, Francki M, et al. Genes controlling seed dormancy and pre-harvest sprouting in a rice-wheat-barley comparison[J]. Functional & Integrative Genomics, 2004(4):84-93.
[10]McCarty D R, Hattori T, Carson C B, et al. The Viviparous-1 developmental gene of maize encodes a novel transcriptional activator[J]. Cell, 1991, 66:895-905.
[11]Bailey P C, McKibbin R S, Lenton J R, et al. Genetic map location for orthologousVP1 genes in wheat and rice[J]. Theoretical and Applied Genetics, 1999, 98:281-284.
[12]杨 燕,张春利,陈新民,等. 红粒春小麦穗发芽抗性鉴定及相关分子标记的有效性验证[J]. 麦类作物学报, 2011, 31(1):54-59.
[13]苗西磊,王德森,夏兰芹,等. 白粒小麦品种(系)穗发芽抗性机制分析[J]. 麦类作物学报, 2011, 31(4):741-746.
[14]杨 燕,王晓丽,刘世鑫,等. 利用STS标记检测我国小麦推广品种的抗穗发芽基因型[J]. 华北农学报, 2013, 28(3):183-188.
[15]Giroux M J, Morris C F. A glycine to serine change in puroindoline b is associated with wheat grain hardness and low levels of starch-surface friabilin[J]. Theoretical and Applied Genetics, 1997, 95:857- 864.
[16]Yang Y, Zhao X L, Xia L Q, et al. Development and validation of aViviparous-1 STS marker for pre-harvest sprouting tolerance in Chinese wheats[J]. Theoretical and Applied Genetics, 2007, 115:971-980.
[17]Murray H G, Thompson W F. Rapid isolation of high molecular weight DNA[J]. Nucleic Acids Research, 1980(8):4321-4325.
[18]张 晶,张晓科,王可珍,等. 陕西小麦籽粒硬度及其基因型分析[J]. 麦类作物学报, 2011, 31(4):666-671.
[19]王雪玲,李建民,魏 乐,等. 青海小麦籽粒硬度等位变异研究[J]. 麦类作物学报, 2014, 34(1):23-27.
[20]李式昭,伍 玲,郑建敏,等. 优质面条商品小麦澳白麦相关品质基因的分子标记鉴定[J]. 中国农业科学, 2012, 45(18):3677-3687.
猜你喜欢
当代水产(2022年7期)2022-09-20
河北科技师范学院学报(2022年2期)2022-08-26
中国特种设备安全(2022年1期)2022-04-26
山东冶金(2022年1期)2022-04-19
现代畜牧科技(2021年4期)2021-12-05
中国饲料(2021年17期)2021-11-02
空间科学学报(2021年1期)2021-05-22
化工管理(2021年7期)2021-05-13
作物研究(2020年5期)2020-12-08
种业导刊(2017年7期)2017-08-22
