
Shh基因在NIPBL+/-胎鼠肢芽内的表达*
2018-05-29 08:27潘金勇嵇继宇张惠荣石河子大学医学院第一附属医院儿科新疆石河子832000
现代医药卫生 2018年10期
王 莉,潘金勇,嵇继宇,张惠荣(石河子大学医学院第一附属医院儿科,新疆石河子832000)
Cornelia de Lange综合征(CdLS)是一种罕见的先天性疾病,患病率为1/100 000~1/10 000[1]。临床表现主要为严重的生长发育迟缓、智力障碍、畸形等[2]。CdLS的患者约60%是NIPBL基因发生了病理性变化[3]。有研究表明,NIPBL基因水平的高低与CdLS的严重程度密切相关[4]。Shh基因是由ZRS增强子调控参与肢芽的发育[5],Shh基因在由ZRS增强子调控的胚胎极化活性区(ZPA)进行肢芽的表达[6]。然而,在小鼠胚胎发育阶段的肢芽中,NIPBL基因对位于ZPA区域的Shh基因具有调控作用,会影响Shh基因的表达。但2个基因之间
的具体交互作用及规律鲜见文献报道。本研究采用NIPBL-Loxp小鼠与Cre小鼠杂交,建造NIPBL+/-小鼠模型,探讨NIPBL对Shh基因表达的影响,为CdLS的诊断和干预提供新的策略。
1 材料与方法
1.1 材料
1.1.1 动物 NIPBL-Loxp小鼠与Cre小鼠的背景品系小鼠是C57BL/6J,SPF级,购于浙江大学,全部小鼠饲养于石河子大学药学院实验动物中心。实验过程遵照石河子大学《动物实验伦理委员会管理条例》。挑选NIPBL+/-小鼠雄性体重35 g左右,雌性体重30 g左右,晚上20:00将雌雄小鼠按2∶1合笼,次日早晨8:00检查雌鼠的阴栓,阴栓阳性者将次日12:00作为其胚胎E0.5天。
1.1.2 仪器与试剂 逆转录试剂盒Prime Script RT reagent Kit购于Thermo公司;总RNA提取剂Trizol Reagent购于Life公司;逆转录试剂盒RevertAid First Strand cDNA Synthesis Kit购于Thermo公司;荧光定量试剂盒SYBR Green PCR Kit购于Thermo公司;普通聚合酶链反应(PCR)仪器购于TaKaRa公司;荧光定量聚合酶链反应(qRT-PCR)仪器购于Life公司。
1.2 方法
1.2.1 动物模型的建立和实验分组 采用NIPBL-Loxp小鼠与Cre小鼠建造NIPBL+/-小鼠模型,NIPBL+/-小鼠与NIPBL+/-小鼠按照雌、雄2∶1合笼进行杂交,分别在孕鼠的E10、E11、E12天取出孕鼠放在超净台内,以颈部脱臼法处死孕鼠,在严格无菌操作条件下,分别在孕鼠的E10、E11、E12天用逆转录聚合酶链反应(RT-PCR)逆转录试剂盒鉴定得到6只NIPBL+/-胎鼠的肢芽作为实验组,另选取6只NIPBL+/+胎鼠的肢芽作为对照组,引物设计见表1;最后分离胎鼠双侧四肢,放入液氮15~20 min,再放入-80℃冰箱,以备后期实验。

表1 NIPBL-Loxp小鼠与Cre小鼠引物信息
1.2.2 采用qRT-PCR检测Shh基因的表达情况 Shh基因引物的设计见表2。

表2 Shh基因和β-actin的引物信息
1.2.3 实验组和对照组RNA的提取 按照RNA提取试剂盒说明书提取总RNA。
1.2.4 实验组和对照组cDNA的合成 按照逆转录试剂盒说明书对上述提取的总RNA进行逆转录。
1.2.5 qRT-PCR检测实验组和对照组Shh基因的表达情况 以上述cDNA为模板,按照荧光定量试剂盒说明书检测实验组和对照组Shh基因的表达量。
1.3 统计学处理 采用SPSS17.0统计软件进行数据处理。符合正态分布的计量资料以±s表示,对数据进行方差齐性检验,方差不齐计量资料的多组间比较采用非参数检验,组间比较采用t检验。P<0.05为差异有统计学意义。
2 结 果
2.1 NIPBL基因敲除小鼠模型的验证 NIPBL基因敲除打靶载体构建策略图,见图1。

图1 野生型及突变型等位基因示意图
2.2 NIPBL-Loxp和Cre小鼠的引物鉴定结果 NIPBLLoxp和Cre小鼠建造的NIPBL+/-小鼠模型亲代鼠,经RT-PCR技术鉴定出在孕鼠E10、E11、E12天的NIPBL+/-和NIPBL+/+胎鼠,经鉴定出来的引物凝胶电泳结果见图 2~4。

图2 NIPBL-Loxp-F/NIPBL-Loxp-R凝胶电泳结果
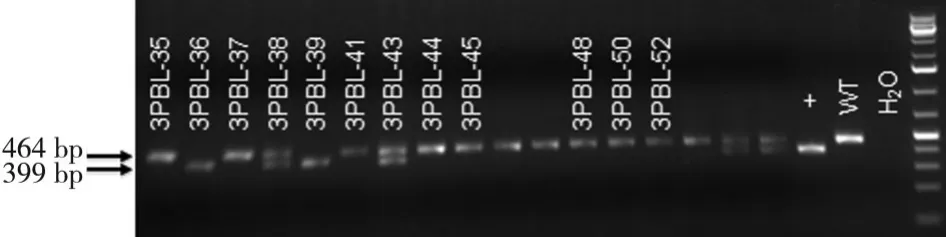
图3 Cre-WT-F/Cre-WT-R凝胶电泳结果
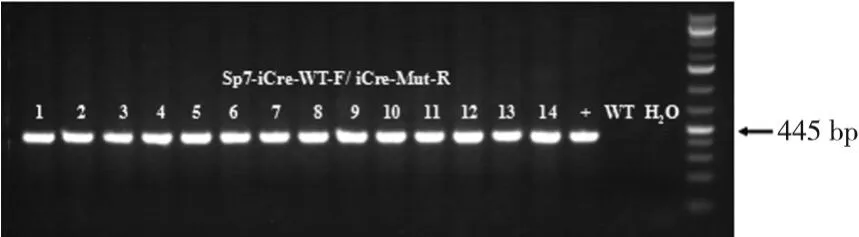
图4 Cre-WT-F/Cre-Mut-R凝胶电泳结果
2.3 qRT-PCR验证结果 实验组和对照组胎鼠肢芽内的Shh基因在E10、11、12天均有表达,实验组和对照组胎鼠肢芽内Shh基因的ΔCt值由大到小依次为E10、E12、E11 天,表明在不同的孕期(E10、E11、E12天)时,实验组和对照组胎鼠肢芽内Shh基因表达趋势均为先升后降,即表示实验组与对照组胎鼠肢芽的Shh基因表达量在E10天时有表达,E11天时达顶峰,E12天时表达量下降,但仍高于E10天表达量。实验组在E10、E11、E12天Shh基因的表达量比较,差异有统计学意义(P<0.01)。与对照组比较,在相同的孕期(E10、E11、E12天)内,实验组Shh基因的ΔCt值大于对照组,表明实验组胎鼠肢芽内Shh基因的表达水平低于对照组,差异有统计学意义(P<0.01),见表3。
表3 实验组与对照组Shh基因ΔCt值的比较(±s)
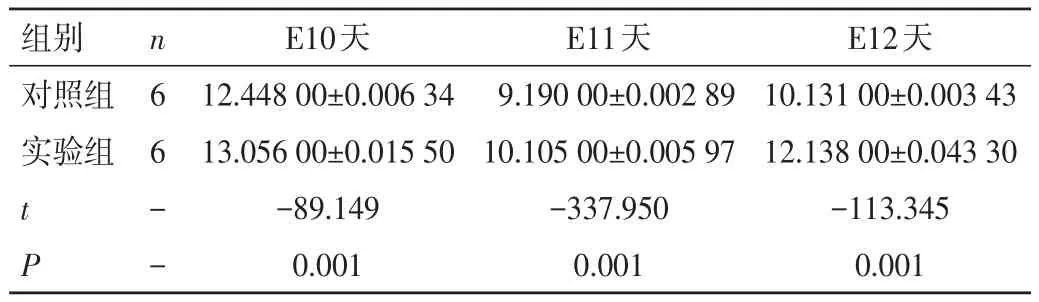
表3 实验组与对照组Shh基因ΔCt值的比较(±s)
注:-表示无此项
组别对照组实验组t P n6 6- -E10天12.448 00±0.006 34 13.056 00±0.015 50-89.149 0.001 E11天9.190 00±0.002 89 10.105 00±0.005 97-337.950 0.001 E12天10.131 00±0.003 43 12.138 00±0.043 30-113.345 0.001
3 讨 论
CdLS是一种多系统发育障碍性的常染色体显性遗传疾病[7]。生长发育迟缓、面部畸形和智力障碍等是CdLS的表型特征,其中生长发育迟缓是CdLS的重要表现[8]。有关研究报道,CdLS受多种基因及信号通路的调控,一旦这些基因及信号通路发生改变,都可能导致CdLS的发生。其中,NIPBL基因缺陷是导致CdLS最常见的原因,研究人员利用分子遗传工具创建了NIPBL基因敲除小鼠,试图了解CdLS的病因学[9]。有关研究报道,Hedgehog信号通路抑制剂限制了骨骼的发育成熟,其包括 Sonic hedgehog(Shh)、Desert hedgehog(Dhh)及Indian hedgehog(Ihh)3 个配体,其中最常见的是 Shh[10]。Shh基因编码一种信号蛋白,在小鼠胚胎肢芽的发育过程中起不可或缺的作用[11]。Shh基因编码的Shh蛋白是与ZPA极化功能有关的一种形态发生素,由ZPA中细胞产生,ZPA区域传递肢芽早期的发育信号,启动Shh基因的表达[12]。有研究表明,小鼠胚胎肢芽中Shh蛋白的变化存在于肢体发育前10 h,且Shh蛋白量的改变会影响肢体缺陷的程度[13]。相关研究显示,NIPBL在ZPA区域和肢芽的间充质细胞对Shh基因进行调控,一旦NIPBL基因缺陷,会引起Shh基因表达的信号通路受到抑制,造成Shh基因的表达缺失,引起ZPA区域染色体位点异常,从而形成了CdLS的特征性改变[14]。因此,本研究通过建立NIPBL基因敲除模型,进一步研究NIPBL和Shh2个基因之间的相互作用机制。
有研究报道,Shh基因通常在正常胎鼠肢芽的E9.5~E11.5天进行表达,且在孕鼠的E10天左右表达量逐渐增多,在E11天大量表达,在E12天左右表达量逐渐减少[15]。本实验观察到胎鼠在E9.5天时仍是个椭球形,很难分辨出胚胎的器官,Shh基因也几乎检测不出表达;E12.5天时,胎鼠的肢芽也几乎检测不到Shh基因的表达。因此,本实验选取胎鼠的E10、E11、E12天作为实验分组。本实验结果表明,在孕鼠的E10、E11、E12天,实验组和对照组的胎鼠肢芽内均有Shh基因的表达,但是实验组胎鼠肢芽内Shh基因的ΔCt值大于对照组,表明实验组胎鼠肢芽内Shh基因的表达水平低于对照组(ΔCt值越高,Shh基因的表达水平越低),说明NIPBL基因缺陷影响胎鼠肢芽内Shh基因表达的正常信号通路,抑制了Shh基因的表达。实验组和对照组胎鼠肢芽内Shh基因ΔCt值大小依次为E10、E12、E11天,表明在不同的孕期(E10、E11、E12天)内,实验组和对照组胎鼠肢芽内Shh基因的表达趋势是E10天时有表达,但表达量较少,随后表达量逐渐增多,在E11天时达到顶峰,随后表达量逐渐减少,E12天表达量较E11天减少,但比E10天表达量增多,差异有统计学意义(P<0.01)。
综上所述,NIPBL+/-基因敲除抑制胎鼠肢芽内Shh基因的表达,而Shh基因在体内正常的软骨发育中起重要调控作用,由此推断NIPBL+/-基因敲除胎鼠可能会抑制软骨的发育成熟,进一步导致CdLS。本研究有助于人们进一步了解NIPBL基因在CdLS发病中的机制和意义,并为CdLS疾病的诊断和干预提供新的策略,帮助研究人员深入了解CdLS疾病,以便探索更好的治疗方法。
[1]MASKOEN AM,LAKSONO B,HAJJAH R,et al.Cornelia de lange syndrome with thyroid agenesis of an indonesian patient[J].Cell Mol Biol,2017,63(8):93-94.
[2]INFANTE E,ALKORTA-ARANBURU G,EL-GHARBAWY A,et al.Rare form of autosomal dominant familial Cornelia de Lange syndrome due to a novel duplication in SMC3[J].Clin Case Rep,2017,5(8):1277-1283.
[3]PUISAC B,TERESA-RODRIGO M,BAQUERO-MONTOYA C,et al.mRNA quantification of NIPBL isoforms a and B in adult and fet al human tissues,and a potentially pathological variant affecting only isoform a in two patients with cornelia de lange syndrome[J].Int J Mol Sci,2017,18(3):481.
[4]KAUR M,MEHTA D,NOON SE,et al.NIPBL expression levels in CdLS probands as a predictor of mutation type and phenotypic severity[J].Am J Med Genet C Semin Med Genet,2016,172(2):163-170.
[5]LETTICE LA,DEVENNEY P,DE ANGELIS C,et al.The conserved sonic hedgehog limb enhancer consists of discrete functional elements that regulate precise spatial expression[J].Cell Rep,2017,20(6):1396-1408.
[6]PELUSO S,DOUGLAS A,HILL A,et al.Fibroblast growth factors(FGFs)prime the limb specific shh enhancer for chromatin changes that balance histone acetylation mediated by E26 transformation-specific(ETS)factors[J].Elife,2017,6(1):28590.
[7]NEWKIRK DA,CHEN YY,CHIEN R,et al.The effect of Nipped-B-like(Nipbl)haploinsufficiency on genome-wide cohesin binding and target gene expression:modeling Cornelia de Lange syndrome[J].Clin Epigenetics,2017,9(1):89.
[8]PARENTI I,TERESA-RODRIGO ME,POZOJEVIC JA,et al.Mutations in chromatin regulators functionally Link Cornelia de Lange syndrome and clinically overlapping phenotypes[J].Hum Genet,2017,136(3):307-320.
[9]KAWAUCHI S,SANTOS R,MUTO AA,et al.Using mouse and zebrafish models to understand the etiology of developmental defects in cornelia de lange syndrome[J].Am J Med Genet C Semin Med Genet,2016,172(2):138-145.
[10]ROBINSON GW,KASTE SC,CHEMAITILLY W,et al.Irreversible growth plate fusions in children with medulloblastoma treated with a targeted hedgehog pathway inhibitor[J].Oncotarget,2017,8(41):69295-69302.
[11]AMANO T,SAGAI T,SEKI R,et al.Two types of etiological mutation in the Limb-Specific enhancer of shh[J].G3,2017,7(9):2991-2998.
[12]MATSUBARA H,SAITO D,ABE G,et al.Upstream regulation for initiation of restricted Shh expression in the chick limb bud[J].Dev Dynam,2017,246(5):417-430.
[13]ZHANG R,LEE C,LAWSON LY,et al.SHH protein variance in the limb bud is constrained by feedback regulation and correlates with altered digit patterning[J].G3,2017,7(3):851-858.
[14]MUTO A,IKEDA S,LOPEZ-BURKS ME,et al.Nipbl and mediator cooperatively regulate gene expression to control limb development[J].PLoS Genet,2014,10(9):1004671.
[15]YOKOYAMA S,FURUKAWA S,KITADA S,et al.Analysis of transcription factors expressed at the anterior mouse limb bud[J].PLoS One,2017,12(5):0175673.
猜你喜欢
波谱学杂志(2021年1期)2021-03-12
中国实验诊断学(2020年12期)2020-12-25
科学(2020年3期)2020-11-26
中国药理学通报(2017年6期)2017-06-22
武昌理工学院学报(2017年1期)2017-05-11
中国实验动物学报(2016年2期)2016-07-15
中国实验诊断学(2016年1期)2016-02-29
中国康复理论与实践(2015年10期)2015-12-24
考试周刊(2015年91期)2015-09-10
中国病理生理杂志(2015年10期)2015-01-26
