
糯玉米单倍体自然加倍特性的研究
2018-05-21 07:56,,2,,,,,,,
种子 2018年4期
, ,2, , , , , , ,
(1.吉林农业大学农学院, 吉林 长春 130118; 2.吉林农业科技学院, 吉林 吉林 132101;3.吉林省科学技术信息研究所, 吉林 长春 130033)
糯玉米(ZeamaysL.certainaKulesh)亦称粘(蜡质)玉米,是在栽培种植玉米过程中发现的一种新类型[1-2]。糯玉米具有绵软适口、甜粘清香的独特口感和风味,既可以蒸煮用于青食,也可用于速冻加工、制作糯玉米罐头等,深受广大玉米食用爱好者的追捧[3]。另外,糯玉米中支链淀粉粘滞性强、透明度高,可作为纺织、造纸等轻工业的重要原料[4]。因此,糯玉米产业在我国发展潜力巨大。我国不同生态地区糯玉米种质资源极其丰富,但对糯玉米育种的研究起步较晚,育种家对大量的种质资源缺乏有效的育种技术手段,从而造成糯玉米育种田间工作量大、育种效率较低[5]。
现如今,基于生物诱导发展起来的DH育种(Doubled Haploid Breeding)技术已经广泛地应用于玉米育种中,该技术与分子标记辅助育种技术、转基因技术构成了现代玉米育种新体系[6]。玉米DH育种技术也正逐渐应用于糯玉米育种中,大大提高了糯玉米杂交组合的配制效率,缩短了优良糯玉米杂交种的选育年限[7]。然而,糯玉米单倍体加倍技术是目前糯玉米DH育种中的主要技术“瓶颈”[7]。迄今为止,糯玉米单倍体加倍很大程度上无法摆脱对化学加倍药剂的依赖,究其原因是缺乏对糯玉米单倍体在自然环境下加倍特性的相关研究,尤其是结合不同遗传背景材料对糯玉米单倍体雌雄育性恢复特征的系统研究等。基于此,本团队利用高频孤雌生殖诱导系所创造的糯玉米单倍体为供试材料,对糯玉米单倍体雌雄育性的自然恢复现象进行了相关探究,目的在于探索糯玉米单倍体自然加倍的实用技术,为进一步加快糯玉米DH育种进程提供理论依据。
1 材料与方法
1.1 材 料
供试糯玉米单倍体源自生产上推广面积较大的糯玉米杂交种(吉糯13、中糯1号、苏玉糯1号、万糯2000)和糯玉米骨干自交系(98 W-2、W 6、BN 2、D 6644、JN 15、景白浚34、JN-1)经诱导系DRL杂交诱导各母本材料产生,糯玉米单倍体诱导及鉴定方法参照Chase[8]提出的遗传标记法进行。对收获的杂交诱导果穗的种子逐粒鉴定,即:顶端胚乳糊粉层呈现紫色标记、胚芽部位无色且胚面较小,呈近似三角形的籽粒为糯玉米单倍体(图1 A、B)。单倍体田间表现为植株生长发育慢、较矮,叶片短且直立,茎秆绿色(图1 C)。
1.2 方 法
将2015年夏在吉林农业大学特用玉米育种基地经高频孤雌生殖诱导系DRL诱导的糯玉米单倍体籽粒分别于2015年冬、2016年按照不同播期种植在海南和北方玉米科研试验地,具体见表1。糯玉米单倍体雌雄穗的自然育性考察标准如下:
表2 吉糯13单倍体雌穗育性的自然恢复

单倍体来源单倍体株数(株)无雌穗株数(株)无雌穗率(%)授粉株数(株)结实株数(株)结实率(%)平均结实数/穗(粒)长春Ⅰ50714.00221359.0917(3-93)长春Ⅱ1091110.09382463.1629(4-67)长春Ⅲ1962010.201016665.3532(4-80)三亚178147.871136961.0630(1-114)平均10.5462.1727

图1 糯玉米单倍体诱导果穗、籽粒及其田间表现
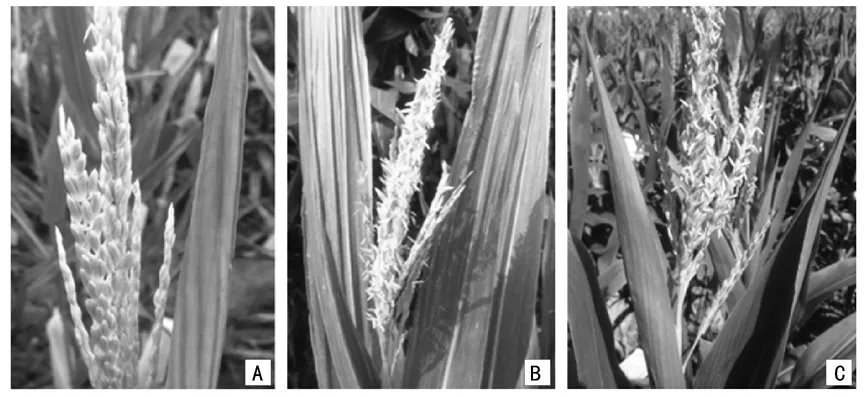
注:A为雄穗不育;B为仅在雄穗的主轴能散粉的单倍体;C为雄穗的主轴和侧枝均能够散粉的单倍体。图2 吉糯13单倍体群体雄穗田间散粉情况
1) 雄穗育性:仔细观察抽雄后的糯玉米单倍体雄穗,将能够散粉的植株记为雄性可育株,否则记为雄性不育株。
雄穗育性程度以散粉株率表示:散粉株率(%)=散粉植株数/观察植株总数×100%。
2) 雌穗育性:观察和记录自然条件下生长的糯玉米单倍体有无雌穗,用杂交种花粉给雄性不育株的雌穗授粉,授粉后能够结实的记为雌性可育株,否则记为雌性不育株。
雌穗育性程度以结实株率、无雌穗率表示:
结实株率(%)=结实植株数/授粉植株总数×100%;
无雌穗率(%)=无雌穗植株数/观察植株总数×100%。
1.3 数据处理及分析
综合利用Excel 2013软件和DPS 7.05软件进行数据统计分析[9]。
2 结果与分析
2.1 糯玉米单倍体雌穗育性的自然恢复
由表2可知,在4种不同生长环境下,不同吉糯13单倍体单株经DRL诱导获得的单倍体无雌穗率分别为14.00%、10.09%、10.20%和7.87%;糯玉米单倍体雌穗经过授粉后平均结实率为62.17%,由此可知,糯玉米单倍体雌穗多数都是可育的。
收获的糯玉米单倍体果穗均较小,但各穗所结实的粒数之间差异很大,多则上百粒,高达114粒,最少的仅结1粒,各果穗平均结实27粒。次年播种,能够长成正常二倍体植株。由此说明,单倍体的雌穗产生了正常的可育雌配子,且就每一雌穗而言,总有一些小花是可育的,但是不同单倍体个体间雌花序育性的恢复程度却存在较大差异。可见,单倍体的雌穗育性并非其自交结实的限制因素。
表1 糯玉米单倍体育性田间观察试验

处理编号时间(年)地点播期(日/月)单倍体来源试验方式A2015三亚08/11吉糯13自然育性观察B2016长春Ⅰ30/04吉糯13自然育性观察C2016长春Ⅱ27/05吉糯13自然育性观察D2016长春Ⅲ22/06吉糯13自然育性观察E2016三亚10/11吉糯13等11份母本材料自然育性观察
注:Ⅰ代表长春早春播;Ⅱ代表长春晚春播;Ⅲ代表长春夏播,下同。
2.2 糯玉米单倍体雄穗育性的自然恢复
2.2.1 糯玉米单倍体血缘对雄穗育性恢复的影响
由图2可知,自然条件下,田间的单倍体植株能够正常抽雄,但是大部分雄穗均表现为不育。在少数可育的雄穗中,有时仅仅是几个花药或小穗能够散粉,有时是几个分枝恢复了育性,有时是大部分分枝甚至整个雄穗都能够散粉。
由表3可知,从吉糯13中诱导产生单倍体散粉株率最高,为32.47%;从D 6644中诱导产生的单倍体散粉株率最低,为5.80%;从糯玉米自交系98 W-2、W 6、BN 2以及杂交种万糯2000中诱导的单倍体的散粉株率分别为30.383%、19.81%、18.18%、12.16%;但从中糯1号、苏玉糯1号、JN 15、景白浚34、JN-1中诱导出的单倍体散粉株率均未超过10%。由此可知,不同血缘的糯玉米单倍体雄穗育性恢复程度存在较大差异。本试验中11份不同来源的糯玉米单倍体雄穗散粉株率平均为14.29%,远远低于雌穗(61.25%)的育性,可见雄穗育性是糯玉米单倍体自交结实的限制因素。
表3 不同来源糯玉米单倍体雄穗育性的自然恢复

单倍体的来源单倍体株数(株)散粉株数(株)散粉株率(%)中糯1号176137.39苏玉糯1号250187.20吉糯131946332.47万糯20001481812.1698W-21334130.83W61062119.81BN2881618.18D66446945.80JN158877.95景白浚34127129.45JN-110165.94平均14.29
2.2.2 生长环境对糯玉米单倍体雄穗育性恢复的影响
对于同时从吉糯13中诱导产生的单倍体而言,不同的生长环境对其散粉株率有一定影响。由表4可知,以长春晚春播(5月27日)单倍体的散粉株率最高,达到14.75%;其次是长春夏播(6月22日),为13.99%;而海南三亚冬播(11月上旬)播种的单倍体散粉株率最低,均值为5.93%,与长春早春播(4月30日)的散粉率(5.77%)类似。
从5个播种时期单倍体的生长环境来看,昼夜温差有所不同。各个播期单倍体昼夜温差值依次为:三亚冬播(5.5 ℃)<长春早春播(8 ℃)<长春夏播(11 ℃)<长春晚春播(12 ℃)。这一顺序与单倍体散粉株率的变化相一致,表明糯玉米单倍体雄穗育性的恢复与环境温度有关,生长期间昼夜温差较小对单倍体雄穗育性的恢复不利,而昼夜温差较大有利于其雄穗育性的恢复。
表4 不同生长环境下糯玉米单倍体雄穗育性观察试验

时间(年)地点播期(日/月)播种昼夜温度(℃)单倍体株数(株)散粉株数(株)散粉株率(%)2015三亚08/1130/2410465.772016长春Ⅰ30/0423/15191178.902016长春Ⅱ27/0530/181832714.752016长春Ⅲ22/0629/181932713.992016三亚10/1126/2111875.93平均9.87
注:2016年长春3种播期供试糯玉米单倍体籽粒数分别为200粒,表中糯玉米单倍体株数为田间除杂后实际统计数目;2015、2016年三亚冬播供试糯玉米单倍体籽粒数均为120粒,表中糯玉米单倍体株数为经田间除杂后实际统计数目。
3 讨论与结论
本试验中11份不同来源糯玉米单倍体间雄花序育性的恢复程度存在巨大差异,雄穗散粉株率变幅为4.41%~32.12%,平均值为11.01%,雄穗育性恢复程度远远低于雌穗(61.25%),是糯玉米单倍体自交结实的限制因素,这与Bordes等[10]、Chalyk[11]、Shatskaya等[12]关于在普通玉米的试验结论一致。Shatskaya等认为,玉米单倍体雄穗散粉株率的差异是受核基因控制的,且呈加性遗传,对于雄穗可育率高的单倍体类型,易获得较多的加倍单倍体(自交后代)[12]。曹孜义等指出,变温处理有利于提高细胞分裂的同步化程度[13]。本试验结果与上述学者关于普通玉米的研究结论基本一致,本研究中糯玉米单倍体植株在昼夜温差较大的环境条件下种植,雄穗散粉株率明显偏高,变幅为13.68%~14.44%,而在昼夜温差较小的环境下种植,糯玉米单倍体散粉株率明显偏低(4.95%~8.51%),表明温差大的环境有利于单倍体雄穗育性恢复。雄穗育性恢复的基因型差异和环境变异性表明,在育种实践中可以选择育雄性恢复能力强的材料重点开展糯玉米DH育种,也可以选择适宜雄穗育性恢复的地点专门用于糯玉米单倍体的自然加倍,即糯玉米DH育种的基地化[14-15]。
参考文献:
[1]宋同明.糯玉米与WX基因[J].玉米科学,1993,1(2):1-2,25.
[2]刘纪麟.玉米育种学(第二版)[M].北京:中国农业出版社,2002:172-177.
[3]袁吉.糯玉米主要品质性状遗传规律研究[D].长春:吉林农业大学,2012.
[4]李艳茹,吉士东,郑大浩,等.糯玉米的营养价值和发展前景[J].延边大学农学学报,2003,25(2):146-148.
[5]王义发,沈雪芳,张璧,等.糯玉米种质资源的评价和创新育种[J].上海农业学报,2003,19(3):16-19.
[6]陈绍江,黎亮,李浩川,等.玉米单倍体育种技术(第2版)[M].北京:中国农业大学出版社,2012,3.
[7]李向永,姜龙,王薪淇,等.糯玉米单倍体诱导和加倍的研究[J].西北农林科技大学学报(自然科学版),2016,44(8):91-96.
[8]Chase S.S.Monoploid frequencies in a commercial double cross hybrid maize and its component single cross hybrids and inbred lines[J].Genetics,1949,34:328-332.
[9]景桂昕,姜龙,王薪淇,等.东北地区几个糯玉米DH系单株产量的配合力分析[J].种子,2015,34(9):82-85.
[10]Bordes J.G.,Charmet R.,Dumas de Vaulx,et al.Doubled haploid versus S1 family recurrent selection for testcross performance in a maize population[J].Theor.Appl.Genet.2006,112:1 063-1 072.
[11]Chalyk S.T.,Bylich V.G.,Chebotar O.D.Transgressive segregation in the progeny of a cross between two inducers of maize maternal haploids[J].Maize Genet Coop Newslett,1994,68:47.
[12]Shatskaya O.A.,Zabirova E.R.,Shcherbak V.S.,et al.Mass induction of maternal haploids[J].Maize Genet Coop Newslett,1994,68:51.
[13]曹孜义.玉米单倍体胚性细胞无性系二倍化研究[J].遗传学报,1983,10(4):274-279.
[14]邓昆鹏,李向永,景桂昕,等.高效糯玉米单倍体加倍方法的研究[J].种子,2017,36(1):82-86.
[15]刘志增,宋同明.玉米单倍体雌雄育性的自然恢复以及染色体的化学加倍[J].作物学报,2000,26(6):947-952.
猜你喜欢
青海农技推广(2022年1期)2022-05-05
南方农业(2021年30期)2021-12-25
热带农业工程(2021年2期)2021-06-25
河南农业科学(2021年4期)2021-05-24
热带农业科技(2020年4期)2020-10-23
山西农业科学(2020年8期)2020-08-13
北京青年周刊(2019年28期)2019-08-02
落叶果树(2018年6期)2018-11-28
河南农业科学(2018年5期)2018-01-19
优雅(2017年10期)2017-10-16
