
重金属胁迫下青藏高原芥菜型油菜种子发芽特征研究
2018-05-21 06:56:05,,,,
种子 2018年4期
, , , ,
(1.西藏高原生态研究所, 西藏 林芝 860000; 2.西藏农牧学院动物科学学院, 西藏 林芝860000; 3.西藏农牧学院资源与环境学院, 西藏 林芝860000)
矿产资源的不合理开发和管理体制的不健全等诸多因素,使土壤重金属污染在全球呈现越来越严重的趋势,土壤重金属污染不但影响着土地资源和水资源,还关系着生态、环境、农畜产品安全等诸多方面。因此,治理土壤重金属污染已经成为国际科学界研究的热点问题之一[1]。西藏是我国西南边陲的重要生态安全屏障,素有“世界屋脊”和地球“第三极”之称,随着矿产资源的采冶,矿区周边重金属污染问题越来越严重[2-4]。近年来,在重金属污染的治理中,利用绿色植物来容纳、转移或转化污染物的植物修复技术已被越来越多的研究学者所重视[5-9]。植物修复技术的基础是从众多植物中培育和筛选出具重金属耐性的积累植物或超积累植物。因此,从矿区废弃地中筛选适宜当地环境特点的植物是修复矿区生态环境的重要途径[4]。目前在重金属积累植物筛选中多注重于单一重金属对种子发芽及根芽长的影响,采用多种不同重金属对种子发芽及根芽长影响的报道相对较少[10-11]。
在Cu、Fe、Mn、Zn、Pb等5种元素中,Cu、Fe、Mn、Zn是植物生长必需的微量元素,Cu元素是构成植物某些氧化酶的重要成分,又参与植物光合作用的电子传递,影响CO2的氧化还原过程[12];Fe是植物必需的微量元素之一,在植物体内含量较少,但它是形成叶绿素所必需的元素之一;Mn在植物中的生理作用上是多方面的,是多种酶的组成和活化剂,与许多酶的活性有关,也能促进碳水化合物、氮的代谢,直接关系植物的生长发育和生物量;Zn是已醇脱氢酶,谷氨酸脱氢酶等的活化剂,植物缺少Zn元素则导致植物株型等性状发生改变[13-15]。但是无论是植物的非必需元素还是必需元素,当Cu、Fe、Mn、Zn、Pb这些元素超过一定含量时,会紊乱植物的代谢过程,阻碍植物的生长发育,甚至导致植物死亡[16-17]。然而,如芥菜型油菜(Brassicajuncea)等植物体内具有某些特定生理机制,使植物能在较高浓度含量的重金属中生存且不受损害[18]。
芥菜型油菜(Brassicajuncea)属于十字花科植物,具有耐旱、抗寒、生物量大、青贮效果好等特征,可富集Cu、Pb、Ni、Cd和Zn等多种重金属[5-6],并且对这些重金属污染的土壤具有较显著的净化作用,在重金属污染矿区土壤修复中可能有一定的应用前景[19]。
基于以上原因,以青藏高原常见的乡土植物——芥菜型油菜为研究对象,探讨在5种不同重金属离子浓度胁迫下,种子发芽及根芽长所受的影响,为在受重金属污染土壤中植物种植的适宜性和进一步了解青藏高原重金属矿区植物的抗性机理以及研究修复重金属污染土壤功能提供基础性依据[20-21]。
1 材料与方法
1.1 材料来源
芥菜型油菜(Brassicajuncea)种子2016年9月采集于西藏自治区拉萨市当雄县的拉乌矿区(Cu-Zn-Pb矿)尾矿库复垦区。该矿山作业区海拔4 300 m以上,尾矿库复垦区海拔4 630 m,复垦采用先锋植物披碱草(ElymusdahuricusTurcz.),油菜为伴生植物。试验选取饱满均匀的油菜种子,弃去杂质以及成熟度较低、有虫蚀、干瘪等的种子,采用0.1%的KMnO4溶液浸泡10 min,然后用去离子水冲洗多次,直至KMnO4红色完全消失,然后用滤纸将水吸干。将消毒后的种子均匀放置于铺有双层滤纸床的发芽盒中,每盒均匀放入100粒种子。依据自然界土壤中重金属背景值和前人研究成果,本试验共设置5个重金属处理组,重金属Cu、Fe、Mn、Zn、Pb分别采用分析纯CuSO4·5 H2O、ZnSO4·7 H2O、PbCl2、FeCl3、MnSO4,不同浓度梯度配见如表1,每个处理重复3次,将配好的中重金属溶液浓度依次加入培养皿中,每皿3 mL,每个处理重复3次,同时以去离子水处理为作为对照(ck)。每个重金属处理组设置不同的5个浓度梯度,所有处理均在全自动恒温光照培养箱内培养,温度为20 ℃,光照时间为16 h(昼)/8 h(夜),相对湿度为80%~90%。试验期间逐日观察记录种子的萌发数(胚根与种子等长、胚芽长度达到种子一半即认为种子已经发芽)。
表1 试验中配制的不同浓度5种重金属溶液
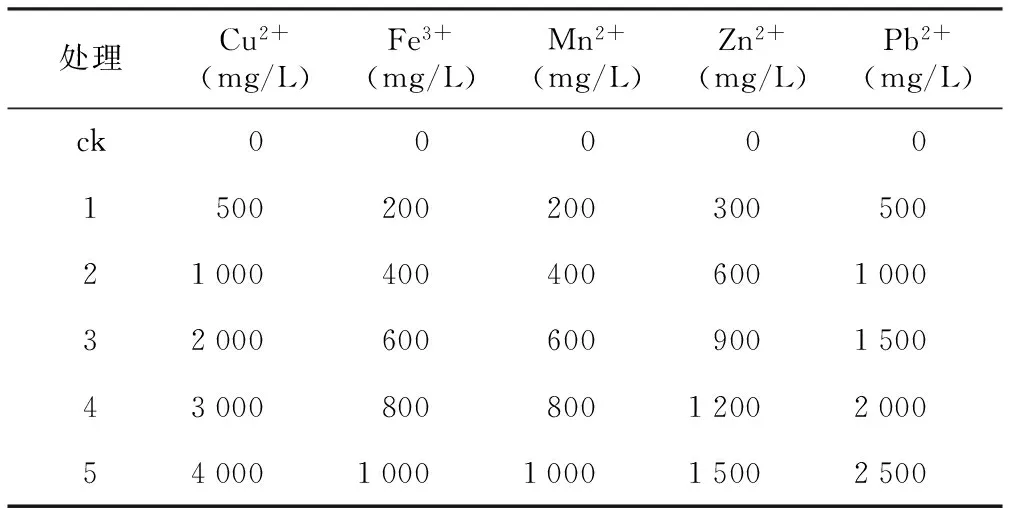
处理Cu2+(mg/L)Fe3+(mg/L)Mn2+(mg/L)Zn2+(mg/L)Pb2+(mg/L)ck0000015002002003005002100040040060010003200060060090015004300080080012002000540001000100015002500
1.2 测定方法
种子的发芽势和发芽率是种子活力的重要指标,也是种子的重要生产性能。种子发芽势、发芽率采用常规统计方法;每处理重复3次,随机选取20株幼苗,采用精度为0.1 mm的游标卡尺分别测量胚芽长、胚根长,求平均值。为保证处理浓度稳定,每隔24 h更换1次处理液。处理第4天时统计种子的发芽势;第7天时统计种子的发芽率,测量胚芽长、胚根长,求平均值。
发芽率(%)=供试种子的发芽数/供试种子总数×100%;
发芽势(%)=4 d内供试种子的发芽数/供试种子总数×100%。
1.3 数据处理与分析
采用Excel 2007进行数据处理和图表绘制,并采用SPASS 23.0软件进行单因素方差分析和最小显著差数法(LSD法)进行多重比较。
2 结果与分析
2.1 Cu2+浓度胁迫对芥菜型油菜种子发芽及根芽长的影响
由图1可知,不同浓度的Cu2+胁迫对油菜种子萌发均有一定的影响。油菜种子在5种浓度梯度胁迫下的发芽率、发芽势均低于对照;Cu2+浓度为500 mg/L时,油菜种子发芽率、发芽势分别达到89.3%、85%,与对照组差异不显著(p>0.05),表现出Cu2+对油菜种子萌发有一定的耐性;当Cu2+浓度为4 000 mg/L时,油菜发芽率、发芽势分别降低为33.3%、16.1%,表现出对油菜种子萌发明显的抑制效应,与对照组差异极显著(p<0.01)。总体趋势来看,油菜种子在Cu2+浓度胁迫下,随着浓度梯度的增加,平均发芽率和发芽势逐渐降低,对种子萌发的抑制作用逐渐增强;当Cu2+浓度≥4 000 mg/L时,种子萌发率最低,抑制作用最强。
由图2可知,Cu2+浓度胁迫对油菜种子根芽长的生长具有明显的抑制作用。Cu2+浓度在500 mg/L时,平均根长4.2 mm、平均芽长10.2 mm,根芽长均显著低于对照组,说明在此浓度下Cu2+对油菜根芽长已经产生明显的抑制作用。随着Cu2+浓度的增加,对油菜种子根芽长的抑制作用逐渐增大。Cu2+浓度达到1 000 mg/L时,油菜只长芽不长根,出现明显的无根现象;当Cu2+浓度≥1 000 mg/L时,抑制作用达到最大,已超过种子萌发的耐受性,芽长和根长均无生长,导致种子死亡。Cu是作物生长的必需元素之一,但浓度过高直接影响植物的正常生长。
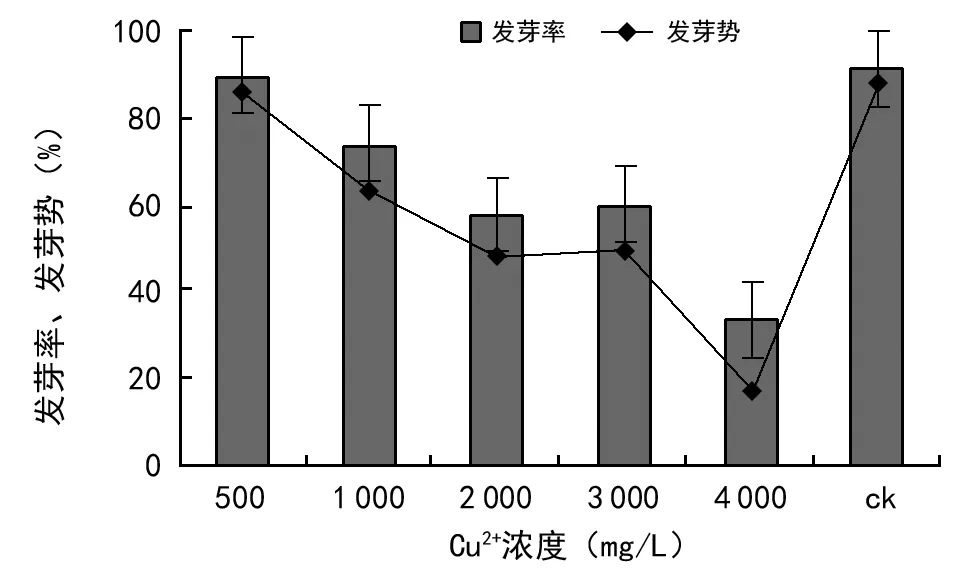
图1 Cu2+浓度胁迫对油菜种子发芽率、发芽势的影响
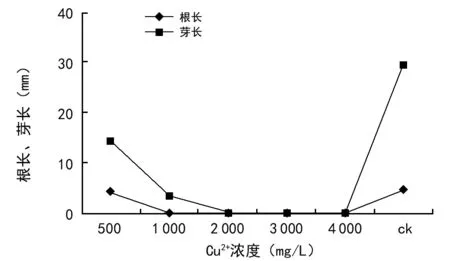
图2 Cu2+浓度胁迫对油菜种子根长、芽长的影响
2.2 Fe3+浓度胁迫对芥菜型油菜种子发芽及根芽长的影响
由图3可知,不同浓度的Fe3+处理对油菜种子萌发均有明显的影响。Fe3+浓度为200 mg/L时,油菜种子萌发率为92%、发芽势为91.3%,比对照分别高0.83%和4.50%,发芽率与对照组差异不显著(p>0.05),发芽势与对照差异达到显著水平(p<0.05)。当Fe3+浓度为400~1 000 mg/L时油菜种子发芽率均低于对照组,且随着Fe3+浓度的增加发芽率和发芽势均存在明显的下降的趋势。说明高浓度的Fe3+胁迫对油菜种子萌发有明显的抑制。经方差分析,Fe3+浓度为400~600 mg/L时,油菜的发芽率和发芽势均显著低于对照(p<0.05);当Fe3+浓度达到600 mg以上时,发芽率和发芽势较对照达到极显著水平(p<0.01),但还未达到致死的程度。Fe是植物必须的微量元素之一,在种子发芽时参加细胞的呼吸作用,对种子萌发有一定的刺激效应[13]。总体来看,低浓度的Fe3+对油菜种子的发芽有一定的促进作用,Fe3+浓度大于200 mg/L时随着浓度的增加对油菜种子萌发抑制作用逐渐增大,发芽率、发芽势呈现出下降的趋势,说明不同浓度的Fe3+离子对油菜种子的萌发所起作用不同,超过1 000 mg/L时抑制作用明显。
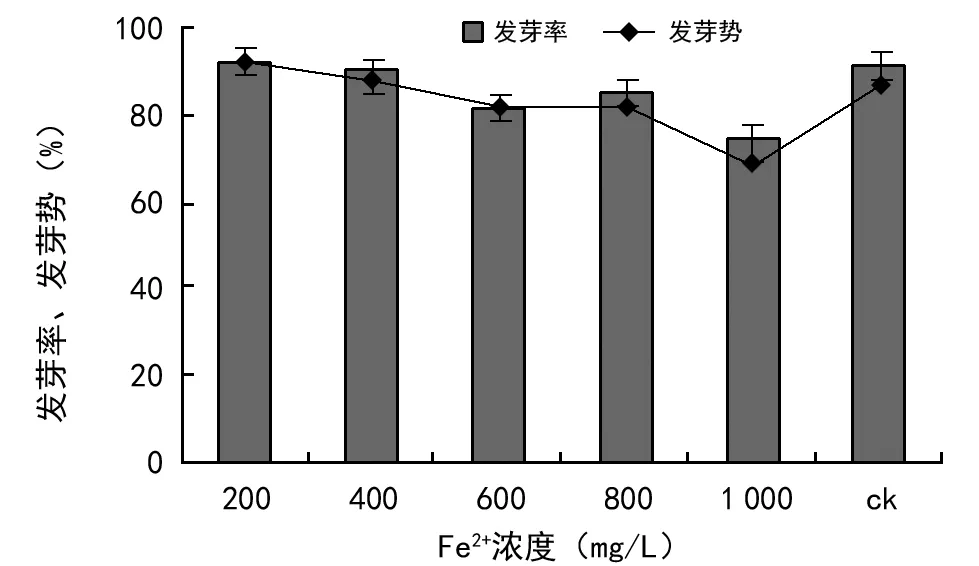
图3 Fe3+浓度胁迫对油菜种子发芽率、发芽势的影响
由图4可以看出,当溶液中Fe3+浓度为200~1 000 mg/L时,油菜种子根长和芽长均低于对照组,当溶液中Fe3+浓度为600 mg/L时油菜的根长最短,仅为2.3 mm、芽长13.6 mm。与大多数研究的结果相似,Fe3+对根长影响明显强于对芽长的影响[11,22-23]。最主要的原因是植物种子萌发后,根最先突破种皮吸水,从而使根的重金属累积量以及受胁迫时间比胚芽大[22-23]。本研究中,Fe3+胁迫既抑制了油菜幼苗根的生长,也抑制了芽的生长。可见,高浓度的Fe3+胁迫对油菜种子的萌发有一定的抑制作用,对根芽的生长抑制作用更为明显[24-25]。
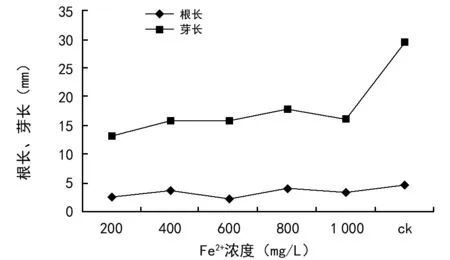
图4 Fe3+浓度胁迫对油菜种子根长、芽长的影响
2.3 Mn2+浓度胁迫对芥菜型油菜种子发芽及根芽长的影响
Mn也是植物必需的矿质元素之一,通常土壤中的Mn元素以Mn2+、Mn3+、Mn4+3种氧化态存在,此外还以螯合状态存在[26],但被植物吸收的主要状态是Mn2+。研究表明,Mn2+能促进种子萌发,是由于Mn2+对刺激生长素促进胚芽鞘伸长有积极效应[27]。由图5可知,Mn2+离子对油菜种子发芽率和发芽势都有明显且积极的影响,在200~1 000 mg/L处理条件下油菜种子发芽率、发芽势均高于对照组,虽然发芽势随着浓度的增大呈现逐渐下降趋势,但各处理发芽率均高于对照。Mn2+浓度在400 mg/L时发芽率达到最高,为95.66%,比ck的平均发芽率上升4.49%,说明Mn2+浓度胁迫对油菜种子萌发表现出促进效应,且差异显著(p<0.05)。从发芽势来看,当Mn2+浓度大于400 mg/L时,随浓度的增加呈现下降的趋势,说明浓度的进一步增加会逐渐对油菜种子萌发速度产生抑制效应。
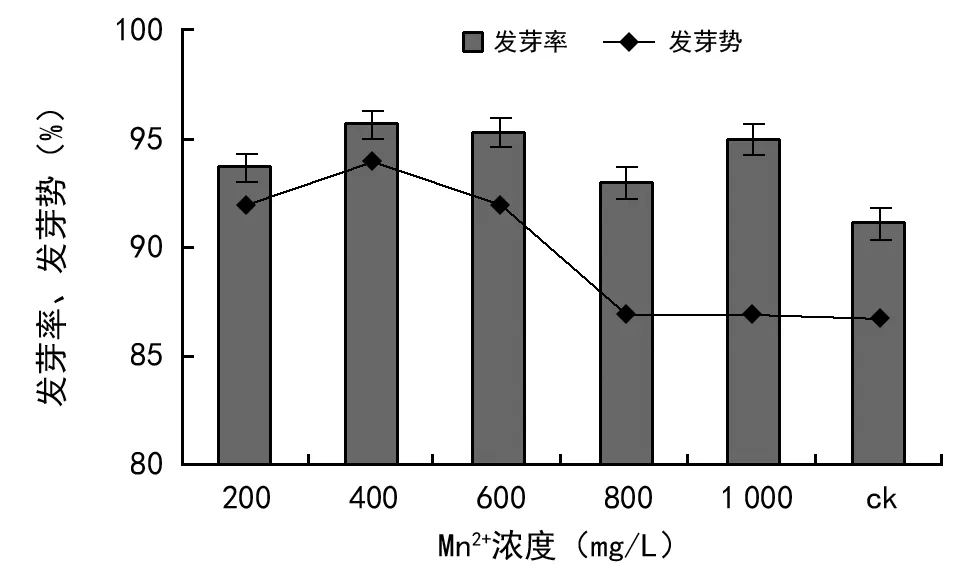
图5 Mn2+浓度胁迫对油菜种子发芽率、发芽势的影响
如图6所示,Mn2+浓度胁迫对油菜种子根芽长生长也有明显的影响。Mn2+浓度在200~1 000 mg/L时,油菜种子的根芽长均低于对照组,说明Mn2+浓度胁迫对油菜种子根芽长生长有明显的抑制效应。且随着Mn2+浓度梯度的不断增加,抑制作用越来越强,甚至导致油菜死亡。
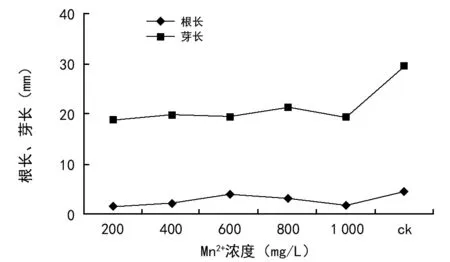
图6 Mn2+浓度胁迫对油菜种子根长、芽长的影响
2.4 Zn2+浓度胁迫对芥菜型油菜种子发芽及根芽长的影响
Zn2+是植物必须的微量元素之一,是已醇脱氢酶、谷氨酸脱氢酶等多种酶的活化剂,通常能促进种子的萌发[12]。如图7所示,不同浓度的Zn2+对油菜种子萌发均具有一定的促进作用。Zn2+浓度为300 mg/L时,平均发芽率最高,达到98.67%;平均发芽势也达到最高的97%,分别高出对照组7.5%和10.2%,两者与对照相比均达极显著水平(p<0.01)。在600,900,1 200,1 500 mg/L浓度下油菜发芽率及发芽势达到97%、94%以上,均高于对照组,在300 mg/L和1 500 mg/L、600 mg/L和1 200 mg/L时发芽势差异不显著,但也高于对照4个百分点以上,说明在此浓度下Zn2+浓度胁迫对油菜种子萌发无明显的抑制作用,且发芽率、发芽势影响趋势相似。
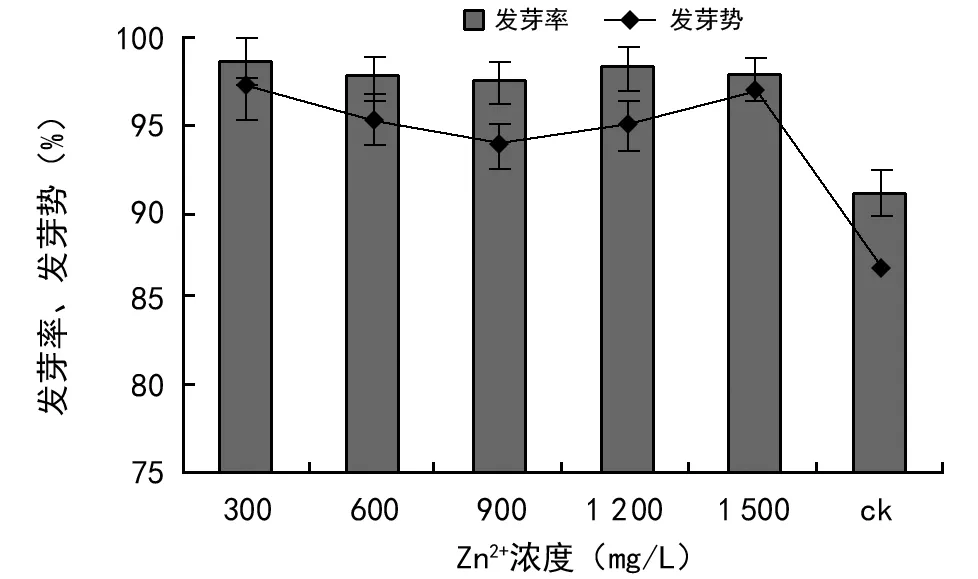
图7 Zn2+浓度胁迫对油菜种子发芽率、发芽势的影响
由图8可知,低浓度的Zn2+对根芽长影响效果不一致。当Zn2+浓度为300 mg/L时油菜根长高于对照组3.85 mm,但在此浓度下油菜芽长低于对照芽长6.4 mm。之后随着Zn2+浓度的增加,开始出现抑制作用,Zn2+浓度≥600 mg/L时,对根芽长抑制作用开始加强,呈现显著下降的趋势,平均根芽长逐渐减小,浓度进一步增加将会导致油菜种子死亡。
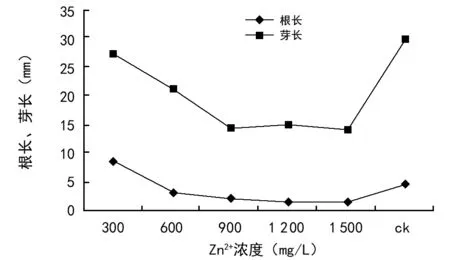
图8 Zn2+浓度胁迫对油菜种子根长、芽长的影响
2.5 Pb2+浓度胁迫对芥菜型油菜种子发芽及根芽长的影响
由图9可知,Pb2+浓度为500 mg/L时,发芽率和发芽势均达到最高,发芽率率达到94.67%、发芽势达到93.30%,高出对照组3.5%和6.5%。在1 000,1 500,2 000,2 500 mg/L时发芽率均低于对照组,说明高浓度抑制了种子的萌发,且浓度越大抑制作用越强。从图9可知,油菜的发芽率大小顺序为500 mg/L>ck>2 000 mg/L>1 000 mg/L>1 500 mg/L>2 500 mg/L;说明Pb2+在低浓度胁迫时对油菜种子萌发有促进效应,在高浓度时出现抑制作用,且浓度≥2 500 mg/L时种子发芽率和发芽势均最低,抑制作用最为明显。
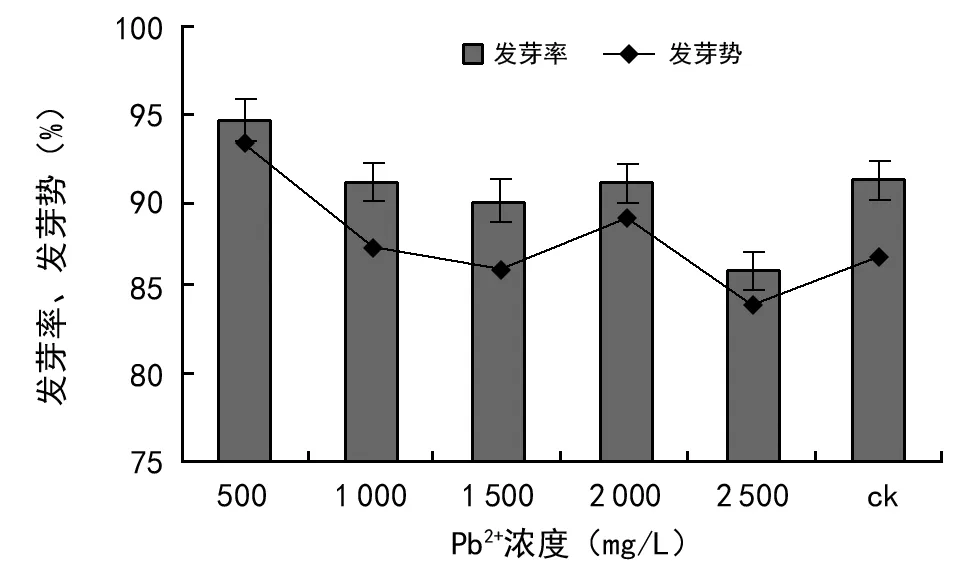
图9 Pb2+浓度胁迫对油菜种子发芽率、发芽势的影响
环境中对植物、动物以及人类毒性最强的重金属之一就是铅。目前发现的铅超积累植物在自然界植物中只有少数几种[28]。由图10可知,Pb2+浓度胁迫对油菜种子根芽长具有明显的抑制作用。在Pb2+胁迫下,油菜种子根长和芽长均显著低于对照(p<0.05)。随着溶液中Pb2+浓度的增加,芽长从对照的24.9 mm缩短到7.7 mm,根长从对照的4.65 mm缩短到1 mm,说明Pb2+胁迫抑制了油菜种子的根芽的伸长,且抑制作用越来越强,甚至导致油菜种子死亡。
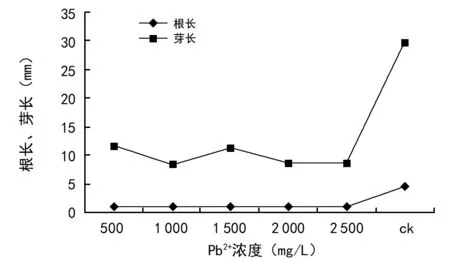
图10 Pb2+浓度胁迫对油菜种子根长、芽长的影响
3 结论与讨论
Cu、Fe、Mn、Zn是植物生长和发育的必需元素,缺少这些元素时植物通常会出现缺素症,但是在土壤中这几种元素超过一定限度时,作物生长也会受到严重损害。作物生长受损后对水分和养分的吸收就会受到影响,致使发育不良甚至死亡[18]。Pb并非植物的必需元素[29],有毒性,超过一定限度会对作物产生毒害。一般情况下,Cu2+、Fe3+、Mn2+、Zn2+对种子萌发抑制效应较为明显,并且随着浓度增加现象更加显著,但由于试验设置的Cu、Pb浓度过高,直接导致油菜种子受毒害严重,甚至导致种子死亡,所以超过土壤质量标准的设置梯度对种子萌发是不适用的。Fe3+是植物必须的微量元素之一,在种子发芽时还参与细胞的呼吸作用,它也是一些植物酶的重要成分。所以试验中Fe3+浓度梯度相对较低,总体来看,不同浓度Fe3+处理下,油菜种子的发芽率、发芽势都较高,主要是由于当较低浓度Fe3+能够促进植物的生长,并且随浓度的增加萌发抑制作用逐渐增大,Fe3+浓度≥1 000 mg/L时抑制作用最强。Mn能促进种子萌发,因为它刺激生长素促进胚芽鞘伸长有积极的效应[27]。Mn2+设置浓度梯度与Fe3+一致,发芽率、发芽势均高于对照组,但对油菜种子发芽率比Fe3+要高,说明Mn2+促进种子萌发能力比Fe3+要强,随浓度的增加都出现了相同的抑制作用;Zn2+也是植物必须的微量元素之一,是谷氨酸脱氢酶、已醇脱氢酶的活化剂,能促进种子的萌发。Zn2+浓度胁迫时种子的平均发芽率、发芽势均高于对照组,且平均发芽率≥97%,Zn2+浓度≤1 500 mg/L时对种子萌发具有促进作用。Pb2+在500 mg/L时对油菜种子发芽率高于对照组,说明低浓度胁迫对油菜种子萌发有促进效应,在高浓度时出现抑制作用,且浓度≥2 500 mg/L时抑制作用最明显,与Cu2+、Fe3+、Mn2+、Zn2+重金属胁迫对油菜种子低促高抑萌发现象基本类似。对青藏高原重金属矿区植物的抗性机理以及在污染土壤中掌握它的耐受度还需进一步研究。
本试验结果与其他的研究结果相似,即重金属对植物种子萌发及幼苗的影响大多存在一个高浓度下的抑制效应和低浓度下的促进效应[29-30]。而造成低促高抑现象的原因可能是低浓度提高了种子胚的生理活性,促进萌发;而高浓度伤害了种子的胚、芽等,并且抑制了蛋白酶、淀粉酶的活性,即高浓度的重金属离子使得种子内贮藏淀粉和蛋白质的分解受到抑制,进而影响到种子萌发,导致种子萌发受到抑制[31]。油菜对Cu2+、Pb2+的吸收能力相对较高,在较高浓度下还可以成活。在相同浓度下,Mn2+对油菜种子的发芽率影响要高于Fe3+。在不同浓度Zn处理下,对油菜种子发芽率有明显的刺激作用,在浓度低于1 500 mg/L时表现出发芽率和发芽势均高于对照的现象,但对油菜种子根芽长的生长抑制明显,浓度过高使种子生长受毒害严重,超过植物的耐受限度就会直接导致死亡。本试验结果表明,低浓度的Cu2+、Fe3+、Mn2+、Zn2+、Pb2+能在一定程度上促进发芽和根芽长的生长,但所受的浓度各不同、程度也不一样;高浓度的Cu2+、Fe3+、Mn2+、Zn2+、Pb2+均明显抑制发芽和根芽长的生长,但所受的浓度各不同,浓度过高致使种子受毒害严重,在较高浓度的重金属影响下,油菜种子的萌发较为缓慢、生长速度缓慢,根长和芽长也相对较低。总体来看,油菜能够在一定浓度的重金属污染环境中生存,具有对重金属一定的抗性,因此,可以考虑作为一种重金属富集植物对西藏矿区污染土壤进行修复。
参考文献:
[1]Chaney RL,Malik M,Li YM,et al.Phytoremediation of soil metals[J].Current Opinions in Biotechnol-ogy,1997,8:279-284.
[2]敬久旺,赵玉红,张涪平,等.藏中矿区表层土壤重金属污染评价[J].贵州农业科学,2011,39(7):126-128.
[3]张涪平,曹凑贵,李苹,等.藏中矿区重金属污染土壤的微生物活性变化生态学报,2010,30(16):4 452-4 459.
[4]赵玉红,拉巴曲吉,罗布,等.铜、镉、铅、锌对4种豆科植物种子萌发的影响[J].种子,2017,36(1):22-28.
[5]吴新民,李恋卿,潘根兴,等.南京市不同功能区土壤中重金属Cu、Zn、Pb、和Cd污染特征[J].环境科学,2003,24(3):105-111.
[6]郑袁明,余柯,吴乱涛,等.北京市城市公园土壤铅含量及其污染评价[J].地理研究,2002,21(4):418-424.
[7]MantaDS,AngeloneM,BellaneaA,et al.Heavy metel inurban soil:A case study from the city of Palermo(Siclily),Ita-ly[J].The Science of the Total Environmental,2002,300(1/3):229-243.
[8]张银龙,陆亚芳,王亚超.大气污染梯度下树木附生苔鲜植物生理生化指标的变化[J].北京林业大学学报(自然科学版),2006,30(5):5-9.
[9]陈同斌,韦朝阳,黄泽春.砷超富集植物蜈蚣草及其对砷的富集特征[J].科学通报,2002,47(3):207-210.
[10]黄耿磊,黄冬芬,刘国道,等.镉胁迫对3种柱花草生长及植株镉积累和分配的影响[J].草地学报,2011(191):97-101.
[11]陈平,余土元,叶丽敏.镉胁迫对弯叶画眉草幼苗生长和生理特性的影响[J].草地学报,2002,10(3):212-216.
[12]周青,黄晓华,彭方晴,等.镧-甘氨酸配合物对镉伤害小白菜的影响[J].环境科学,1999,20(1):91-94.
[13]王振河,汤菊香,代海芳,等.锰和锌对盐渍土中水稻幼苗生长的影响[J].湖北农业科学,2007,46(4):547-549.
[14]王宝山.植物生理学[M].北京:科学出版社,2007.
[15]魏海姆,F H.植物生理学实验[M].北京:科学出版社,1974.
[16]王艳,辛士刚,马莲菊,等.翦股颖和高羊茅对铜、铅吸收及耐受性[J].应用生态学报,2007,18(3):625-630.
[17]段昌群,王焕校.重金属对蚕豆的细胞遗传学毒理作用和对蚕豆根尖微核技术的探讨[J].植物学报,1995,37(1):14-24.
[18]杨洋,黎红亮,陈志鹏,等.郴州尾矿区不同油菜品种对重金属吸收积累特性的比较[J].农业资源与环境学报,2015,32(4):370-376.
[19]侯艳伟,池海峰.毕丽君,等.生物炭施用对矿区污染农田土壤上油菜生长和重金属富集的影响[J].生态环境学报,2014,23(6):1 057-1 063.
[20]张震,徐丽,杨洁,等.重金属胁迫对高羊茅种子萌发的影响[J].中国农学通报,2008(04):386-389.
[21]Naresh Kumar Agarwal,Harrison Lawrence.Office Location Map of Individuals in the Library and Other College Campus Buildings:A Proof-of-Concept Wayfinding System[J].Journal of Web Librarianship,2014,8(3).
[22]慈恩,高明,王子芳,等.镉对紫花苜蓿种子萌发与幼苗生长的影响研究[J].中国生态农业学报,2007(15):96-98.
[23]梅丽娜,袁庆华,姚拓,等.不同品种苜蓿芽期对重金属镉的耐性研究[J].作物杂志,2010(2):15-18.
[24]周启星,复合污染生态学[M].北京:中国环境科学出版社,1995,18(3):24-29.
[25]丁园、宗良纲,不同土壤重金属复合污染有效态离子冲量表征[J],污染与防治,2003,6(25):173-178.
[26]傅家瑞,宋松泉,黄上志,等.植物生理学[M].广州:中山大学出版社,2005.
[27]陆景陵.植物营养学(上册)[M].北京:中国农业大学出版社,2003.
[28]李红霞,马伟芳,赵新华.浅述利用植物对受污染土壤中重金属的修复[J].吉林农业科学,2007,32(1):35-39.
[29]多立安,高玉葆,赵树兰.重金属递进胁迫对黑麦草初期生长的影响[J].植物研究,2006,26(1):117-122.
[30]闫晓明,何金柱,苗青松.污染土壤植物修复技术研究进展[J].中国生态农业学报,2004,12(3):131-133.
[31]葛成军,陈秋波,俞花美,等.Cd胁迫对2种热带牧草种子发芽与根伸长的抑制效应[J].热带作物学报,2008,29(5):567-571.
猜你喜欢
现代园艺(2020年19期)2020-10-02 03:15:48
湖北农业科学(2019年20期)2019-12-11 10:03:58
阅读与作文(初中版)(2019年8期)2019-08-27 08:18:28
种子(2019年6期)2019-07-13 07:45:16
种子(2019年4期)2019-05-28 02:04:14
现代农村科技(2019年4期)2019-01-06 14:13:19
农业与技术(2018年5期)2018-04-25 10:57:04
农产品加工(2017年19期)2017-11-14 12:30:17
北方文学(2017年22期)2017-08-25 07:29:07
山东农业科学(2016年1期)2016-05-30 10:48:04
