
Ecosystem carbon and nitrogen storage following farmland afforestation with black locust(Robinia pseudoacacia)on the Loess Plateau,China
2018-05-19 03:36GuangqiZhangPingZhangYangCao
Journal of Forestry Research 2018年3期
Guangqi Zhang•Ping Zhang•Yang Cao
Introduction
Afforestation of agricultural land has been proposed as an effective method of carbon(C)sequestration because of the rapid accumulation and long-term potential for C storage in plant biomass and soil(Jackson and Schlesinger 2004;Metz et al.2007;Sang et al.2013).In 1999,the government of China started the Grain for Green program(GGP)to control soil erosion and increase vegetation cover on degraded lands.The GGP has converted over 20 million hectares of steeply sloping(greater than 25 degrees),degraded farmland and barren land into primarily treebased plantations.This has increased the magnitude and distribution of C sequestration at regional and national scales.Some 246 Mt C were sequestered during the first decade of GGP(1999–2008;Persson et al.2013),and 524.36 Mt C could be sequestered by the end of this century(Liu et al.2014).On the Loess Plateau,the amount of C sequestered was 96.2 Mt C for the period 2000–2008 and soil C stocks increased at a rate of 0.71 Mt C year-1in the top 20 cm soil layer(Chang et al.2011;Feng et al.2013).
It is generally accepted that during stand development,ecosystem C sequestration increases due to an increase in plant biomass(Chen et al.2013;Zhang et al.2014).However,soil is the largest terrestrial C pool,accounting for over two-thirds of C stored in forest ecosystems(Arevalo et al.2009;Dixon et al.1994).A slight change in the soil C pool can greatly in fluence ecosystem C cycling and atmospheric CO2concentrations(Davidson and Janssens 2006).Numerous studies have improved our understanding of the effects of in fluencing factors on C sequestration after land use change,including previous land-use type,soil texture,tree species,stand age,and management practices at global,regional,and local scales(Deng et al.2013;Jo´zefowska et al.2017;Guo and Gifford 2002;Laganiere et al.2010;Post and Kwon 2000).Mazurek and Bejger(2014)found that the content and pools of organic C were greatest in samples taken from farmland closest to black locust trees and that this can be an important factor of C stock increments in soil.Liu et al.(2011)reported that soil C stocks under croplands were greater than under grasslands and forestlands.Song et al.(2014)showed that soil C accumulation was significantly greater after conversion of cropland to forest than after conversion of cropland to grassland.Pietrzykowski and Krzaklewski(2007)reported that reclamation in opencast sand mines significantly accelerate the development of soil-forming processes in the recreated ecosystem.Meta-analysis studies have shown that plant type in fluences soil C accumulation,and that pine plantations reduce soil C and N stocks by 15 and 20%,respectively(Berthrong et al.2009;Song et al.2014).Soil C concentrations are greater in plantations of N2- fixing species(e.g.,black locust)than in plantations of non-N2-fixing species(Paul et al.2002).Moreover,the productivity of the target trees steadily increases with plant growth among the N2- fixing species(Khanna 1997).Thus,it may be possible to accelerate soil C accumulation rates through the use of N2- fixing species,either through establishing plantations or understory vegetation(Paul et al.2002).
Stand age is a powerful predictor of ecosystem structure,function,and C storage in forest ecosystems(Bradford and Kastendick 2010;Pregitzer and Euskirchen 2004).Deng et al.(2014a)reported that stand age after afforestation had greater impact than temperature or precipitation on soil C stock changes after cropland conversion.Afforestation generally causes reduced soil C storage during the first few years,and is then followed by a gradual increase with stand development(Hu et al.2009;Laganiere et al.2010;Paul et al.2002;Ritter 2007).Mao et al.(2010)found that soil C and N stocks of poplar plantations in northeast China recovered to pre-afforestation levels after 15 years of being used as agricultural lands.In the hill-gully terrain of the Loess Plateau,planted perennial alfalfa caused a decline in soil C storage during the first 9 years of growth but C storage recovered to pre-plantation levels after 13 years,dropping again after 16 years(Deng et al.2014c).Soil C in Chinese fir plantations in natural forests decreased to a minimum level at 16 years after planting,re-accumulated between 16 and 21 years,and stabilized during the overmature stage,attaining an equilibrium soil C pool 30%lower than during pre-harvest levels(Chen et al.2013).C storage in deeper soil layers can provide a more complete picture of soil C dynamics(Chen et al.2013).Better understanding of the long-lasting effects of planted forests on soil pro file C stocks following afforestation is needed(Chen et al.2013;Cheng et al.2014;Zhou et al.2006).
It has been suggested that N is a key element in the regulation of terrestrial C sequestration(Luo et al.2004;Rastetter et al.1997).Yang et al.(2011)synthesized 124 studies from around the world and found that N changes exhibited linear increases with C changes in various ecosystem components,but not in mineral soils in forest ecosystems.In contrast,Li et al.(2012)pooled data from 292 sites and showed that changes in soil C and N stocks are correlated and have similar temporal patterns.Therefore,monitoring the temporal patterns of N stocks in plantation ecosystems might prove useful in helping us to understand the sustained long-term ecological processes and driving forces of the C cycle.This could provide insight into solutions for the improvement of forest C sequestration mechanisms(Fang et al.2001;Lal 2004;Li et al.2012).
Due to its early sprouting,fast growth,and high rate of N- fixation,black locust(Robinia pseudoacacia)has been widely planted on the Loess Plateau to aid in soil and water conservation,with more than 70,000 ha planted over the last few decades(Qiu et al.2010).Recent studies have investigated the effects of black locust afforestation on C and nutrients in plants and soil on the Loess Plateau(Li et al.2013a;Li and Liu 2014;Qiu et al.2010;Wang et al.2012).However,those studies did not provide a thorough depiction of C sequestration by black locust afforestation on farmlands.In order to provide further insight and data,the present study examined the changes in C and N pools in the tree components[herbs,litter,and soil(0–100 cm depth)]following afforestation of farmlands using an agesequence of a black locust plantation.
Materials and methods
Study site and experimental design
This study was conducted in the Zhifanggou watershed(36°46′28′′–36°46′42′′N, 109°13′03′′–109°16′46′′E,1010–1431 m a.s.l.),of the Yanhe River basin of Ansai County,North Shaanxi Province,China(Fig.S1).The climate is that of a transitional zone from a semi-humid warm climate to semiarid climate with an average annual precipitation of 504 mm(1970–2006)(Jiao et al.2012).Rains fall between July and September,accounting for 74%of annual rain fall.Mean annual temperature in the watershed ranges from 7.7 to 10.6°C and the average frost-free period is 157 days.The soils are classi fied as Calcic Cambisols according to the soil map of the world(FAO–UNESCO 1974).The most common soil in this watershed develops on loess soil and has a texture dominated by fine silt(0.05–0.002 mm)(Zhang et al.2004).This region is subject to severe soil erosion(Shi and Shao 2000)and gully erosion is well developed with gully density of 8.1 km/km2(Zhang et al.2004).Land degradation,deserti fication,soil erosion and declining soil fertility threaten the environment.However,after more than 30 years of management,the watershed shows signi ficant improvements in the ecological environment(Zhang et al.2007).
Sample collection and analyses
Black locust plantations aged 9,17,30,and 37 years were randomly selected in the whole watershed and each age class was represented by three plantations,all of which were afforested on steeply sloping farmland.We also sampled 1-to 2-year old abandoned farmlands that we categorized as ‘‘pre-afforestation sites’’for comparison(0 years old).The sites were all located near the tops of loess mounds and there were few differences among the sites in regard to aspect(south),gradient(28°–33°),elevation(1150–1260 m),and previous farming practices.The distance between each sampled stand ranged from 0.5 to 4 km.
A 20 m×20 m plot was established in the central area of each afforested site for sampling in August 2011.Diameter at breast height(DBH at 1.3 m height)and tree height(H)were measured for all trees in each plot.To establish allometric models for estimating tree biomass,35 black locust trees from the four black locust age-groups were sampled(7–9 trees per age-group selected based on DBH range).After the selected trees were felled,the total H of each tree was measured and then divided into 1-mlong sections.Each section was separated into stem,bark,leaf,or branch.The entire root system was manually excavated to a depth of 100 cm and soil particles were carefully removed.All components were dried at 70°C to a constant weight.Allometric regression equations relating tree DBH and H were developed across this chronosequence.The allometric regression equations were well fitted for each tree component(Table 1).These equations were used to calculate the biomass of each tree and the total biomass of each black locust stand.
Only the herb layer was present in the understory layer of all selected sites.All herb species were identi fied by two observers working together by randomly selecting five 1 m×1 m quadrants at each stand.Litter and herb biomass,including roots,were also harvested from these five subplots.All samples including tree tissues(leaves,branches,stems,and roots),herbs,and litter were weighed and oven-dried at 65°C to a constant weight in the lab and reweighed through wet-to-dry mass conversion factors.The dried samples were ground and used to measure the plant C and N contents through the use of an element analyzer(Carlo Erba 1106 Italy).
In each plot, five soil cores(5 cm in diameter)were randomly collected at 0–20,20–40,and 40–100 cm depth and thoroughly mixed into a homogenized sample.After removing the plant roots,fauna,and debris by hand,the soil was air dried at room temperature around 20°C,and then ground and passed through a sieve(2 mm)for measurement of C and N contents.C content was determined using a mixture of K2Cr2O7and H2SO4wet oxidation method from Walkley and Black(1934).N content was measured using the Kjeldahl method(Bremner and Mulvaney 1982).After excluding recognizable soil surface litter,stainless cutting rings(5 cm in diameter)were used to sample five replicated 100 cm3of soil at each layer at the same depth intervals of 0–20 cm,20–40 cm,and 40–100 cm layers.The ring soil samples were scraped out and roots were manually removed.The soils were dried at 105°C to constant weight in order to calculate bulk density.
Data calculation and analysis
Indices used to quantify plant diversity were species richness,Simpson’s diversity(Simpson 1949),Shannon–Wiener diversity(Shannon and Weaver 1949),and Pielou’s evenness(Pielou 1969),as determined by the following equations:
Species number as the richness index (S),

Table 1 Allometric equations used to calculate the various tree component biomasses of black locust on the Loess Plateau,China
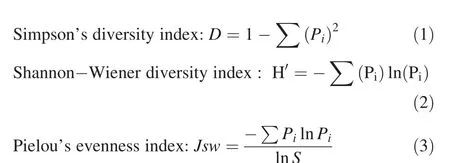
where,Sis the number of species,Piis the proportion of individuals or the abundance of theith species expressed as a proportion of the total in the community,andlnis log base-e.
Total ecosystem C and N were computed based on the combination of the tree,herb,litter,and soil pools.Leaf,branch,stem,and root C and N contents were multiplied by the tree component biomasses from the derived allometric equations(Table 1)to partition C and N stocks among the tree components,and summed for each tree and site level to calculate stand level tree biomass C and N stocks.For herbs and litter,C and N contents were multiplied by the component mass at plot level to calculate stand level C and N stocks.The stocks of soil C and N for each soil layer were based on the depth of the soil layer,its bulk density(Table S1),and content of these elements,and then the values for all studied layers(0–20 cm,20–40 cm,and 40–100 cm)were summed to estimate the total stock of soil C and N to a depth of 100 cm.
All statistical analyses were performed using SPSS statistical software(version 20.0,SPSS Inc,Chicago,IL,USA)and the signi ficance level was set at α=0.05.All comparisons were performed using ANOVA followed by multiple comparisons(LSD tests).
Results
Stand characteristics and plant diversity
Throughout stand development,there were signi ficant differences in stand structure and plant diversity.Stand density slightly decreased from the initial planting density of about 3300 trees ha-1to 2791 trees ha-1at the 9-year old stand.Density then declined to 1916,1325,and 1300 trees ha-1at the 17-,30-,and 37-year old stands,respectively,but tree density did not differ by the three stands age.Tree DBH and H among the 9-,17-,and 30-year old stands were 5.19 and 5.06,7.21 and 5.92,and 9.65 cm and 7.19 m,respectively,and there were no signi ficant differences both in Tree DBH and H among these stands.However,there was a signi ficant increase from the 30-year old stand to a DBH of 14.05 cm and an H of 9.14 m at the 37-year old stand.
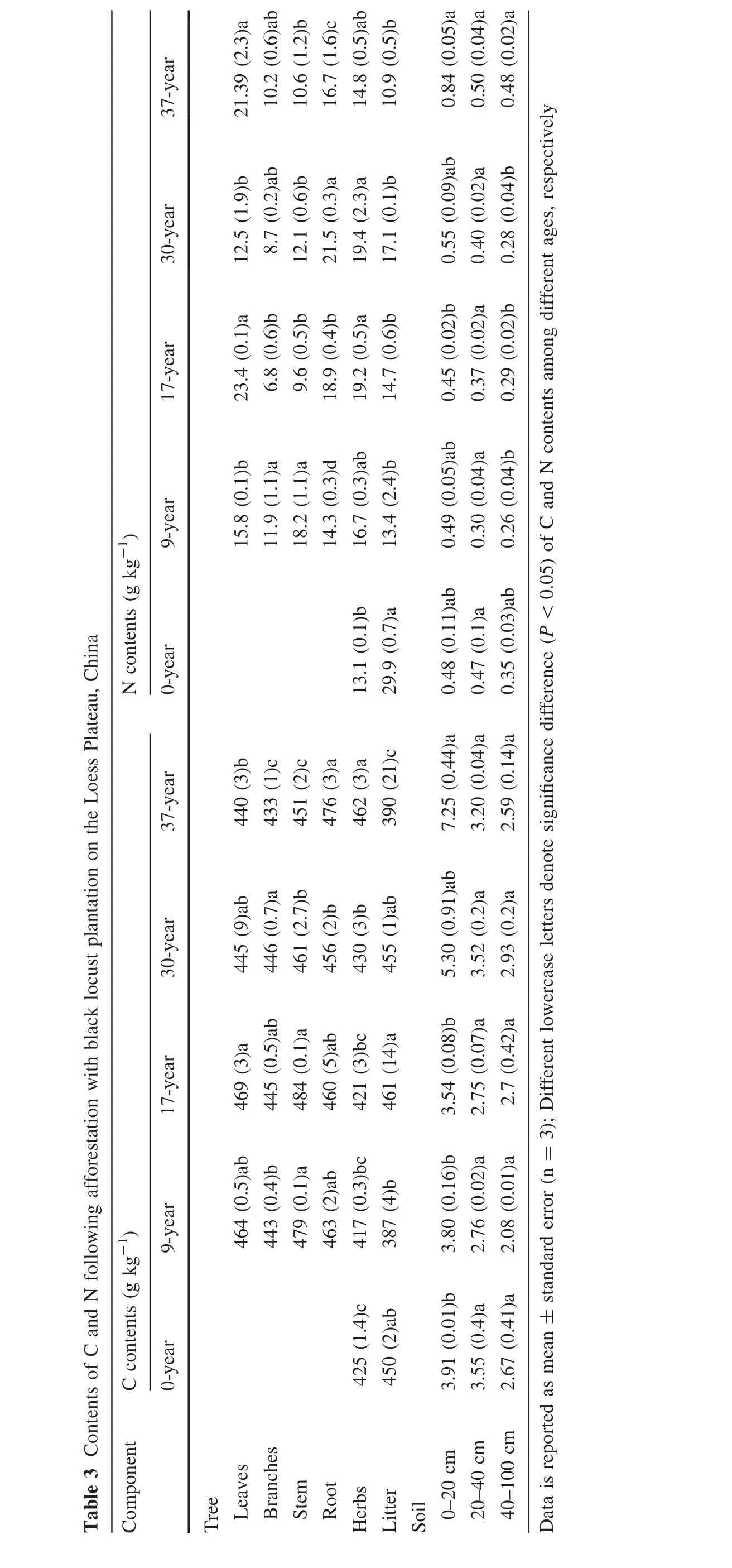
Among the herbaceous plants,the dominant species having the highest mean coverage wasParthenocisus tricuspidataat the pre-afforestation sites,followed bySetaria viridisat the 9-year stand andArtemisia sacrorumat the 17-year stand.At the more mature stages of 30-and 37-years old,the dominant colonizing species wasStipa bungeanaTrin.Species richness,diversity,and evenness were greatest at the pre-afforestation sites(Table 2).These parameters reached their lowest values at the early afforestation period(9-year old stand)and then gradually increased as stands aged(Table 2).
Tree,herb,and litter biomass C and N storages
C and N contents of tree tissues,herbs,and litter showed signi ficant age-dependent changes(Table 3).C content of leaves,branches,and stems decreased significantly during stand development.C and N contents in other tree tissues fluctuated during stand development.Total tree biomass C and N storage,as well as storage in individual tree tissues,slightly decreased from 9-to 17-year old stands,and then significantly increased from 17-to 37-year old stands(Figs.1,2).Stems and roots were the major tree C and N pools,each accounting on average for about 40%of storage.The proportion of stem biomass C and N storage to total tree biomass C and N storage increased during stand maturation,from 37%at the 9-year old stand to 42%at the 37-year old stand.In contrast,the proportion of root C and N stocks to the total tree biomass C and N storage decreased during stand aging from 45%at 9-year old stands to 39%at 37-year old stands.
Relative to those in the pre-afforestation sites,herb C and N storage decreased to minimum values at 9-years,and then significantlyincreasedasstandsaged.ThegreatestCstorage in herbs was 1.24 t ha-1at 37 years,165%of the value recorded at pre-afforestation sites.Herb N stocks signi ficantly increased from 0.02 t ha-1at 0-year stand to 0.04 t ha-1at17-yearstands,whichweredoublethoseatthe pre-afforestation sites.There were no signi ficant increases from the 17-to the 37-year old ages(Fig.1).However,the minimum C and N storage values in litter were recorded at pre-afforestation sites,where they significantly increased to their greatest values in 9-year old stands.C and N storage in litter showed a decrease characterized by fluctuations from the 9-to 37-year old stands(Fig.1).
Soil C and N contents and storage
Contents of C and N changed significantly during stand development,especially in the surface soil layer.The C and N contents of the 0–20 cm depth soil,as well as the N content at 40–100 cm depth,continually increased until the 37-year old stage(P<0.01 for all;Table 3).C and N storage varied by soil depth after afforestation.Compared with pre-afforestation sites,C storage at 0–20 cm depth remained at constant levels until 30 years of age,and then significantly increased from 30 to 37 years old(26.02 t ha-1),reaching 152%of the value recorded at preafforestation sites.Although C storage at the 20–40 cm depth increased from 9 to 37 years old,there were no clear enhancements in C sequestration compared to the pre-afforestation sites.Soil C storage at 40–100 and 0–100 cm depths initially decreased until reaching 17 years of age and then significantly increased to levels higher than those in the pre-afforestation sites;there was an increase of 25%at the 40–100 cm depth and 29%at the 0–100 cm depth(Figs.1,2).
Compared to the pre-afforestation sites,soil N storage at 0–20 cm depth remained constant during stand development.N storage at 20–40 cm,40–100 cm,and 0–100 cm depths all initially decreased in the afforested stands at 9 years of age but differed thereafter.N storage increased at 20–40 cm but remained 23%below the pre-afforestation levels after 37 years.At the 40–100 and 0–100 cm depths,N storage increased to pre-afforestation levels between years 9 and 37(Figs.1,2).
Ecosystem C and N storages
Total ecosystem C in the pre-afforestation sites was 79.71 t ha-1,and 98%of which was stored in the soil.After planting black locust,total ecosystem C decreased from 76.23 t ha-1in the 9-year old stand to 68.87 t ha-1in the 17-year old stand.Thereafter,total ecosystem C significantly increased to 93.26 t ha-1in the 30-year old stand and 138.94 t ha-1in the 37-year old stand,accounting for 117 and 174%of the pre-afforestation levels,respectively(Fig.2).Total ecosystem N storage remained constant until stands reached 30 years of age and then significantly increased from 30 to 37 years old.C and N pools of vegetation(including all tree tissues,herbs,and litter)after afforestation increased significantly.C and N pools of herbs and litter showed age-dependent changes,and accounted for only less than 1%of total vegetation C and N storage(Figs.1,2).Thus,the increased C and N pools in vegetation were mainly a result of increased tree DBH and height.The C and N in tree biomass contributed an average of 17.3%and 7.8%of the total ecosystem,respectively,and increased with stand development.The C stocks in the soil(101.4 t ha-1)were more than twice that in the vegetation(37.5 t ha-1)and the N stocks(12.9 t ha-1)were more than seven times that in the vegetation(1.8 t ha-1).Thus,more C and N were stored in the soil than in the vegetation and the relative contribution of soil to the total ecosystem C and N decreased with stand development(Fig.2).
Discussion
Tree biomass C and N of black locust plantations
Afforestation can help sequester atmospheric CO2through increased C and N storage in soil and vegetation.Forest structure,species composition,and primary productivity change as vegetation grows,and this can directly support the process of C and N accumulation in vegetation biomass(Aryal et al.2014;Drake et al.2011).In this study,we recorded stand density of 2791 trees ha-1in 9-year old stands that declined to 1300 trees ha-1in 37-year old stands while tree DBH of 5.19 cm and H of 5.06 m in 9-year old stands increased to 14.05 cm and 9.14 m in 37-year old stands.As a consequence,total tree biomass Cand N storage,as well as storage in individual tree tissues,increased with stand age.However,a slow growth was recorded from ages 9–17 years(Fig.1).These results are consistent with previous studies which found that tree biomass C and N storage declined in young plantations and then increased as stands aged(Cheng et al.2014;Deng et al.2014b;Hu et al.2009;Li et al.2013a).Shen and Zhang(2014)reported that black locust plantations reach their highest annual net tree biomass carbon storage after 5 years of afforestation and then maintain this rate with continuing maturation.Signi ficant decrease in tree biomass C in older stands(53 years)of black locust plantations was reported for the Loess Plateau due to low soil moisture,which resulted in wilting of the top of the crown and caused the death of individual trees(Li and Liu 2014).The higher C pool in the stems and N pools in roots were due to the relatively higher contents of C in stems and contents of N in roots,as well as the higher proportion of stem and root biomass(Table 3;Fig.1).This was consistent with results from previous studies(Li et al.2013a;Li and Liu 2014;Mei et al.2009;Wang et al.2007)which reported that most tree biomass C and N stocks(about 40%)are sequestered in stems and roots.Hu et al.(2009)reported that stems(accounting for 41.1%of C and 45.9%of N)and roots(accounting for 27.9%of C and 23.0%of N)were major biomass C stocks in 5-and 10-year-old poplar plantations.

Table 2 Species diversity indices of herbs following afforestation with black locust plantation on the Loess Plateau,China
?
Understory biomass C and N of black locust plantations
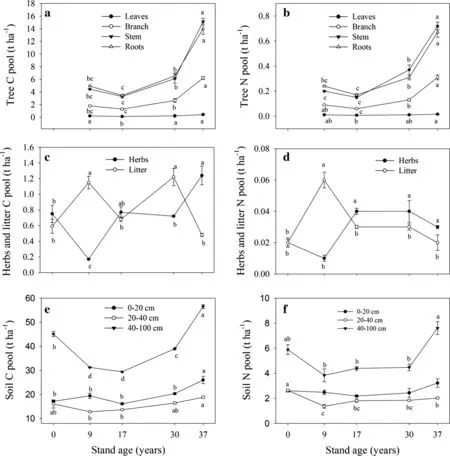
Fig.1 Dynamics of C(left)and N(right)storage of tree tissues(a,b);herb and litter(c,d);and different soil layers(e,f)following afforestation with black locust plantation on the Loess Plateau,China.Error bars indicate the standard error of the mean value.Bars having the same lowercase letter above them indicate no signi ficant difference between them(P<0.05)
The accumulation and decomposition of understory vegetation and litter play important roles in increasing soil C and N pools(Forrester et al.2013;Lee et al.2011).However,the storages in understory vegetation in our study comprised less than 1%of total ecosystem C and N storage,con firming previous results of studies of plantations and secondary forests(Li and Liu 2014;Yang et al.2014;Zhang et al.2014).In our study,C and N storage increased in herbs but decreased in litter during stand development(Fig.1).The decrease in litter C and N following afforestation in this study contrasts with the increases in litter C and N storage that come with stand age and the age-independent mechanisms observed in temperate and tropical plantations and secondary forests(Aryal et al.2014;Li and Liu 2014;Yang et al.2014).Litter is highly susceptible to disturbances,variation in stand treatment,litter input,and decomposition rate,all of which might account for the inconsistent relationships between stand age and C and N in the litter(Li et al.2011;Peichl and Arain 2007;Zhang et al.2014).We recorded higher plant diversity in the herb layers of the pre-afforestation sites.Plant diversity at these sites was lowest at 9 years after plantation mainly due to human destruction of herbs during afforestation,Diversity subsequently increased as stands matured.Similar results were reported forPinus tabulaeformisplantations at varied densities when compared to stands without afforested trees(Chen and Cao 2014).Understory vegetation in the black locust plantation of the Loess Plateau was poor,almost exposed,and barely covered by herbaceous plants.Therefore,the limited survival of black locust shrubs should not be considered a limiting factor in assessing the ecological success of restoration on the Chinese Loess Plateau.Instead,natural restoration has been suggested as an appropriate method of enhancing C accumulation and ecological restoration on the Loess Plateau(Jiao et al.2012;Song et al.2014).
Soil C and N of black locust plantations
Many factors have been considered in the study of the dynamics of soil C and N storage after afforestation,including previous land use practices,forest type,tree species,soil properties,and climate conditions(Lu et al.2013;Paul et al.2002;Pregitzer and Euskirchen 2004;Pietrzykowski and Daniels 2014;Zhang et al.2014).C and N contents are expected to be higher in the surface soil than in the deeper layers,and their contents in younger stands tend to be lower than in older stands(Jobba´gy and Jackson 2001).We found that soil C and N contents at 0–20 cm depth increased gradually during stand development,whereas their contents at subsoil depths,with the exception of N content at the 40–100 cm depth,were not related to stand age(Table 3).This result is consistent with results from a study by(Zhang et al.2014)that was conducted inCryclobalanopsisplantations in southwest China.Studies have shown that C contents in subsoils decrease with plantation age(Cote et al.2000;Sartori et al.2007).
Soil C and N storage across the studied chronosequence were characterized by initial declines from pre-afforestation stands,recovery at 30 years of age to pre-afforestation levels,and signi ficant increases in levels in 37-year old stands compared to pre-afforestation sites(Figs.1,2).Similar patterns have been reported in other studies(Guo and Gifford 2002;Liao et al.2010).For example,soil C decreased by 38.9%to minimum levels 16 years after the conversion of natural forests to plantations,but the increased soil C in the mature stage was still 30%lower than the pre-harvest levels(Chen et al.2013).
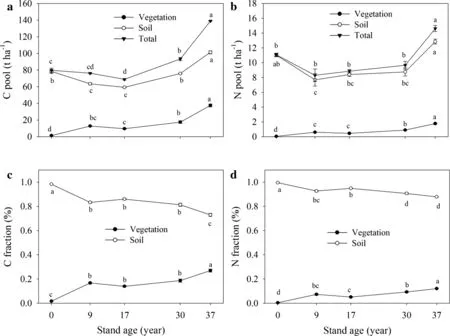
Fig.2 Dynamics of C(a)and N(b)storage in vegetation(including tree,herb and litter),soil(0–100 cm)and total ecosystem,and the relative share of C(c)and N(d)storage in vegetation and soil to the total ecosystem.Error bars indicate the standard error of the mean value.Bars having the same lowercase letter above them indicate no signi ficant difference between them(P<0.05)
Intensive management practices,such as clear-cutting,slash burning,and site preparation,and the interactions with local climatic,topographical,and edaphic conditions can cause high initial soil C and N losses following landuse conversion.The retention of organic matter in soils provides a mechanism that alleviates soil C loss or even increases soil C sequestration,thus soil disturbance weakens the physical protection of organic matter and soil particles and simultaneously increases soil temperature,which can accelerate soil C loss(Chen et al.2013;Oades 1988).However,in our study,the patterns in soil C and N storage varied by soil depth across the studied chronosequence(Fig.1),indicating that deep soils were the ones facing initial C and N loss during the conversion,and the recovery process during stand development was longer in deeper soil layers than in surface soils.The higher C loss in deeper soils and a delay in replenishing the C pool during stand development was also observed in Chinese fir plantations in south China,suggesting that the subsoils contain large amounts of labile C that is sensitive to plantation practices due to alteration of the soil’s thermal environment,which includes the mechanical mixing and redistribution of C and nutrients in the pro file(Chen et al.2013).
Ecosystem C and N of black locust plantations
Soil is considered to be the main sink for C and N,although the contribution of soil C and N to total ecosystem C and N storage decreased due to increases in biomass C and N accumulation as stands age,representing 73 and 87%of total ecosystem C and N storage in 37-year old stands,respectively(Fig.2).The similar results in afforestation of Mongolian pine in China’s Horqin Sandy Land indicates that plants play the most important role in ecosystem C sequestration,whereas soil plays the most important role in ecosystem N sequestration(Li et al.2013b).However,to achieve the initial aim of increasing ecosystem C and N storage following conversion of farmlands to plantations,a stand age of 30 years old needs to be reached(Fig.2).The net increase ecosystem C storage is reached at around 16 years for Chinese fir plantations that have been converted from natural broadleaved forests in south China(Chen et al.2013).With continual succession,ecosystem C and N storage increased significantly from 30-to 37-year old stands(Fig.2).Therefore,it is essential to understand the ecosystem equilibrium interactions between C and N storage in the vegetation and soil at over-mature phases in orderto fully understand the long-term effects of afforestation of farmlands on C sequestration.
Conclusion
The large plantation areas on the hilly Loess Plateau effectively enhance C sequestration within the forest ecosystem.Tree biomass C and N storage in the black locust plantations slightly increased from 9 to 30 years old,and then significantly increased from 30 to 37 years old.Thus,the increased C and N pools in the vegetation were mainly a result of the increased tree DBH and height and the biomass of tree components can be better predicted from allometric equations using DBH and height as independent variables.Afforestation on farmland caused soil C and N loss during the early stages of stand development,especially in the subsoil(40–100 cm depth).The relative contribution to the total ecosystem C and N pools increased in trees and decreased in soil during the observed period.Therefore,in carbon management or systems management,we should pay more attention to the role of soil,especially in the early stages of afforestation.Compared to storage in pre-afforestation sites,C and N storage in the soil(0–100 cm depth)recovered to pre-afforestation levels at around 30 years of age.C and N contents in subsoil(20–40 cm depth)remained lower than those of the preafforestation sites during the entire study period.Thus,the dynamics of C and N stocks in the subsoil and the increase in biomass throughout stand maturation should be considered in long-term assessments of ecosystem C and N stocks following afforestation of farmlands on the Loess Plateau.
AcknowledgementsThis research was supported by the National Nature Science Foundation of China(No.41201088,41371506 and 41601058).The authors would like to acknowledge the contributions made by Christian J.Rivera(Princeton University,USA)regarding the English language revision of the manuscript in the early work.In addition,the authors wish to thank Journal of Forestry Research editors and reviewers for their constructive suggestions and language polish to improve the quality of this article.
References
Arevalo CBM,Bhatti JS,Chang SX,Sidders D(2009)Ecosystem carbon stocks and distribution under different land-uses in north central Alberta,Canada.For Ecol Manag 257:1776–1785
Aryal DR,De Jong BHJ,Ochoa-Gaona S,Esparza-Olguin L,Mendoza-Vega J(2014)Carbon stocks and changes in tropical secondary forests of southern Mexico.Agric Ecosyst Environ 195:220–230
Berthrong ST,Jobbagy EG,Jackson RB(2009)A global metaanalysis of soil exchangeable cations,pH,carbon,and nitrogen with afforestation.Ecol Appl 19:2228–2241
Bradford JB,Kastendick DN(2010)Age-related patterns of forest complexity and carbon storage in pine and aspen-birch ecosystems of northern Minnesota,USA.Can J For 40:401–409
Bremner JM,Mulvaney CS(1982)Nitrogen-total.In:Page AL,Miller RH,Keeney DR(eds)Methods of soil analysis.Part 2.Chemical and microbiological properties.American Society of Agronomy,Madison,pp 595–624
Chang RY,Fu BJ,Liu GH,Liu SG(2011)Soil carbon sequestration potential for ‘‘Grain for Green’’project in Loess Plateau,China.Environ Manag 48:1158–1172
Chen Y,Cao Y(2014)Response of tree regeneration and understory plant species diversity to stand density in maturePinus tabulaeformisplantations in the hilly area of the Loess Plateau,China.Ecol Eng 73:238–245
Chen GS,Yang ZJ,Gao R,Xie JS,Guo JF,Huang ZQ,Yang YS(2013)Carbon storage in a chronosequence of Chinese fir plantations in southern China.For Ecol Manag 300:68–76
Cheng XQ,Han HR,Kang FF,Song YL,Liu K(2014)Variation in biomass and carbon storage by stand age in pine(Pinus tabulaeformis)planted ecosystem in Mt.Taiyue,Shanxi,China.J Plant Interact 9:521–528
Cote L,Brown S,Pare D,Fyles J,Bauhus J(2000)Dynamics of carbon acid nitrogen mineralization in relation to stand type,stand age and soil texture in the boreal mixedwood.Soil Biol Biochem 32:1079–1090
Davidson EA,Janssens IA(2006)Temperature sensitivity of soil carbon decomposition and feedbacks to climate change.Nature 440:165–173
Deng L,Shangguan ZP,Sweeney S(2013)Changes in soil carbon and nitrogen following land abandonment of farmland on the Loess Plateau,China.PLoS ONE 8:e71923
Deng L,Liu GB,Shangguan ZP(2014a)Land-use conversion and changing soil carbon stocks in China’s ‘Grain-for-Green’Program:a synthesis.Glob Change Biol 20:3544–3556
Deng L,Shangguan ZP,Sweeney S(2014b) ‘‘Grain for Green’’driven land use change and carbon sequestration on the Loess Plateau,China.Sci Rep 4:7039
Deng L,Wang KB,Li JP,Shangguan ZP,Sweeney S(2014c)Carbon Storage Dynamics in Alfalfa(Medicago sativa)Fields in the Hilly-Gully Region of the Loess Plateau,China.Clean Soil Air Water 42:1253–1262
Dixon RK,Brown S,Houghton RA,Solomon AM,Trexler MC,Wisniewski J(1994)Carbon pools and flux of global forest ecosystems.Science 263:185–190
Drake JE,Davis SC,Raetz LM,DeLucia EH(2011)Mechanisms of age-related changes in forest production:the in fluence of physiological and successional changes.Glob Change Biol 17:1522–1535
Fang JY,Chen AP,Peng CH,Zhao SQ,Ci L(2001)Changes in forest biomass carbon storage in China between 1949 and 1998.Science 292:2320–2322
FAO–UNESCO(1974)Soil map of the world(1:5,000,000).Food and Agricultural Organisation of the United Nations,UNECO,Paris
Feng X,Fu B,Lu N,Zeng Y,Wu B(2013)How ecological restoration alters ecosystem services:an analysis of carbon sequestration in China’s Loess Plateau.Sci Rep 3:2846
Forrester DI,Pares A,O’Hara C,Khanna PK,Bauhus J(2013)Soil organic carbon is increased in mixed-species plantations of eucalyptus and nitrogen- fixing.Acacia Ecosyst 16:123–132
Guo LB,Gifford RM(2002)Soil carbon stocks and land use change:a meta analysis.Glob Change Biol 8:345–360
Hu Y,Zeng D,Jiang T(2009)Effects of afforested poplar plantations on the stock and distribution of C,N,P at Keerqin Sandy Lands.Acta Ecol Sin 29:4206–4214
Jackson RB,Schlesinger WH(2004)Curbing the U.S.carbon de ficit.Proc Natl Acad Sci USA 101:15827–15829
Jiao JY,Zhang ZG,Bai WJ,Jia YF,Wang N(2012)Assessing the ecological success of restoration by afforestation on the Chinese Loess Plateau.Restor Ecol 20:240–249
Jobba´gy E,Jackson R(2001)The distribution of soil nutrients with depth:global patterns and the imprint of plants.Biogeochemistry 53:51–77
Jo´zefowska A,Pietrzykowski M,Wos´B,Cajthaml T,Frouz J(2017)The effects of tree species and substrate on carbon sequestration and chemical and biological properties in reforested post-mining soils.Geoderma 292:9–16
Khanna PK(1997)Comparison of growth and nutrition of young monocultures and mixed stands ofEucalyptus globulusandAcacia mearnsii.For Ecol Manag 94:105–113
Laganiere J,Angers DA,Pare D(2010)Carbon accumulation in agricultural soils after afforestation:a meta-analysis.Glob Change Biol 16:439–453
Lal R(2004)Soil carbon sequestration to mitigate climate change.Geoderma 123:1–22
Lee YC,Nam JM,Kim JG(2011)The in fluence of black locust(Robinia pseudoacacia) flower and leaf fall on soil phosphate.Plant Soil 341:269–277
Li T,Liu G(2014)Age-related changes in carbon accumulation and allocation in plants and soil of a black locust forest on the Loess Plateau.Chin Geogr Sci 24:414–422
Li X,Yi MJ,Son Y,Park PS,Lee KH,Son YM,Kim RH,Jeong MJ(2011)Biomass and carbon storage in an age-sequence of Korean Pine(Pinus koraiensis)plantation forests in Central Korea.J Plant Biol 54:33–42
Li D,Niu S,Luo Y(2012)Global patterns of the dynamics of soil carbon and nitrogen stocks following afforestation:a metaanalysis.New Phytol 195:172–181
Li H,Li J,He YL,Li SJ,Liang ZS,Peng CH,Polle A,Luo Zh(2013a)Changes in carbon,nutrients and stoichiometric relations under different soil depths,plant tissues and ages in black locust plantations.Acta Physiol Plant 35:2951–2964
Li YQ,Brandle J,Awada T,Chen YP,Han JJ,Zhang FX,Luo YQ(2013b)Accumulation of carbon and nitrogen in the plant-soil system after afforestation of active sand dunes in China’s Horqin Sandy Land.Agric Ecosyst Environ 177:75–84
Liao CZ,Luo YQ,Fang CM,Li B(2010)Ecosystem carbon stock in fluenced by plantation practice:implications for planting forests as a measure of climate change mitigation.PLoS ONE 5:e10867
Liu ZP,Shao MA,Wang YQ(2011)Effect of environmental factors on regional soil organic carbon stocks across the Loess Plateau region,China.Agric Ecosyst Environ 142:184–194
Liu ZP,Shao MA,Wang YQ(2014)The contribution of China’s Grain to Green Program to carbon sequestration.Landsc Ecol 29:1675–1688
Lu N,Liski J,Chang RY,Akujarvi A,Wu X,Jin TT,Wang YF,Fu BJ(2013)Soil organic carbon dynamics of black locust plantations in the middle Loess Plateau area of China.Biogeosciences 10:7053–7063
Luo Y,Bo S,William SC,Jeffery SD(2004)Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide.Bioscience 54:731–739
Mao R,Zeng DH,Hu YL,Li LJ,Yang D(2010)Soil organic carbon and nitrogen stocks in an age-sequence of poplar stands planted on marginal agricultural land in Northeast China.Plant Soil 332:277–287
Mazurek R,Bejger R(2014)The role of black locust(Robinia pseudoacaciaL.)shelterbelts in the stabilization of carbon pools and humic substances in chernozem.Pol J Environ Stud 23:1263–1271
Mei L,Zhang Z,Gu J,Quan X,Yang L,Huang D(2009)Carbon and nitrogen storages and allocation in tree layers ofFraxinus mandshuricaandLarix gmeliniiplantations.Chin J Appl Ecol 20:1791–1796
Metz B,Davidson OR,Bosch PR,Dave R,Meyer LA(2007)Contribution of Working Group III to the fourth assessment report of the Intergovernmental Panel on Climate Change
Oades JM(1988)The retention of organic matter in soils.Biogeochemistry 5:35–70
Paul KI,Polglase PJ,Nyakuengama JG,Khanna PK(2002)Change in soil carbon following afforestation. For Ecol Manag 168:241–257
Peichl M,Arain MA(2007)Allometry and partitioning of above-and belowground tree biomass in an age-sequence of white pine forests.For Ecol Manag 253:68–80
Persson M,Moberg J,Ostwald M,Xu JT(2013)The Chinese Grain for Green Programme:assessing the carbon sequestered via land reform.J Environ Manag 126:142–146
Pielou EC(1969)An introduction to mathematical ecology.Wiley,New York
Pietrzykowski M,Daniels WL(2014)Estimation of carbon sequestration by pine(Pinus sylvestrisL.)ecosystems developed on reforested post-mining sites in Poland on differing mine soil substrates.Ecol Eng 73:209–218
Pietrzykowski M,Krzaklewski W(2007)Soil organic matter,C and N accumulation during natural succession and reclamation in an opencast sand quarry(southern Poland).Arch Agron Soil Sci 53:473–483
Post WM,Kwon KC(2000)Soil carbon sequestration and land-use change:processes and potential.Glob Change Biol 6:317–327
Pregitzer KS,Euskirchen ES(2004)Carbon cycling and storage in world forests:biome patterns related to forest age.Glob Change Biol 10:2052–2077
Qiu LP,Zhang XC,Cheng JM,Yin XQ(2010)Effects of black locust(Robinia pseudoacacia)on soil properties in the loessial gully region of the Loess Platea,Chinau.Plant Soil 332:207–217
Rastetter EB,Agren GI,Shaver GR(1997)Responses of N-limited ecosystems to increased CO2:a balanced-nutrition,coupledelement-cycles model.Ecol Appl 7:444–460
Ritter E(2007)Carbon,nitrogen and phosphorus in volcanic soils following afforestation with native birch(Betula pubescens)and introduced larch (Larixsibirica)in Iceland.PlantSoil 295:239–251
Sang PM,Lamb D,Bonner M,Schimdt S(2013)Carbon sequestration and soil fertility of tropical tree plantations and secondary forest established on degraded land.Plant Soil 362:187–200
Sartori F,Lal R,Ebinger MH,Eaton JA(2007)Changes in soil carbon and nutrient pools along a chronosequence of poplar plantations in the Columbia Plateau,Oregon,USA.Agric Ecosyst Environ 122:325–339
Shannon CE,Weaver W (1949)The mathematical theory of communication.University of Illinois Press,Urbana
Shen JP,Zhang WH(2014)Characteristics of carbon storage and sequestration ofRobinia pseudoacaciaforest land converted by farmland in the Hilly Loess Plateau Region.Acta Ecol Sin 34:2746–2754
Shi H,Shao MA(2000)Soil and water loss from the Loess Plateau in China.J Arid Environ 45:9–20
Simpson EH(1949)Measurement of diversity.Nature 163:688
Song X,Peng C,Zhou G,Jiang H,Wang W(2014)Chinese Grain for Green Program led to highly increased soil organic carbon levels:a meta-analysis.Sci Rep 4:4460
Walkley A,Black IA(1934)An examination of the Degtjareff method for determining soil organic matter,and a proposed modi fication of the chromic acid titration method.Soil Sci 34:29–38
Wang H,Huang Y,Feng Z,Wang S(2007)C and N stocks under three plantation forest ecosystems of Chinese fir,Michelia macclureiand their mixture.Front For Chin 2:251–259
Wang B,Liu GB,Xue S(2012)Effect of black locust(Robinia pseudoacacia)on soil chemical and microbiological properties in the eroded hilly area of China’s Loess Plateau.Environ Earth Sci 65:597–607
Yang YH,Luo YQ,Finzi AC(2011)Carbon and nitrogen dynamics during forest stand development:a global synthesis.New Phytol 190:977–989
Yang Y,Wang GX,Shen HH,Yang Y,Cui HJ,Liu Q(2014)Dynamics of carbon and nitrogen accumulation and C:N stoichiometry in a deciduous broadleaf forest of deglaciated terrain in the eastern Tibetan Plateau.For Ecol Manag 312:10–18
Zhang QJ,Fu BJ,Chen LD,Zhao WW,Yang QK,Liu GB,Gulinck H(2004)Dynamics and driving factors of agricultural landscape in the semiarid hilly area of the Loess Plateau,China.Agric Ecosyst Environ 103:535–543
Zhang F,Zhang SL,Cheng ZJ,Zhao HY(2007)Time structure and dynamics of the insect communities in bush vegetation restoration areas of Zhifanggou watershed in Loess hilly region.Acta Ecol Sin 27:4555–4562
Zhang H,Song TQ,Wang KL,Du H,Yue YM,Wang GX,Zeng FP(2014)Biomass and carbon storage in an age-sequence ofCyclobalanopsis glaucaplantations in southwest China.Ecol Eng 73:184–191
Zhou GY,Liu SG,Li Z,Zhang DQ,Tang XL,Zhou CY,Yan JH,Mo JM(2006)Old-growth forests can accumulate carbon in soils.Science 314:1417
 Journal of Forestry Research2018年3期
Journal of Forestry Research2018年3期
- Journal of Forestry Research的其它文章
- In vitro propagation of conifers using mature shoots
- ‘Relationships between relationships’in forest stands:intercepts and exponents analyses
- Effects of application date and rate of foliar-applied glyphosate on pine seedlings in Turkey
- Assaying the allelopathic effects of Eucalyptus camaldulensis in a nursery bed incorporated with leaf litter
- Effects of soil compaction on growth variables in Cappadocian maple(Acer cappadocicum)seedlings
- Variation and selection analysis of Pinus koraiensis clones in northeast China