
ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴对脂肪组织糖脂代谢的影响
2018-05-16 05:27马池发史婷婷袁明霞
基础医学与临床 2018年5期
马池发,史婷婷,袁明霞
(首都医科大学附属北京同仁医院 内分泌科 糖尿病防治研究北京市重点实验室, 北京 100730)
肾素-血管紧张素系统(renin angiotensin system,RAS)的经典作用是调节血压和维持水、电解质平衡。RAS不仅存在于循环系统,还存在于局部组织。RAS存在多条生物活性轴,其中备受关注的是血管紧张素转化酶(ACE)-血管紧张素Ⅱ(Ang Ⅱ)-AT1轴和血管紧张素转化酶2(ACE2)-血管紧张素(1- 7)[Ang(1- 7)]-Mas轴,后者在一定程度上拮抗前者的作用, 近年来越来越多研究证明它们在糖脂代谢、胰岛素抵抗及糖尿病并发症发挥重要作用[1- 6]。课题组前期研究证实,ACE2改善胰岛β细胞线粒体功能,促进胰岛素分泌[7];ACE2/Ang(1- 7)/Mas轴可以激活AKT信号通路,缓解肝脏脂肪变性[8];Ang(1- 7)通过Mas受体抑制脂肪细胞氧化应激,增加葡萄糖摄取[9]。这些结果表明ACE2/Ang(1- 7)/Mas轴可能起到有利的代谢效应。脂肪组织是糖脂代谢的重要器官,RAS在脂肪组织糖脂代谢的作用近年成为研究的热点[10- 11]。本文就RAS中ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴在脂肪组织糖脂代谢方面的作用进行综述,为研究糖脂代谢提供新思路。
1 ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴生物学特性
1.1 ACE/Ang Ⅱ/AT1轴
血管紧张素原在肾素作用下转变为血管紧张素Ⅰ(angiotensin Ⅰ, Ang Ⅰ),后者在血管紧张素转化酶(angiotensin-converting-enzyme, ACE)作用下转变为血管紧张素Ⅱ(Ang Ⅱ)(图1)。Ang Ⅱ是RAS中重要活性肽,是目前所知收缩血管最强的物质之一。Ang Ⅱ主要通过G蛋白偶联受体AT1和AT2受体发挥作用,在心血管、肾脏、骨骼肌、肝脏和脂肪等器官、组织扮演的角色不尽相同。
1.2 ACE2/Ang(1- 7)/Mas轴
血管紧张素转化酶2(ACE2)是2000年被克隆的人类ACE的第一个同源基因。ACE2可催化Ang Ⅱ转变为血管紧张素(1- 7)[Ang(1- 7)],也可催化Ang Ⅰ转变为血管紧张素(1- 9)[Ang(1- 9)],后者可经过ACE或中性肽链内切酶(NEP)作用下转变为Ang(1- 7),Ang(1- 7)也可以直接由Ang Ⅰ在NEP作用下生成,Ang(1- 7)通过Mas受体发挥作用(图1)。ACE2/Ang(1- 7)/Mas轴在心血管、肾脏、脂肪组织等器官可拮抗ACE/Ang Ⅱ/AT1 的作用。
1.3 ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴的联系
ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴是对立、统一的,其组成成分存在一定的联系,两条轴均可作用在机体器官参与糖脂代谢调节,并且和肥胖及代谢综合征存在潜在的联系,多数研究认为后者可拮抗前者不利的代谢效应,进而对机体产生保护效应。在胰腺中,Ang Ⅱ引起胰岛β细胞功能障碍[12],ACE2/Ang(1- 7)减少高脂饮食鼠的胰岛β细胞去分化[13],ACE2促进胰岛细胞分泌胰岛素[7];在肝脏中,通过运动有助于ACE2/Ang(1- 7)/Mas轴拮抗ACE/Ang Ⅱ/AT1轴的作用,减少代谢紊乱和避免非酒精性脂肪肝的发生[14],Ang(1- 7)抑制肝脏脂肪合成[15],ACE2/Ang(1- 7)/Mas轴缓解肝脏脂肪变性[8];在骨骼肌中,常规剂量的Ang Ⅱ引起骨骼肌胰岛素抵抗,高剂量的Ang Ⅱ反而使胰岛素抵抗趋于正常,可能与ACE2及Mas蛋白表达增加相关[16]。此外,在骨代谢方面,ACE2/Ang(1- 7)/Mas轴可以拮抗ACE/Ang Ⅱ/AT1轴对骨代谢的不利作用,进而改善骨代谢[17]。
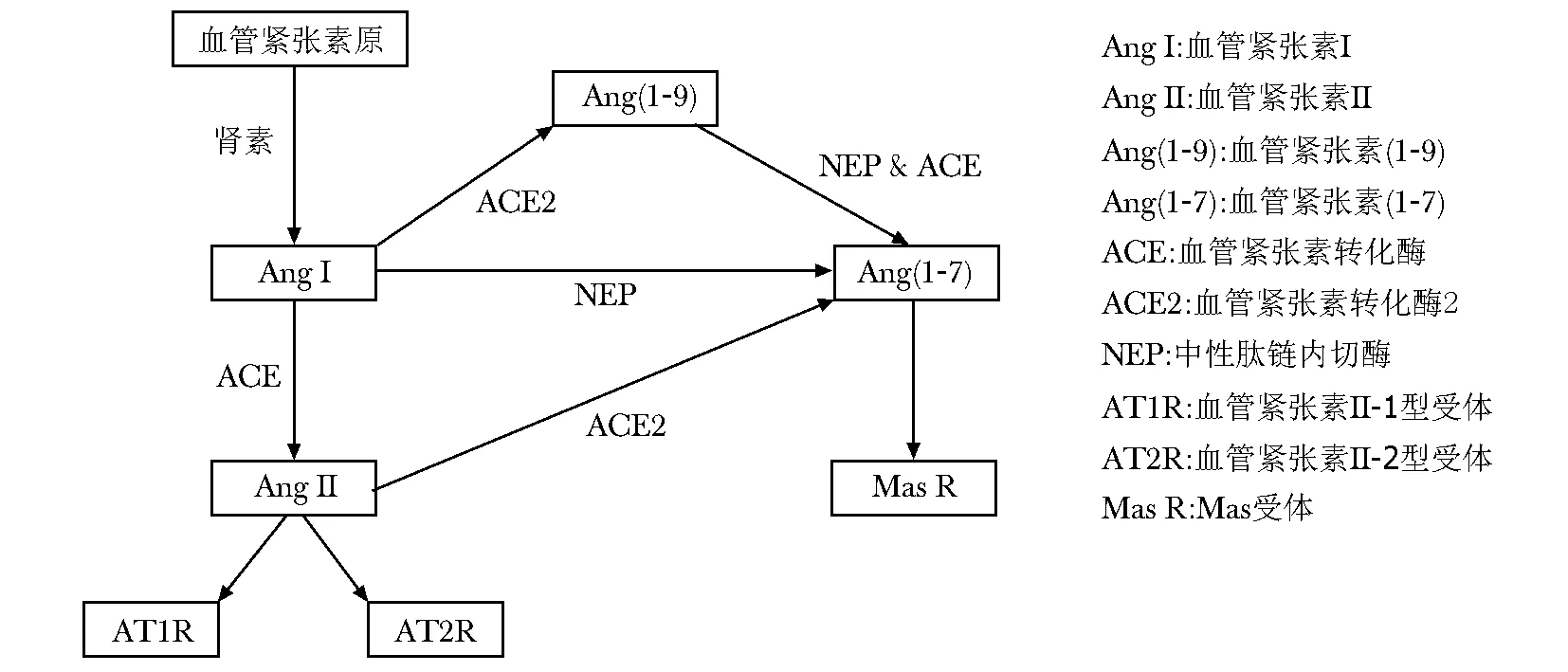
图1 ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴Fig 1 ACE/Ang Ⅱ/AT1 axis and ACE2/Ang(1- 7)/Mas axis
2 ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴与脂肪组织糖脂代谢的关系
2.1 脂肪组织的内分泌功能
哺乳动物的脂肪组织主要分为白色脂肪和棕色脂肪两种,白色脂肪不仅是能量储存库,还是重要的内分泌器官,通过分泌脂肪因子在机体的代谢中发挥重要的作用。棕色脂肪主要功能是非寒战产热。脂肪因子(adipokines)是脂肪组织分泌的各种激素和细胞因子,包括瘦素(leptin)、脂联素(adiponectin)、白介素- 6(interleukin- 6,IL- 6)、内脂素(visfatin)及RAS成员等。这些脂肪因子以自分泌、旁分泌或内分泌方式作用于脂肪组织或作用于机体其他组织,广泛参与调节和维持糖、脂代谢平衡,调控机体内分泌、能量代谢及炎性反应的发生。RAS成员在脂肪组织中表达,可调节其他脂肪因子的表达对代谢产生影响。RAS过度激活与人类及其他动物的肥胖相关。
2.2 ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴对脂肪组织糖代谢的影响
ACE/Ang Ⅱ/AT1轴可以引起脂肪组织糖代谢异常。Ang Ⅱ抑制脂肪组织摄取葡萄糖[18],AT1受体阻断剂氯沙坦促进3T3-L1脂肪细胞摄取葡萄糖[19]。
ACE2/Ang(1- 7)/Mas轴可以改善脂肪组织糖代谢紊乱。TGR(A1- 7)3292鼠可通过产生Ang(1- 7)融合蛋白缓慢升高其血浆Ang(1- 7),研究表明来自TGR(A1- 7)3292鼠的脂肪细胞在胰岛素刺激下其葡萄糖摄取量明显增加[20]。Ang(1- 7)可通过Mas受体增加脂肪细胞摄取葡萄糖[9]。Mas基因敲除鼠脂肪细胞对胰岛素刺激的葡萄糖摄取下降[21]。
2.3 ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴对脂肪组织脂代谢的影响
2.3.1 ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴对脂肪细胞形态的影响:肥大的脂肪细胞中脂肪因子的表达和分泌是紊乱的,脂肪细胞的大小与代谢性疾病和脂肪组织机能紊乱密切相关。研究表明AT1受体阻滞剂能减小3T3-L1脂肪细胞体积[19],Ang(1- 7)同样能减小脂肪细胞的体积[22],ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴通过对脂肪细胞大小调节进而对代谢产生影响。
2.3.2 ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴对脂肪细胞分化的影响:在脂肪分化方面,ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴起拮抗作用(图2)。脂肪分化过程是由前体脂肪细胞、干细胞等分化形成载脂的、对胰岛素反应的成熟脂肪细胞的过程。脂肪的分化伴随着脂肪的合成,有时也使用脂肪合成的指标来反应脂肪分化的程度。Ang Ⅱ通过AT1受体抑制脂肪干细胞分化,通过AT2受体促进脂肪干细胞分化[23]。Ang(1- 7)/Mas拮抗Ang Ⅱ/AT1的作用促进脂肪分化[24]。过氧化物酶体增殖物激活受体γ(PPARγ)是一种核受体超家族成员基因,是调控脂肪组织中前体脂肪细胞分化的关键因子,并且在调节脂类代谢中发挥重要的作用,其磷酸化后可失去活性。Ang(1- 7)/Mas通过激活PI3K/AKT/PPARγ途径和抑制Ang Ⅱ/ATI激发的MAPK酶/ERK1,2途径促进脂肪细胞分化[24]。
2.3.3 ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴对脂肪合成的影响:体内脂肪的积累主要受脂肪合成和分解的影响。脊椎动物的能量主要是以三酰甘油的形式储存在脂肪细胞,而三酰甘油主要来自于脂肪酸的从头合成。脂肪酸合成酶(FAS)是脂肪酸合成的关键酶,它能催化乙酰辅酶A和丙二酰辅酶A在还原型辅酶Ⅱ(NADPH)存在的条件下生成长链脂肪酸。Ang Ⅱ 通过AT2受体促进3L3-L1和人类脂肪细胞三酰甘油合成[25](图2)。不同类型的RAS阻滞剂对脂肪细胞脂肪合成作用不同[26]。ACE2激动剂乙酰甘氨酸重氮氨苯脒(diminazene aceturate,DIZE)抑制小鼠脂肪组织脂肪合成[27],Ang(1- 7)也抑制小鼠脂肪组织脂肪合成[15]。但是有研究证实Mas基因敲除鼠脂肪组织的脂肪合成因子受抑制[28],腹部脂肪却明显增加[21],脂肪合成因子表达与脂肪量的多少存在矛盾,关于RAS对脂肪合成的作用仍在研究中。
2.3.4 ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴对脂肪分解的影响:脂肪分解的过程主要涉及三酰甘油分解形成脂肪酸和甘油的过程。研究证实,Ang Ⅱ通过AT1受体抑制脂肪细胞脂肪分解[29],Ang(1- 7)通过MasR/PI3K/eNOS途径促进脂肪细胞脂肪分解[30](图2)。
3 问题与展望
探讨ACE2/Ang(1- 7)/Mas轴相关作用的研究方法主要包括构建相应的生物模型,多采用基因敲除法使ACE2、Mas基因缺失, 病毒转染法使ACE2、 Mas基因过表达,或者使用药物Ang(1- 7)替代。新近研究表明抗锥虫药物DIZE是ACE2内源性激动剂,使内源性激动ACE2成为可能。DIZE可以提高小鼠ACE2的活性,增加体内Ang(1- 7) 水平,下调肾脏炎性反应及纤维化水平,从而延缓糖尿病肾病的进展[4]。使用ACE2的激动剂DIZE后,小鼠体质量、血胆固醇、血三酰甘油、附睾及腹膜后脂肪重量明显低于对照组,这与小鼠附睾脂肪组织ACE2表达升高及ACE表达下降相关,DIZE激动的小鼠附睾脂肪组织脂肪合成相关基因乙酰辅酶A羧化酶、脂肪酸合成酶mRNA水平均下降[27]。这些研究结果进一步证实激活ACE2/Ang(1- 7)/Mas轴可能改善脂代谢。
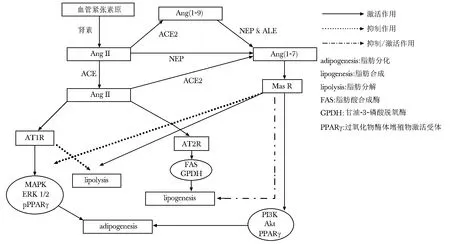
图2 ACE/AngⅡ/AT1轴和ACE2/Ang(1- 7)/Mas轴对脂肪组织脂代谢的影响
RAS对脂肪组织糖脂代谢发挥重要的作用。在糖代谢方面,多数观点认为两轴存在拮抗作用,ACE/Ang Ⅱ/AT1轴引起脂肪组织糖代谢紊乱,ACE2/Ang(1- 7)/Mas轴可改善糖代谢。而在脂代谢方面,ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴参与脂肪细胞分化、脂肪合成和脂肪分解过程,两条轴也存在部分拮抗调节作用,但是对脂肪组织脂肪合成的作用在不同背景下的实验结果仍存在矛盾,需要更多研究去证实。深入研究ACE/Ang Ⅱ/AT1轴和ACE2/Ang(1- 7)/Mas轴对脂肪组织糖脂代谢的作用环节,有可能为改善糖脂代谢及临床药物开发新的治疗靶点。
参考文献:
[1] Thorwald M, Rodriguez R, Lee A,etal. Angiotensin receptor blockade improves cardiac mitochondrial activity in response to an acute glucose load in obese insulin resistant rats[J]. Redox Biol, 2018, 14: 371- 378.
[2] Hussain SA, Utba RM, Assumaidaee AM. Effects of azilsartan, aliskiren or their combination on high fat diet-induced non-alcoholic liver disease model in rats[J]. Med Arch, 2017, 71: 251- 255.
[3] Souza-Mello V. Hepatic structural enhancement and insulin resistance amelioration due to AT1 receptor blockade[J]. World J Hepatol, 2017, 9: 74- 79.
[4] 王园园,曹新冉,杨旻,等. ACE2内源性激动剂DIZE对糖尿病肾病大鼠的保护作用[J]. 中国病理生理杂志,2017, 33: 469- 474.
[5] Patel VB, Mori J, Mclean BA,etal. ACE2 deficiency worsens epicardial adipose tissue inflammation and cardiac dysfunction in response to diet-induced obesity[J]. Diabetes, 2016, 65: 85- 95.
[6] Wu Y, Ma KL, Zhang Y,etal. Lipid disorder and intrahepatic renin-angiotensin system activation synergistically contribute to non-alcoholic fatty liver disease[J]. Liver Int, 2016, 36: 1525- 1534.
[7] Shi TT, Yang FY, Liu C,etal. Angiotensin-converting enzyme 2 regulates mitochondrial function in pancreatic beta-cells[J]. Biochem Biophys Res Commun, 2018, 495: 860- 866.
[8] Cao X, Yang F, Shi T,etal. Angiotensin-converting enzyme 2/angiotensin-(1- 7)/Mas axis activates Akt signaling to ameliorate hepatic steatosis[J]. Sci Rep, 2016, 6: 1- 11.
[9] Liu C, Lv XH, Li HX,etal. Angiotensin-(1- 7) suppresses oxidative stress and improves glucose uptake via Mas receptor in adipocytes[J]. Acta Diabetol, 2012, 49: 291- 299.
[10] Fonseca-Alaniz MH, Higa TS, Ferraz-De-Campos TB,etal. ACE-modulated adiposity is related to higher energy expenditure and independent of lipolysis and glucose incorporation into lipids in adipocytes[J]. Physiol Genomics, 2017, 49: 712- 721.
[11] Tsukuda K, Mogi M, Iwanami J,etal. Enhancement of adipocyte browning by angiotensin Ⅱ type 1 receptor blockade[J]. PLoS One, 2016, 11:1- 12
[12] Chan S, Lau YS, Miller AA,etal. Angiotensin Ⅱ causes beta-cell dysfunction through an ER stress-induced proinflammatory response[J]. Endocrinology, 2017, 158: 3162- 3173.
[13] Xuan X, Gao F, MaX,etal. Activation of ACE2/angiotensin (1- 7) attenuates pancreatic beta cell dedifferentiation in a high-fat-diet mouse model[J]. Metabolism, 2017, 81:83- 96.
[14] Frantz E, Medeiros RF, Giori IG,etal. Exercise train-ing modulates the hepatic renin-angiotensin system in fructose-fed rats[J]. Exp Physiol, 2017, 102: 1208- 1220.
[15] Moreira C, Lourenco FC, Mario EG,etal. Long-term effects of angiotensin-(1- 7) on lipid metabolism in the adipose tissue and liver[J]. Peptides, 2017, 92: 16- 22.
[16] Surapongchai J, Prasannarong M, Bupha-Intr T,etal. Angiotensin Ⅱ induces differential insulin action in rat skeletal muscle[J]. J Endocrinol, 2017, 232: 547- 560.
[17] Abuohashish HM, Ahmed MM, Sabry D,etal. Angiotensin (1- 7) ameliorates the structural and biochemical alterations of ovariectomy-induced osteoporosis in rats via activation of ACE-2/Mas receptor axis[J]. Sci Rep, 2017, 7: 1- 11.
[18] Boschmann M, Ringel J, Klaus S,etal. Metabolic and hemodynamic response of adipose tissue to angiotensin Ⅱ[J]. Obes Res, 2001, 9: 486- 491.
[19] Pan Y, Qiao QY, Pan LH,etal. Losartan reduces insulin resistance by inhibiting oxidative stress and enhancing insulin signaling transduction[J]. Exp Clin Endocrinol Diabetes, 2015, 123: 170- 177.
[20] Santos SH, Braga JF, Mario EG,etal. Improved lipid and glucose metabolism in transgenic rats with increased circulating angiotensin-(1- 7)[J]. Arterioscler Thromb Vasc Biol, 2010, 30: 953- 961.
[21] Santos SH, Fernandes LR, Mario EG,etal. Mas deficiency in FVB/N mice produces marked changes in lipid and glycemic metabolism[J]. Diabetes, 2008, 57: 340- 347.
[22] Marcus Y, Shefer G, Sasson K,etal. Angiotensin 1- 7 as means to prevent the metabolic syndrome: lessons from the fructose- fed rat model[J]. Diabetes, 2013, 62: 1121- 1130.
[23] Sysoeva VY, Ageeva LV, Tyurin-Kuzmin PA,etal. Local angiotensin Ⅱ promotes adipogenic differentiation of human adipose tissue mesenchymal stem cells through type 2 angiotensin receptor[J]. Stem Cell Res, 2017, 25: 115- 122.
[24] Than A, Leow MK, Chen P. Control of adipogenesis by the autocrine interplays between angiotensin 1- 7/Mas receptor and angiotensin Ⅱ/AT1 receptor signaling pathways[J]. J Biol Chem, 2013, 288: 15520- 15531.
[25] Jones BH, Standridge MK, Moustaid N. Angiotensin Ⅱ increases lipogenesis in 3T3-L1 and human adipose cells[J]. Endocrinology, 1997, 138: 1512- 1519.
[26] Caminhotto RO, Sertie RA, Andreotti S,etal. Renin-angiotensin system blockers regulate the metabolism of isolated fat cellsinvitro[J]. Braz J Med Biol Res, 2016, 49:1- 8.
[27] de Macedo SM, Guimarares TA, Andrade JM,etal. Angiotensin converting enzyme 2 activator (DIZE) modu-lates metabolic profiles in mice, decreasing lipogenesis[J]. Protein Pept Lett, 2015, 22: 332- 340.
[28] Mario EG, Santos SH, Ferreira AV,etal. Angiotensin-(1- 7) Mas-receptor deficiency decreases peroxisome proliferator-activated receptor gamma expression in adipo-cytes[J]. Peptides, 2012, 33: 174- 177.
[29] Goossens GH, Blaak EE, Arner P,etal. Angiotensin Ⅱ: a hormone that affects lipid metabolism in adipose tissue[J]. Int J Obes (Lond), 2007, 31: 382- 384.
[30] Oh YB, Kim JH, Park BM,etal. Captopril intake decreases body weight gain via angiotensin-(1- 7)[J]. Peptides, 2012, 37: 79- 85.
猜你喜欢
现代临床医学(2022年4期)2022-09-29
清华金融评论(2022年4期)2022-04-13
昆明医科大学学报(2022年1期)2022-02-28
中老年保健(2021年9期)2021-08-24
昆明医科大学学报(2021年8期)2021-08-13
空间科学学报(2021年2期)2021-07-21
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
天然产物研究与开发(2018年9期)2018-10-08
中成药(2018年2期)2018-05-09
