
2017年全球低致病性禽流感疫情分析
2018-05-09 06:12陈泽金兰邹然
中国动物检疫 2018年5期
陈泽金,兰邹然
(1. 福建省南安市农业局,福建南安 362300;2. 山东省动物疫病预防与控制中心,山东济南 250022)
禽流感(Avian Influenza,AI)是由粘病毒科A型流感病毒(Avian influenza virus,AIV)引起的禽类传染病。根据毒力不同,AIV可分为高致病性(HPAIV)和低致性(LPAIV)两种。HPAIV因致病性强,能造成高发病率和病死率,受到养殖从业者的高度关注,而LPAIV致病性较弱,但对家禽生产存在一定影响,且常常与其他疫病混淆,不易鉴别诊断,往往被忽视。然而LPAIV通过长时间进化,有些毒株已对养禽生产造成较大影响[1-2]。其一,LPAIV通过自然重组、基因突变等方式,可以变异为HPAIV[1],比如我国2013年出现的H7N9亚型LPAIV,到2016年底进化成为HPAIV,并于2017年导致多起疫情暴发[3];其二,LPAIV的许多亚型也是人流感的潜在感染亚型,比如H3、H5和H7亚型,其中H3亚型是引起人流感的主要亚型[4-5]。为此,基于世界动物卫生组织(OIE)全球动物卫生信息系统的低致病性禽流感疫情报告数据,统计分析全球2017年低致病性禽流感流行态势,从而为我国禽流感防控提供参考。
1 疫情概况
2017年,全球共有柬埔寨、法国、德国、荷兰、智利、美国、多米尼加、南非、利比亚以及中国台湾等10个国家或地区报告低致病性禽流感疫情57起,涉及易感禽568 000余只,其中发病23 040只(发病率约为4.05%),死亡22 547只,销毁282 299只(表1),造成了一定的经济损失。

表1 2017年全球各国报告的低致病性禽流感疫情统计
2 疫情分布
2.1 毒株分布
从流行毒株情况看,2017年全球流行的低致病性禽流感亚型包括H5N1、H5N2、H5N3、H5N6、H5N9、H7N3、H7N6和 H7N9等多个亚型。其中H5N2亚型疫情数最多,报告20起,其他较多的是H5N3和H5N1亚型,各报告11起,再次是H7N9亚型,报告6起(图1)。由此可见,2017年全球报告的低致病性禽流感疫情数虽然不多,但流行毒株类型较复杂。
2.2 空间分布
从空间分布看,疫情在全球呈散发态势,欧洲和美洲各有3个国家报告发生低致病性禽流感疫情,亚洲和非洲各有2个国家和地区报告该疫情。2017年1—4季度疫情分布见图2。
2.3 时间分布

图1 2017年不同亚型低致病性禽流感报告统计
从时间分布看,气温偏低的1—3月份报告的疫情数最多,共40起;气温偏高的7—9月份的最少,仅1起(图3)。
2.4 群间分布
从疫情发生场点类型看,绝大多数疫情发生在规模场,共54起;其次为散养户和村庄,共3起。屠宰场、自然公园、动物园、市场和森林等场点未报告发生疫情(图4)。
3 疫情特点
3.1 呈散发状态
从空间分布看,除大洋洲和南极洲外,其他4大洲均有疫情报告,但每个洲仅有2~3个国家或地区报告发生疫情,全年仅发生57起,呈明显的点状散发状态,未出现区域性暴发。同时其危害和影响与高致病性禽流感相比非常轻微,全年仅有2.3万多只禽发病,28万多只禽被销毁,仅相当于1起较大高致病性禽流感疫情导致的损失。

图2 2017年1—4季度全球低致病性禽流感疫情分布

图3 2017年各月份低致病性禽流感报告疫情统计
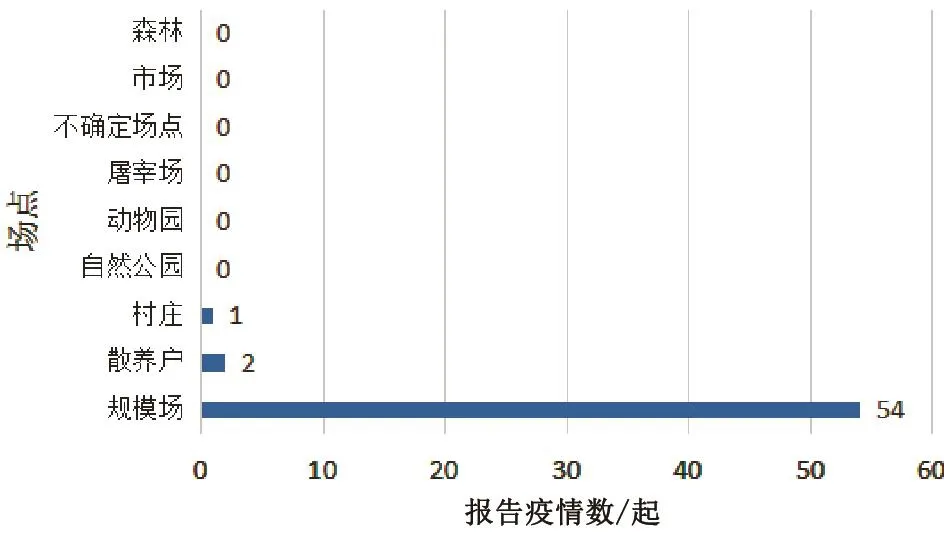
图4 2017年各场点报告疫情统计
3.2 亚型多样,但相对集中
2017年全球已确定的低致病性禽流感亚型共8个,包括H5N1、H5N2、H5N3、H5N6、H5N9、H7N3、H7N6和H7N9(部分H7亚型未鉴定N亚型),但报告的H5和H7亚型疫情数量最多,未报告发现对养禽生产影响较大且文献中经常报道的H9N2亚型[8-10]。这是否存在信息偏倚值得关注,或许与报告国更关注具有潜在高致病性威胁的H5和H7亚型有关。
3.3 寒冷季节多发
从疫情暴发的时间分布来看,疫情多发生于气温较低的冬春季节。如1—3月和12月,共报告疫情45起,占全年疫情的83.3%。AIV对温度的抵抗性有限,在炎热季节,病毒在外界环境中生存能力有限,不易引发疫情,而在寒冷季节却容易存活,从而造成疫情发生和传播。这是所有AIV的共同特点,包括H7N9流感。
3.4 规模场是重灾区
从全年报告的疫情来看,57起疫情中,有54起发生于规模化养禽场。这可能与规模养殖条件下,LPAIV可以在更多宿主中选择定殖有关,当然也可能与规模化养禽场的监测更受重视存在一定关系。
4 小结
综上所述,2017年全球低致性禽流感呈现点状散发特点,除大洋洲和南极洲外,其余4大洲均有国家或地区报告发生疫情;规模场是低致病禽流感疫情的主要发生地;报告并确定的低致病性禽流感血清亚型较多,共有8个亚型,但以H5和H7亚型引发的疫情数量居多;H5和H7亚型病毒有变异为高致病性的风险,应引起高度重视。
参考文献:
[1] 刁有祥,丁家波. 低致病性禽流感病毒对养禽业的危害[J]. 中国家禽,2006,28(6):1-3.
[2] 黄建龙,朱春霞,刘道新. 我国低致病性禽流感研究概况[J]. 动物医学进展,2014,35(4):121-124.
[3] 杜建,兰邹然,刘祥,等. 2017年第1季度全球禽流感疫情分析[J]. 中国动物检疫,2017,34(6):22-25.
[4] 张文形,姜庆五. 全球历年人甲型流感病毒H3A1抗原的分子进化研究[J]. 中华流行病学杂志,2005,26(11):843-847.
[5] 董丽波,张烨,温乐英,等. 1995—2005年中国H3N2亚型人流感病毒血凝素基因变异与流行相关性研究[J]. 病毒学报,2007,23(5):339-344.
[6] 朱迪国,宋建德,黄保续. 当前全球禽流感流行概况及特点分析[J]. 中国动物检疫,2015,32(3):41-47.
[7] 张晓红,张虎芳. 候鸟迁徙与禽流感(Avian Influenza,AI)传播[J]. 太原师范学院学报(自然科学版),2005,4(4):81-84.
[8] PATHAK A P,MURUGKAR H V,NAGARAJAN S,et al. Survivability of low pathogenic(H9N2)avian influenza virus in water in the presence of Atyopsis moluccensis(Bamboo shrimp)[J]. Zoonoses public health,2018,65(1):e124-129.
[9] YU G,WANG A,TANG Y,et al. Vertical transmission of H9N2 avian influenza virus in goose[J]. Frontiers in microbiology,2017,15(8):1559.
[10] 蒋文明,侯广宇,彭程,等. 鄱阳湖与青海湖地区野鸟H9N2亚型禽流感病毒的生物学特性[J]. 中国动物检疫,2017,34(6):10-13.
猜你喜欢
动物医学进展(2022年9期)2022-11-26
云南畜牧兽医(2022年4期)2022-08-19
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
中国土壤与肥料(2021年5期)2021-12-02
科技信息·学术版(2021年19期)2021-10-25
青年生活(2019年32期)2019-09-10
今日农业(2019年11期)2019-08-13
中成药(2017年12期)2018-01-19
中国猪业(2017年11期)2017-12-11
