
杜仲短周期矮林的密度效应1)
2018-05-04 08:21刘慧东丁欢欢朱景乐殷世雨李铁柱杜红岩孙志强
东北林业大学学报 2018年4期
刘慧东 丁欢欢 朱景乐 殷世雨 李铁柱 杜红岩 孙志强
(国家林业局泡桐研究开发中心,郑州,450003) (信阳市南湾实验林场) (国家林业局泡桐研究开发中心)
速生树种的短周期高密度矮林种植模式不但可在短期内实现生物质能源的大量供应,还可提供医药及其他工业领域所需的原材料[1]。这种种植模式具有可控制和可持续的特点,同时因其周期短、收益高[2],近年来在许多国家得到了迅猛的发展,例如用于提供生物质能源和造纸原料的杨树、柳树和桉树的短周期矮林模式[2-4];以及生产叶片用于天然药用成分提取的银杏矮林模式[5-6]。杜仲(EucommiaulmoidesOliver)是中国特有的贵重中药材和橡胶工业原料树种。但是,杜仲药用和杜仲胶产业化目前所面临的主要瓶颈问题是收获成本过高、原材料产量不足[7-8]。作为重要的工业原材料,如何短期内获取大量的叶和皮是杜仲产业化开发的关键问题。
杜仲适生于温带和亚热带的不同土壤条件,萌蘖和快速生长能力强[9],这些特点使杜仲适合于短轮伐期矮林栽植。短周期高密度矮林的生物量因树种、栽植密度、轮伐周期、根桩年龄等的不同会表现出较大的差异[10]。有关杜仲短周期高密度矮林模式的研究主要集中在以下几个方面,不同平茬方式对杜仲萌蘖能力的影响[11]、生长季内生物量的变化动态[12]、叶片内杜仲胶含量和相对分子质量的年变化动态[13]及枝丫材的利用和其醋液的抑菌活性[14-15]等。截至目前,几乎没有关于杜仲矮林密度效应对生长性状和产量影响的相关报道。
密度效应是自然界植物种群普遍存在的特征。密度制约通过改变种群内每个植株可获得性资源的数量,使邻体间发生地上的光竞争和地下的水分与矿质营养竞争,从而引起植物形态特征及生长特性发生改变,进而影响生物量的变化[16]。合适栽植密度是最重要的优化光捕获、最大化增加产量的影响因素;过密自然稀疏,影响投入产出比;过稀杂草丛生,增加管理成本[17]。为解析杜仲短周期矮林模式下栽植密度对单株生长特点和生长性状及地上部分生物量的影响,本研究以2015年建立的包含5个栽植密度的短周期杜仲矮林为研究对象,于2016年10月详细对比了首次皆伐后不同密度杜仲单株在一个生长季内的生长性状及地上部分生物量。研究结果为杜仲矮林模式栽植密度的设置提供理论依据。
1 研究区域概况
试验地位于中国林业科学研究院经济林研究开发中心孟州基地。该基地南临黄河,属黄河滩地,大陆性暖温带季风型气候,年降水量约549.9 mm,年平均气温14.3 ℃,无霜期224 d。具有中部平原地区黄河古道沙地典型立地特征。沙壤土,土壤0~40 cm处的理化性质为:pH=8.67,有机质质量分数4.07 g·kg-1,总氮质量分数0.03 g·kg-1,有效氮质量分数18.79 mg·kg-1,有效磷质量分数4.80 mg·kg-1,有效钾质量分数156.62 mg·kg-1。
2 研究方法
2.1 试验设计
杜仲矮林于2015年3月营造,总面积3 000 m2。栽植前开沟1.0 m宽、0.8 m深,隔1.0 m开一个沟,沟内施农家肥后回填。采用宽窄行栽植模式,宽行距1.0 m,窄行距0.5 m,窄行内株距分别为1.0、0.8、0.6、0.4和0.2 m,苗木为1年生实生苗,平均苗高1.2 m,分别相当于13 333、16 667、22 222、33 333和66 667株·hm-2。按完全随机区组试验设计,设5种栽植密度,3个重复,每小区栽植面积为120 m2(东西长20 m,南北宽6 m)。于2015年10月平茬,2016年春季在杜仲生长初期进行抹芽,每株留1个主芽。适时浇水、除草,试验期间不施肥。
2.2 生长指标和生物量的测定
2016年10月9日随机剪取所有不同密度处理的苗木各50株,地上部分留5 cm,分别用卷尺和游标卡尺测量植株的株高和基径,统计枝干上的单株叶片数量和分枝数,随机选取茎干的中间1/3部分的4片叶,称量其鲜质量,将每个叶片拍照,随后采用万深叶面积仪(杭州万深LA-S)测其叶面积;剪取每株苗木中间20 cm的茎干,剥皮后分别称皮和杆的鲜质量。将采集的叶片、树皮和枝干带回室内于65 ℃烘干后再分别称其质量(叶(W1)、皮(W2)、杆(W3))。
单叶干质量=W1/4,单位面积叶片质量=单叶干质量/单叶面积,叶面积指数=单叶面积×单株叶片数×单位面积株数/土地面积,单株叶干质量=W1/4×单株叶片数,单株皮干质量=W2/20×株高,单株杆干质量=W3/20×株高,单位面积叶生物量=单株叶干质量×密度,单位面积皮生物量=单株皮干质量×密度,单位面积杆生物量=单株杆干质量×密度。
2.3 数据处理及统计分析
采用SPSS软件(20.0版)进行数据分析,用sigmaplot 10.0作图。采用单因素方差分析(p<0.05)检验不同密度的植株各生长性状的差异并进行Duncan多重比较,用双变量Person双尾检验(p<0.05)进行各指标间的相关性分析。
3 结果与分析
3.1 不同密度下单株生长性状
不同密度下杜仲单株的平均分枝数、基径、株高、单株叶片数、单位面积叶片质量和叶面积指数差异极显著,而平均单叶干质量和单叶面积差异不显著(F=1.117,p>0.05;F=2.557,p>0.05)。栽植密度13 333、16 667、22 222、33 333株·hm-2的分枝数在4.2~4.6之间,相互差异不显著,但均显著多于栽植密度66 667株·hm-2的分枝数1.4(表1)。
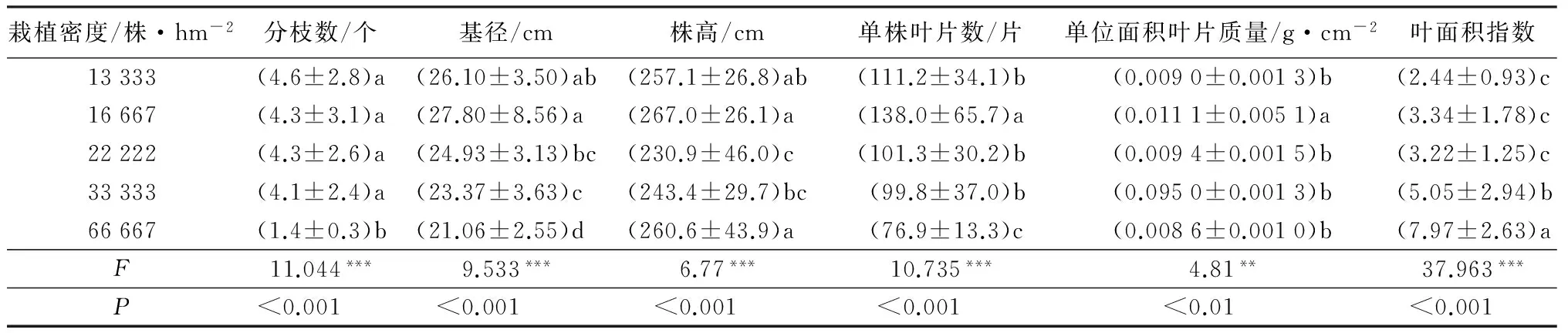
表1 不同栽植密度杜仲生长指标比较
注:表中数据为“平均数±标准差”;*** 表示p<0.001差异极显著;**表示p<0.01差异非常显著。
单株的平均分枝数与密度呈极显著负相关(R2=-0.424,p<0.001),与单叶干质量也呈显著负相关(R2=-162,p<0.05),而与基径(R2=0.297,p<0.001)和单株叶片数(R2=0.516,p<0.001)呈显著正相关(表2)。平均基径随着密度增加而减小,表现出与密度的极显著负相关关系(R2=-0.372,p<0.001)(表2);在66 667株·hm-2的最高密度下,平均基径比密度16 667株·hm-2的小24%(表1)。同时,平均基径与株高(R2=0.293,p<0.001)、单株叶片数(R2=0.438,p<0.001)呈显著正相关关系(表2)。
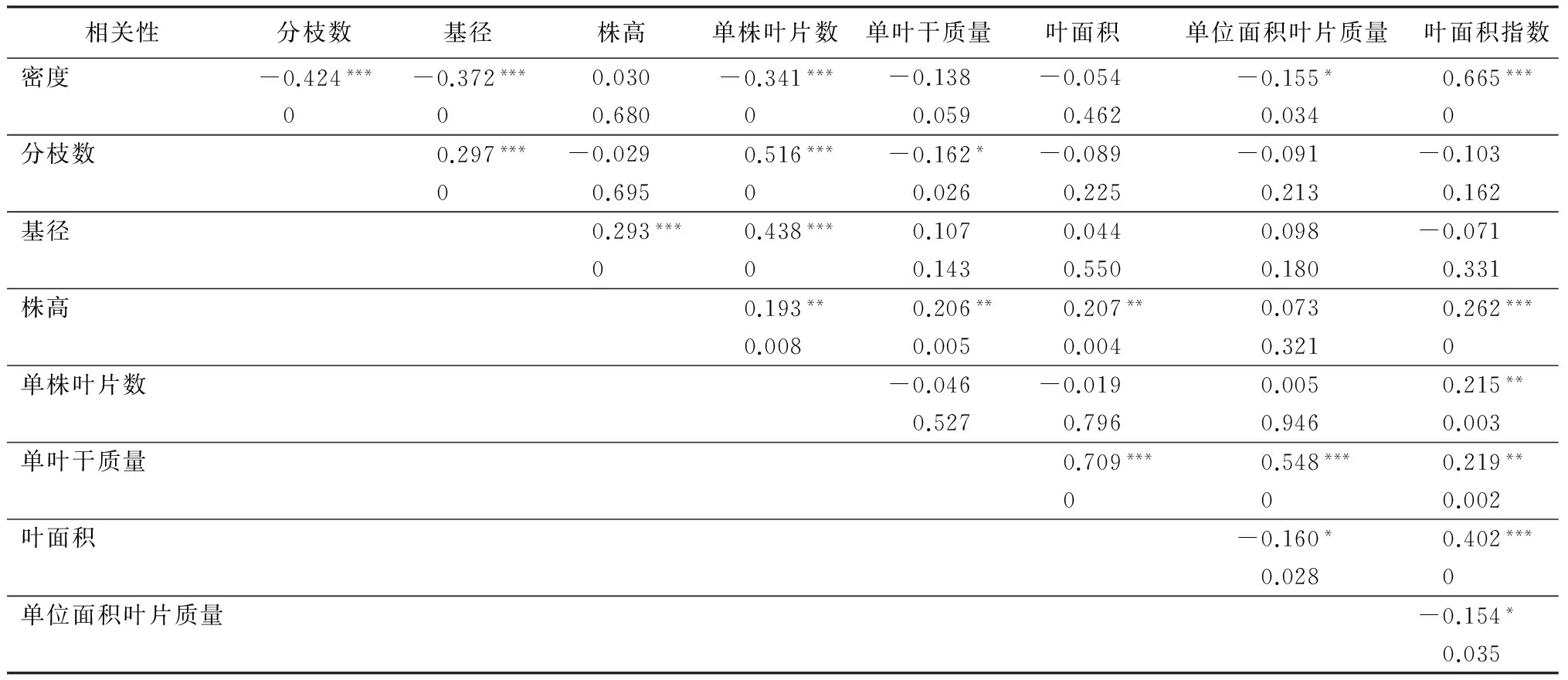
表2 杜仲生长指标与栽植密度的相关性分析
注:*** 表示p<0.001极显著;** 表示p<0.01非常显著;*表示p<0.05显著。
平均株高与密度没有显著的相关关系(p>0.05),但与单株叶片数呈显著的正相关(R2=0.193,p<0.01)(表2)。密度16 667和66 667株·hm-2的平均株高分别达到267.0和260.6 cm,与密度13 333株·hm-2的257.1 cm差异不显著,但显著高于密度22 222、33 333株·hm-2的株高(表1)。
单株叶片数与密度呈显著的负相关(R2=-0.341,p<0.001),而与叶面积指数呈显著正相关(R2=0.215,p<0.01)(表2)。密度16 667株·hm-2的单株叶片数量最多,达到137片·株-1,显著多于其他几个密度的单株叶片数;密度13 333、22 222、33 333株·hm-2的单株叶片数在100~138片之间,且彼此间差异不显著,但整体上均显著高于密度66 667株·hm-2的单株叶片数(表1)。
叶面积指数与密度呈显著的正相关关系(R2=0.665,p<0.001)。密度66 667株·hm-2的叶面积指数最大,达到7.98,显著高于其他几个密度的叶面积指数。
单位面积叶片质量与密度呈显著负相关(R2=-0.155,p<0.05)。密度16 667株·hm-2的单位面积叶片质量0.011 1最高,显著高于其他几个密度的单位面积叶片质量;密度13 333、22 222、33 333、66 667株·hm-2的单位面积叶片质量差异不显著,在0.008 6~0.009 5 g·cm-2。
3.2 单株生物量和单位面积产量的密度效应
单株叶干质量(R2=-0.380,p<0.001)、单株皮干质量(R2=-0.358,p<0.001)、单株杆干质量(R2=-0.417,p<0.001)与密度均呈极显著的负相关(表3),即密度越小,单株的生物量越大。不同密度下杜仲单株叶干质量(F=9.238,p<0.001)、单株皮干质量(F=14.311,p<0.001)、单株杆干质量(F=16.069,p<0.001)差异均达到极显著水平(表4)。在密度13 333株·hm-2条件下,平均单株叶干质量、皮干质量和杆干质量为205.91、43.31和317.71 g,分别是密度66 667株·hm-2的2.0、1.8、1.9倍。
在所有指标中,地上部分生物量的最优决定因素是叶面积指数,单位面积叶、皮和茎干生物量与叶面积指数呈极显著正相关关系,相关系数分别达0.906,0.700和0.674(表3)。不同密度下单位面积叶生物量(F=21.129,p<0.01)、皮生物量(F=67.031,p<0.01)、杆生物量(F=46.597,p<0.01)均差异极显著(表4)。密度66 667株·hm-2下单位面积的叶生物量、皮生物量、杆生物量分别为6 864.84、1 605.73、11 445.30 kg·hm-2,分别是密度13 333株·hm-2的2.5、2.8、2.7倍。同时,上述三者均与密度呈极显著的正相关外,还与单株的株高呈极显著正相关关系。
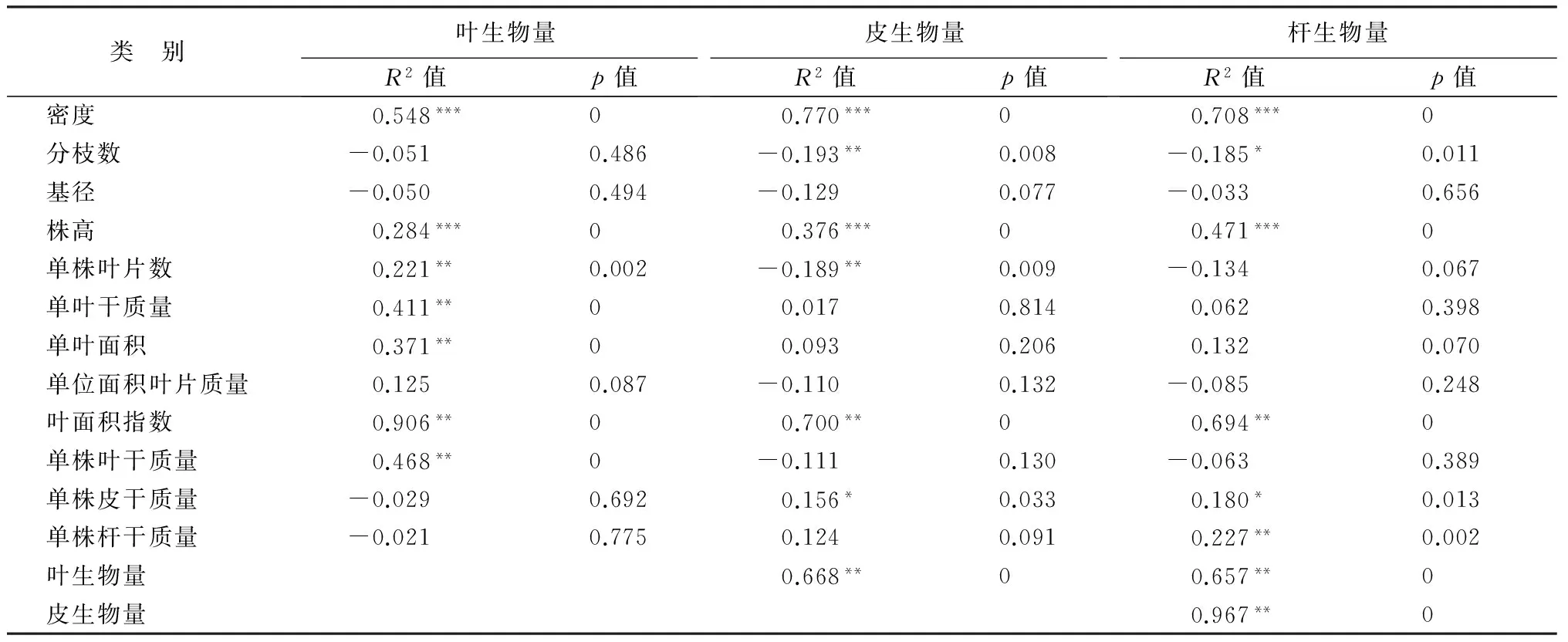
表3 地上部分各器官生物量与生长指标的相关性分析
注:*** 表示p<0.001极显著;** 表示p<0.01非常显著;*表示p<0.05显著。
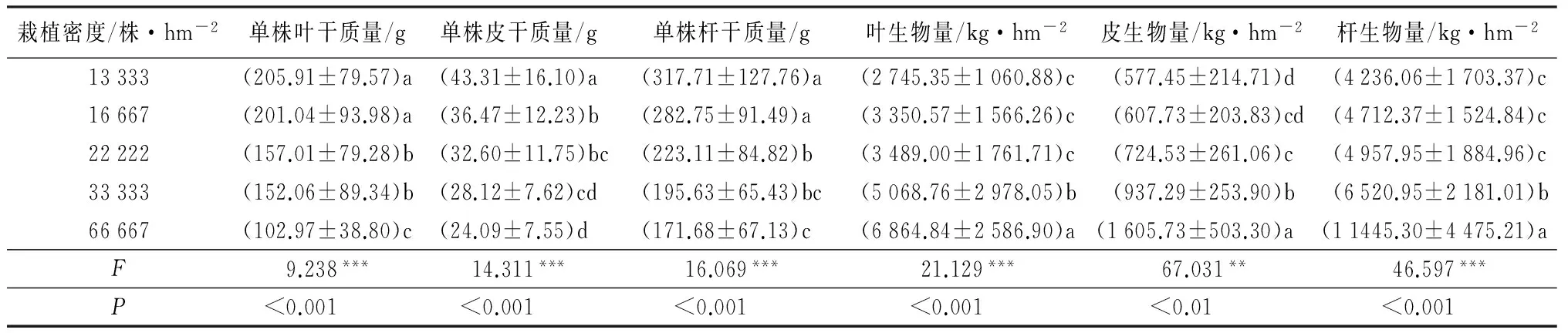
表4 不同栽植密度杜仲单株干质量与单位面积生物量的比较
注:表中数据为“平均数±标准差”;*** 表示p<0.001极显著;** 表示p<0.01非常显著。
4 讨论与结论
生长性状的变化是密度效应的一种表现[18]。栽植密度的大小直接影响和改变着单个植株的生长特征[19]。本研究中,33 333株·hm-2及以下密度的分枝数在一个范围变化,但没有明显差异,这意味着分枝数的密度效应在密度大于33 333株·hm-2时解除。基径和单株叶片数量的变化趋势一致,分别在密度16 667株·hm-2时达到最大值,这说明适合杜仲单株直径和叶片萌生的合适密度是16 667株·hm-2。株高和单位面积叶片质量随密度的变化均在密度66 667株·hm-2时出现最大值,但株高同时在密度66 667株·hm-2时也表现为最大值,这可能是高密度时顶端优势明显,而随着密度的减小,顶端优势缓解,侧枝发展明显,植株高生长减少;达到一定密度时,生长资源充足,高生长增加;但密度过低时,杂草等外界因素会抑制植物的高生长[18,20]。这也验证了不同密度强度下,植物可以通过调节自身生长形态,从而改变生物量的分配,实现资源利用的最优化[21-22]。由于植物自身的生物学特性,植物各个部分的生长存在竞争性,但又具有明显的相关性[16,22]。分枝数与基径呈显著的正相关,推测这与密度的大小密切相关,密度较小对应的植株横向生长空间大,基径和分枝数的增量就大。同时本研究中基径和株高存在极显著的正相关,没有出现过强的横向生长抑制纵向生长的彼此抑制关系。
叶片是植物截获光能的物质载体,合理的叶面积指数是反映植物群体光合能力的重要指标[23]。在植物的营养生长阶段,植株叶面积指数随密度的增大表现出明显的增大的趋势[24-26],本研究结果也证实了这一点;同时,在所有指标中,地上部分生物量的最优决定因素是叶面积指数,杜仲矮林群体叶面积指数随密度的增大而明显增大。由于不同密度条件下,平均单叶面积差异不显著,因此,单位面积的叶片数量是单位面积生物量的决定因素。
植物生长性状随密度的变化是植物适合度的重要体现,其充分反映了植物应对竞争所采取的环境资源获取策略和个体间生物量分配趋向的平衡[19,27]。有研究表明,林分中单株及其各组分的生物量均随林分密度的增加而降低[28]。本研究结果与之对应,杜仲矮林单株叶干质量、皮干质量、杆干质量随密度的增大均显著降低。密度过大,个体相互争夺资源,单株生物量必然减少[29-30],这既是密度影响到植物的生长特性所致,也是生态条件限制的必然结果[31]。
杜仲短周期密集矮林以提供杜仲胶和有效活性成分工业化开发的原材料为目标,因此杜仲叶片和树皮的年产量是衡量这种栽培模式的重要指标,而栽植密度是决定地上部分生物量的关键因素。杜仲矮林产量是矮林单株生物量与密度相乘之积,密度过大或过小都会严重影响产量[31]。本研究中设置的5个密度的叶生物量、皮生物量、杆生物量均随密度的增大而增大,并没有出现因密度过大而自我稀疏导致的生物量下降的趋势[22],这说明以生物量收获为目标的杜仲矮林密度可以更大,或单位面积的茎干数量更多,从而使产出增加。另一方面,该模式下产出的杜仲茎秆也提供了高质量的木质纤维素,用于制作纸浆[32]、生产生物质能源[15,33],还可作为食用菌培养基的添加料[34]。因此,对于以收获杜仲矮林叶为目的的杜仲应用工业来说,应选择较大的密度,如66 667株·hm-2等。
[1] HAUK S, KNOKE T, WITTKOPF S. Economic evaluation of short rotation coppice systems for energy from biomass—a review[J]. Renewable and Sustainable Energy Reviews,2014,29(2):435-448.
[2] MAMASHITA T, LAROCQUE G R, DESROCHERS A, et al. Short-term growth and morphological responses to nitrogen availability and plant density in hybrid poplars and willows[J]. Biomass and Bioenergy,2015,81(6):88-97.
[3] VERLINDEN M S, BROECKX L S, CEULEMANS R. First vs. second rotation of a poplar short rotation coppice above-ground biomass productivity and shoot dynamics[J]. Biomass and Bioenergy,2007,73(4):174-185.
[4] KNAPIC S, PIRRALHO M, LOUZADA J L, et al. Early assessment of density features for 19 Eucalyptus species using X-ray microdensitometry in a perspective of potential biomass production[J]. Wood Science and Technology,2014,48(1):37-49.
[5] 曹福亮,汪贵斌,郁万文.银杏叶用林定向培育技术体系的集成[J].南京林业大学学报(自然科学版),2014,38(6):146-152.
[6] 周志军,黄亿勋,王小民,等.银杏叶用林栽培技术[J].现代农业科技,2009(15):212.
[7] SUN Z Q, LI F D, DU H Y, et al. A novel silvicultural model for increasing biopolymer production fromEucommiaulmoidesOliver trees[J]. Industrial Crops and Products,2013,42(1):216-222.
[8] 魏艳秀,刘慧东,杜红岩,等.外源激素提高杜仲叶中有效成分含量的效果[J].中南林业科技大学学报,2017,37(5):42-49.
[9] 杜红岩,刘攀峰,孙志强,等.我国杜仲产业发展布局探讨[J].经济林研究,2012,30(3):129-133,144.
[10] SIMS R E H, SENELWA K, MAIAVA T, et al.Eucalyptusspecies for biomass energy in New Zealand-Part II: Coppice performance[J]. Biomass and Bioenergy,1999,65(4):333-343.
[11] 许喜明,徐咏梅,彭锋,等.多年生杜仲叶林栽培模式及其更新复壮[J].陕西林业科技,2006(1):22-24.
[12] 何文广,苏印泉.叶林模式杜仲生物量的动态研究[J].福建林业科技,2011,38(3):48-53.
[13] 何文广.杜仲叶中杜仲胶含量与相对分子质量的动态变化[J].经济林研究,2011,29(1):46-51.
[14] 高海霞,苏印泉,张强,等.杜仲叶林枝木醋液化学成分及抑菌活性研究[J].西北植物学报,2011,31(10):2106-2112.
[15] ZHU M Q, WEN J L, WANG Z W, et al. Structural changes in lignin during integrated process of steam explosion followed by alkaline hydrogen[J]. Applied Energy,2015,158:233-242.
[16] 列志旸,许松葵,薛立.不同密度大叶相思林的竞争效应[J].亚热带植物科学,2016,45(2):127-130.
[17] 赵宏魁,马真,张春辉,等.种植密度和施氮水平对燕麦生物量分配的影响[J].草业科学,2016,33(2):249-258.
[18] 刘战东,肖俊夫,于景春,等.春玉米品种和种植密度对植株性状和耗水特性的影响[J].农业工程学报,2012,28(11):125-131.
[19] 樊艳荣,陈双林,杨清平,等.毛竹林下多花黄精种群生长和生物量分配的立竹密度效应[J].浙江农林大学学报,2013,30(2):199-205.
[20] 马一学,阳会兵,陈金湘,等.棉花种植方式和密度效应研究[J].作物研究,2014,28(3):269-271,275.
[21] 黎磊,周道玮,盛连喜.密度制约决定的植物生物量分配格局[J].生态学杂志,2011,30(8):1579-1589.
[22] 郭子武,杨清平,李迎春,等.密度对四季竹地上生物量分配格局及异速增长模式的制约性调节[J].生态学杂志,2013,32(3):515-521.
[23] 贺昆仑,杜明伟,田晓丽,等.棉花种植密度对LAI-2200植物冠层分析仪测定叶面积指数的影响[J].作物杂志,2015(5):123-127.
[24] 冯尚宗,王世伟,彭美祥,等.不同种植密度对夏玉米产量、叶面积指数和干物质积累的影响[J].江西农业学报,2015,27(3):1-5.
[25] 胥少东,许培.不同密度对玉米新品种浚单0898叶面积指数、干重和产量的影响[J].农业科技通讯,2015(6):56-58.
[26] 张晓艳,杜吉到,郑殿峰,等.密度对大豆群体叶面积指数及干物质积累分配的影响[J].大豆科学,2011,30(1):96-100.
[27] 毕常锐,白志英,杨訸,等.种植密度对小麦群体光能资源利用的调控效应[J].华北农学报,2010,25(5):171-176.
[28] 叶绍明,覃连欢,龙滔,等.尾叶桉人工林生物量密度效应研究[J].安徽农业科学,2010,38(21):11594-11596,11607.
[29] 庞均喜.陕南地区不同种植密度对甜玉米信甜502的产量效应研究[J].现代农业科技,2015(5):38-42.
[30] ECHARTE L, MAGGIORA A D, CERRUDO D, et al. Yield response to plant density of maize and sunflower intercropped with soybean[J]. Field Crops Research,2011,121(3):423-429.
[31] 杜心田,王同朝.作物密度效应递增律及其意义[J].河南科学,2003,21(6):733-737.
[32] 赵泾峰,冯德君,王新爱.一年生杜仲平茬枝木材纤维形态与密度的研究[J].陕西林业科技,2003(1):7-9.
[33] ZHU M Q, WANG Z W, WEN J L, et al. The effects of autohydrolysis pretreatment on the structural characteristics, adsorptive and catalytic properties of the activated carbon prepared fromEucommiaulmoidesOliver based on a biorefinery process[J]. Bioresource Technology,2017,232:159-167.
[34] 黄文豪,董儒贞,梁臣,等.杜仲枝、叶栽培功能型杜仲香菇技术[J].食用菌,2015(3):45-46.
猜你喜欢
安徽农学通报(2022年6期)2022-04-07
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
新疆农业科学(2020年1期)2020-02-14
中成药(2018年8期)2018-08-29
畜牧与饲料科学(2018年3期)2018-05-08
湖南林业科技(2017年1期)2017-02-06
陕西画报(2016年1期)2016-12-01
中国麻业科学(2015年5期)2015-12-28
小雪花·小学生快乐作文(2015年9期)2015-09-29
小雪花·小学生快乐作文(2015年9期)2015-09-29
