
鲢鱼酶解产物对酵母菌的抗冻保护作用
2018-05-03 02:02熊思佳
食品与机械 2018年2期
熊思佳
王发祥1,2
俞 健1,2
李向红1,2
王建辉1,2
郭星辛1,2
刘永乐1,2
(1.长沙理工大学化学与生物工程学院,湖南 长沙 410114;2.湖南省水生资源食品加工工程技术研究中心,湖南 长沙 410114)
冷冻面团技术是20世纪50年代末发展起来的面包生产新工艺,利用冷冻原理将面团冻结和冷藏,需要时再解冻、醒发和烘烤,可避免传统面包在保藏、运输、销售等环节的老化和品质劣变,确保消费者吃上新鲜面包[1]。目前,冷冻面团技术已在许多国家和地区相当普及,中国虽然起步较晚,但仍具有广阔的发展前景。然而,在冷冻面团实际生产中,酵母菌的耐冷冻性能一直是影响面团品质的技术难题[2]。目前,国内外不少学者已在耐冻酵母的选育上开展了大量工作[3-4],但其应用效果有限且安全性存在较大争议,因此酵母抗冻剂的优选和开发仍然是改善冷冻面团品质的最有效途径。冰结构蛋白是目前研究最多的一类酵母抗冻保护剂,但含量少且价格高,限制了其在冷冻面团中的应用[5];其它抗冻剂如甘油[6]、海藻糖[7]、γ-聚谷氨酸钠[8]也具有一定的酵母抗冻保护效果,但与冰结构蛋白相比,这些传统保护剂既没有抗氧化、降血压等功效,也不能改善冷冻面团的风味。
鲢鱼(silver carp,Hypophthalmichthysmolitrix)又名白鲢、鲢子,属鲤形目鲤科,因其生长速度快、疾病少、容易养殖,2016年养殖产量达4.506 6×106t,仅次于草鱼,占中国淡水鱼产量的16.01%。但鲢鱼肉质发面、肌间刺多、土腥味重,不仅食用品质差,而且价格低廉,是最典型的大宗低值淡水鱼,急需开发其高值化利用途径。研究发现,鲢鱼酶解产物具有较好的抗冻保护效果[9],同时兼具良好的抗氧化性能[10-11],若能将其应用于冷冻面团中酵母菌抗冻,不仅来源丰富、成本低廉,其丰富的游离氨基酸、短肽等还能增加面包的营养价值。目前,已有学者对鲢鱼酶解产物的抗冻保护性能进行研究[12],但没有将鲢鱼酶解产物作为酵母菌抗冻保护剂的相关报道,本试验在前期研究基础上,利用复合蛋白酶制备鲢鱼酶解产物,分析其对面包酵母冻融循环过程中存活率的影响,旨在为应用鲢鱼酶解产物提高冷冻面团中酵母菌的耐冻性能提供理论依据。
1 材料与方法
1.1 材料
新鲜鲢鱼:购自湖南省长沙县当地农民手中;
面包酵母:实验室分离保藏;
麦芽汁培养基、PDA培养基:广东环凯微生物科技有限公司;
彩虹预染超低分子量蛋白Marker:1.2~45.0 kD,中科瑞泰(北京)生物科技有限公司;
复合蛋白酶(来源于芽孢杆菌属,酶活力≥1.5 AU/g)、N-[三(羟甲基)甲基]甘氨酸(Tricine):上海笛柏化学品技术有限公司;
其它试剂均为国产分析纯或生化试剂;
电子天平:BL-920S型,日本岛津;
自动发酵罐:BIOTECH-5M-7000A型,上海保兴生物设备工程有限公司;
恒温水浴锅:DK-98-ⅡA型,天津市泰斯特仪器有限公司;
高速离心机:LG10-24A型,北京金立离心机有限公司;
低温高速冷冻离心机:Centrifuge 5430 R型,艾本德中国有限公司;
2) 移动基站。通过获取手机连接的基站信息在运营商的库中转换成对应的经纬度,将经纬度对应到地图上。基站定位的优点是只要有手机信号就能定位,在室内和室外均可,无成本;缺点是定位误差大,无法在室内进行精准定位。
pH计:DELTA 320型,梅多勒-托利多(上海)有限公司;
双垂直蛋白电泳仪:DYCZ-24 型,北京六一仪器厂。
1.2 方法
1.2.1 酶解产物的制备 新鲜鲢鱼宰杀取肉,多功能粉碎机捣碎,按固液比1∶5 (g/mL)的比例分散于水溶液中,加入自动发酵罐,升温至50 ℃,用1 mol/L NaOH调节pH至6.5,按鱼肉干重的3%加入复合蛋白酶,搅拌水解,水解过程中自动滴加1 mol/L NaOH,使pH在6.5±0.05内波动,分别于15,30,60,120,240 min后取样,90 ℃灭酶15 min,4 000×g离心收集上清液后,冷冻干燥,得到系列不同水解度的鲢鱼酶解产物[13]。
1.2.2 Tricine-SDS-聚丙烯酰胺凝胶电泳(PAGE) 电泳缓冲液、样本缓冲液、AB-3(49.5% T,3% C)和AB-6(49.5% T,6% C)凝胶储存液的配制及电泳过程均参考Schägger H[14]的方法。按表1中的配方分别制备分离胶、积层胶和浓缩胶,待凝固后加入电极缓冲液,点样10 μL,内槽加入阴极电极缓冲液,外槽加入阳极缓冲液,开始电泳,初始电压30 V,直到样品完全进入积层胶之后加压至100 V,到达底部之前停止;后固定并硝酸银染色,再进行拍照。
1.2.3 酵母菌培养液的制备 从斜面上取一环面包酵母接种至50 mL麦芽汁培养基中,28 ℃摇床培养24 h,为一级种子液;再取5 mL重新接种至50 mL麦芽汁培养基中,28 ℃摇床培养12 h,即为酵母菌培养液[15],备用。
1.2.4 酵母菌抗冻性能的分析 取4支装有9 mL无菌水的离心管,无菌条件下分别加入1 mL酵母菌培养液,震荡混匀制得10-1稀释度的菌悬液,取1支以涂布法进行平板稀释计数(PDA培养基固体平板),计算初始活菌数N0;其余3支置于-20 ℃冰箱冷冻保藏,每间隔1个冻融循环(-20 ℃冷冻16 h,4 ℃解冻8 h)取出1支进行平板稀释计数,计算活菌数Ni,反复冻融1~3个循环,按式(1)计算酵母菌的存活率。
(1)
式中:
P——第i个冻融循环的存活率,%;
i——冻融循环数;
Ni——第i个冻融循环活菌数,CFU/mL;
N0——初始活菌数,CFU/mL。
1.2.5 鲢鱼酶解产物对酵母菌的抗冻保护能力分析
(2) 酶解产物浓度对酵母菌存活率的影响:称取酶解30 min 的鲢鱼酶解产物,按1.2.5(1)的方法制备含终浓度为2%,4%,6%,8%酶解产物的酵母菌悬液,置于-20 ℃冰箱进行1个冻融循环,取出以涂布法进行平板稀释计数,计算酵母菌的存活率,确定酶解产物的浓度。
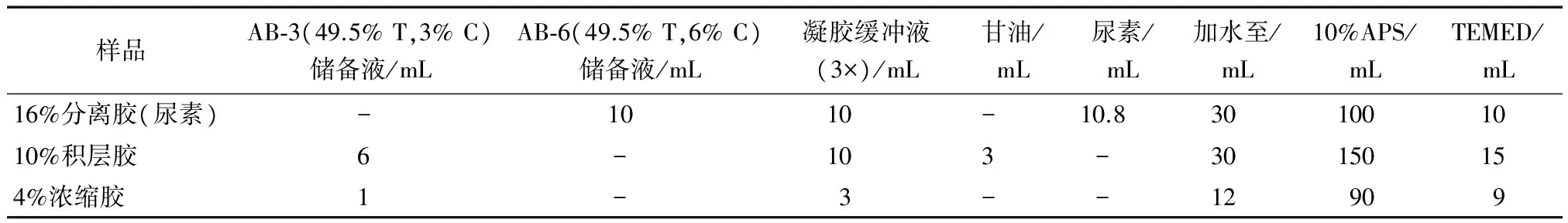
表1 制胶配方Table 1 Gel composition and preparation method for tricine-SDS-PAGE
2 结果与分析
2.1 鲢鱼酶解产物的制备
不同酶解时间的鲢鱼酶解产物的基本组成见表2。由表2 可知,随着酶解时间的延长,其水解度增加,但不同水解度酶解产物的水分、灰分、蛋白质(凯式定氮法)的含量区别不大。
2.2 酵母菌的抗冻性能
酵母菌经过冻融处理后的生长情况和存活率见图1。由图1可见,经过1个冻融循环处理后存活率仅有0.69%,第3个冻融循环存活率几乎为零,表明在没有保护剂的条件下面包酵母菌极不耐冻,要使其在冷冻后仍然维持存活,添加抗冻保护剂非常必要。
2.3 不同酶解产物对酵母菌的抗冻保护活性
酵母菌液中加入酶解不同时间制备的酶解产物,经过冻融处理后其生长情况和存活率见图2。由图2可见,甘油对酵母菌的抗冻保护效果明显,冻融循环后其存活率达63.5%;酶解产物组中酵母菌的生长情况及存活率都不及甘油组,但均高于空白组,表明不同酶解产物均对酵母菌有一定的抗冻保护作用。出现这种现象的原因是:冷冻过程中,细胞间隙中的水分首先形成冰晶,导致细胞间隙的蒸气压下降,形成细胞内外蒸气压差,在此作用下,细胞内水分外渗,细胞质浓度增大,进一步造成酶系统失活,细胞受损[16];同时胞内冰晶的形成也会损伤细胞膜,导致细胞死亡[17]。鲢鱼酶解产物具有抑制重结晶的作用,可减少冷冻过程中冰晶的形成,因而添加酶解产物后酵母菌的存活率增加[17];其中酶解15,30 min制备的鲢鱼酶解产物对酵母菌的抗冻保护效果较好,其存活率分别为22.91%,21.01%,随着酶解时间的延长,其抗冻保护活性逐渐降低。考虑到15,30 min酶解产物的抗冻效果相差不大,而30 min酶解产物的色泽和溶解性优于15 min酶解产物,故选用30 min酶解产物为后续的研究对象。
表2鲢鱼酶解产物的水解度和基本组成
Table 2 Degree of hydrolysis and composition of the enzymatic hydrolysates from silver carp

图1 冻融循环次数对酵母菌生长情况及存活率的影响Figure 1 Effect of freeze-thaw cycles on the growth of yeast and its survival rate
2.4 酶解产物浓度对酵母菌冻融存活率的影响
将30 min的酶解产物以终浓度为2%,4%,6%,8%的比例加入酵母菌悬液,冻融处理后分析其存活率,结果见图3。由图3可见,随着酶解产物浓度的增加,酵母菌的存活率显著增加,浓度为4%时,酵母菌的存活率达到90.73%,较阳性对照(8%甘油)提高了42.88%,说明4%的酶解产物对酵母菌的抗冻保护作用较好;酶解产物浓度继续增加到6%,8%时,酵母菌的存活率有所降低,可能与体系渗透压增大导致的细胞代谢损伤有关[18],但存活率仍大于80%,远优于阳性对照组(8%甘油)。

图2 添加不同酶解产物对酵母菌生长情况和存活率的影响Figure 2 Effect of different enzymatic hydrolysates on the growth of yeast and its survival rate
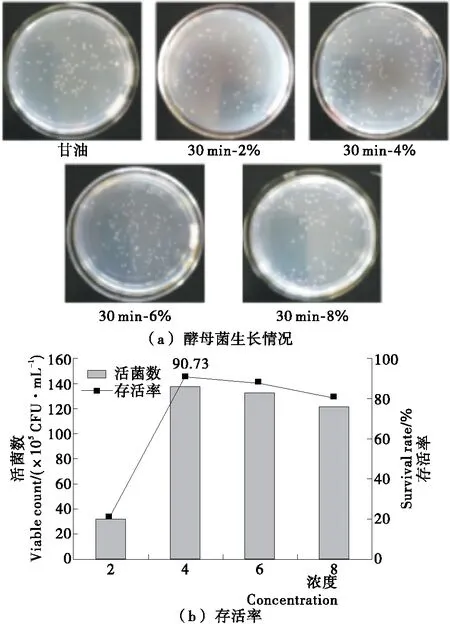
图3 酶解产物浓度对酵母菌生长情况及存活率的影响Figure 3 Effect of concentration of enzymatic hydrolysates on the growth of yeast and its survival rate
2.5 酶解产物的Tricine-SDS-PAGE分析
用Tricine-SDS-PAGE对不同时间鲢鱼酶解产物的分子量分布进行分析,结果见图4。由图4可见,与酶解前鲢鱼蛋白(0 min)相比,酶解产物的条带分布发生了较大变化。鲢鱼蛋白的电泳条带主要分布在16 kDa以上,尤其是40 kDa以上的区域,表明未酶解的鲢鱼蛋白分子量较大;而酶解产物的蛋白条带基本相似,且主要分布在16 kDa以下,随着酶解时间的延长,条带逐渐减弱,水解240 min后,条带已非常微弱,表明酶解过程中大分子量蛋白部分被逐渐水解为小分子量蛋白、肽或氨基酸[19]。此外,酶解开始后,在约13 kDa附近新增一条明显条带(图4箭头处),其信号随着酶解时间的延长呈逐渐减弱趋势,与图2中其抗冻保护效果的趋势基本一致,推测该组分很可能与酶解产物的抗冻保护活性有重要关系,但还需要进一步的试验验证。
3 结论
本研究通过复合蛋白酶有限酶解制备一系列不同酶解时间的鲢鱼酶解产物,分析了其对面包酵母菌的抗冻保护活性。结果表明,面包酵母耐冻性能较差,经过1个冻融循环处理后存活率仅有0.69%,而添加酶解产物能明显提高其冷冻存活率,其中酶解15,30 min 的酶解产物对酵母菌的抗冻保护效果最好,进一步研究表明,添加4%的30 min酶解产物能使酵母菌的冷冻存活率提高到90.73%,优于目前公认的对微生物抗冻保护效果较好的甘油。Tricine-SDS-PAGE分析发现,酶解产物的蛋白条带分布基本相似,条带信号随着酶解时间的延长逐渐减弱,其中新增了一条大约13 kDa的条带,可能与酶解产物的抗冻保护活性有重要关系,但还需要进一步的试验验证。本研究表明鲢鱼酶解产物可作为一种天然抗冻剂用于提高酵母菌在速冻和解冻过程中的存活率,为增强冷冻面团中酵母菌的耐冻性提供了试验依据。

图4 不同时间鲢鱼酶解产物的Tricine-SDS-PAGE图谱Figure 4 Comparison of peptide band distribution of different enzymatic hydrolysates
[1] 李爱华, 金茂国.冷冻面团技术——国内面包加工新模式[J].食品与机械, 2000(1): 14-15.
[2] 梁万礼, 邹恩坤.冷冻面团法生产面包的关键工艺讨论[J].粮食与食品工业, 2015, 22(2): 22-25.
[3] 李楠, 奚震, 任云霞, 等.耐冻性面包酵母菌种的选育及其特性的研究[J].食品与发酵工业, 2003, 29(11): 55-59.
[4] TRIPP M, LUSK L, RHODES T, et al.Method of expressing antifreeze proteins in yeast: US, 180524[P].1998-12-15.
[5] 任士贤.冰结构蛋白及在中式冷冻面团中的应用[D].无锡: 江南大学, 2010: 13-15.
[6] 黄卫宁, 吕莹果.一种用甘油处理酵母抗冻发酵生产冷冻面包面团及其制造方法: 中国, 200510040427.5[P].2005-06-06.
[7] 周洁, 周慧明.添加海藻糖对酵母抗冻能力的影响[J].食品工业科技, 2004(11): 64-66.
[8] 时晓剑, 缪冶炼, 卫昊, 等.γ-聚谷氨酸钠对面包酵母的抗冻作用及其机理[J].食品科技, 2012(10): 2-6.
[9] 李向红, 刘忠祥, 邓海萍, 等.鲢鱼酶解产物在冷冻鱼糜中的抗冻性能研究[J].食品与机械, 2015(3): 125-130.
[10] CHALAMAIAH M, DINESH K B, HEMALATHA R, et al.Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review[J].Food Chemistry, 2012, 135(4): 3 020-3 038.
[11] 李向红, 陈志军, 刘永乐, 等.鲢鱼酶解产物分子质量组成与抗氧化性[J].食品科学, 2013, 34(17): 28-32.
[12] 俞健, 王发祥, 邓海萍, 等.鲢鱼酶解产物在冷冻鱼糜中的抗冻性能研究[J].食品与机械, 2015(3): 125-130.
[13] 赵玉红.鲢鱼副产物蛋白酶解的研究[D].哈尔滨: 东北农业大学, 2000: 15-16.
[14] SCHAGGER H.Tricine-SDS-PAGE[J].Nat Protoc, 2006, 1(1): 16-22.
[15] WANG Shao-yun, ZHAO Jun, CHEN Lin, et al.Preparation, isolation and hypothermia protection activity of antifreeze peptides from shark skin collagen[J].LWT-Food Science and Technology, 2014, 55(1): 210-217.
[16] JIA Chun-li, HUANG Wei-ning, WU Chao, et al.Characterization and yeast cryoprotective performance for thermostable ice-structuring proteins from Chinese Privet (Ligustrum Vulgare) leaves[J].Food Research International, 2012, 49(1): 280-284.
[17] 孙丽洁, 张晖, 王立, 等.鱼皮抗冻多肽的制备及其对冷冻面团热力学性质的影响[J].食品与发酵工业, 2017, 43(7): 87-92.
[18] PATRICIA L P, JAMES H B, GRAHAM G S, et al.渗透压和乙醇对酵母活力和形态的影响[J].啤酒科技, 2004(9): 61-65.
[19] 张锐昌, 王绮, 张应龙, 等.Tricine-SDS-PAGE测定小麦蛋白酶解物分子量分布[J].食品研究与开发, 2012, 33(12): 168-171.
猜你喜欢
科学养鱼(2022年5期)2022-11-25
河南科学(2022年9期)2022-11-09
今日农业(2022年1期)2022-06-01
科学养鱼(2021年10期)2021-11-10
兰州工业学院学报(2021年3期)2021-06-29
今日农业(2020年24期)2020-03-17
海峡姐妹(2020年1期)2020-03-03
农民致富之友(2019年32期)2019-11-23
乡村科技(2018年11期)2018-02-11
北方交通(2016年12期)2017-01-15
