
玉米强杂种优势组合的基因型分析与分子预测
2018-04-19 09:50:54马海林白建荣杨瑞娟张丛卓
山西农业科学 2018年4期
李 锐 ,马海林 ,2,,白建荣 ,张 洁 ,关 超 ,杨瑞娟 ,张丛卓
(1.山西省农业科学院作物科学研究所,作物遗传与分子改良山西省重点实验室,山西太原 030031;2.山西省农业科学院玉米研究所,山西忻州 034000;3.山西大丰种业有限公司,山西太原 030031)
杂种优势是保证和提高玉米产量的重要原因。我国玉米生产和育种面临的主要限制因素是种质基础薄弱。解决此问题的重要途径是从玉米多样性中心系统引进优良种质,并根据杂种优势模式原理进行改良和利用。玉米是山西省的第一大作物,在山西有很多从事玉米育种的公司与研究单位。随着商业化育种的进程加快,育种家们从国内外引进了很多自交系,同时也对外引自交系按照育种目标进行改良。目前,很多自交系之间的亲缘关系不清,杂种优势群的归属不明。如果对这些材料进行广泛组配,通过后代的表现选育优良组合,但由于人力、物力、财力等因素的影响,只能进行很少一部分的工作,并且由于环境因素的影响选择效果并不会太理想。随着分子标记技术的迅速发展并逐步应用于玉米遗传多样性分析中,分子标记技术已经在玉米自交系的遗传多样性分析及杂种优势群的划分[1-7]、玉米品种指纹图谱的构建及遗传多样性分析[8-18]、杂交种真实性与纯度检测[19-21]等方面大量应用。很多育种人员迫切希望目前成熟的分子标记技术及已测序完成的玉米基因组信息能应用到实际的育种工作中,在基因型水平上预测杂种优势的亲本组合,增加育种工作的目的性和预测性,加快育种进程。但目前此方面的研究甚少。
本研究对山西大丰种业公司多年来收集和培育的大量自交系进行了基因型分析和杂种优势群划分;在此基础上,以一个杂优组合模式为例子,选择出了一些候选亲本自交系,并预测了一些潜在优势杂优组合,这将为玉米自交系的高效选育和杂优亲本的最佳组配提供依据和方法。
1 材料和方法
1.1 材料
1.1.1 自交系材料 1~81为收集与培育的自交系,82是先玉335、先玉508和先玉696的共同母本。83,84,85分别是先玉335、先玉508和先玉696的父本。86是郑单958的母本郑58,87是郑单958的父本昌7-2(表 1)。

表1 试验材料
1.1.2 SSR标记的引物 以中华人民共和国农业行业标准(NY/T 1432—2014《玉米品种鉴定技术规程SSR标记法》)的20对核心引物和20对辅助核心引物作为标记引物。
1.1.3 仪器与设备 扩增产物在美国PE公司Labchip GXTouch大分子分析系统进行分析。使用HT DNA Extended Range LabChip微流体实验室芯片;HT DNA 1K Reagent Kit试剂盒;并使用LABCHIP GXT software USB软件收集原始数据。Taq DNA聚合酶购于宝生生物工程(大连)有限公司;引物和dNTP等试剂购于上海生工生物技术有限公司。HTDNAExtended Range LabChip微流体实验室芯片和HT DNA 1K Reagent Kit试剂盒均购于美国PE公司中国代理公司。
1.2 试验方法
1.2.1 基因组DNA的提取、纯化和检测 采用混合提取法,每个供试材料2个样本,每样本由10个单株(随机取样)的叶片混合而成。采用CTAB法提取并纯化DNA,用分光光度计检测DNA的质量和浓度,调整DNA的工作液浓度为20 ng/μL,备用。
1.2.2 扩增与产物荧光检测 扩增反应在C1000扩增仪上进行。PCR反应总体系为20 μL:10×PCR Buffer(25 mmol/LMg2+)2 μL,2.5 mmol/LdNTPs 1 μL,10 μmol/L SSR 引物 1 μL,Taq DNA 聚合酶0.5 U,DNA模板 80 ng,ddH2O补足 20 μL。PCR 反应程序为:95℃预变性5 min,1个循环;94℃变性1 min,60℃复性30 s,72℃延伸1 min,共35个循环;最后于72℃延伸10 min。
1.3 数据统计和分析方法
1.3.1 遗传多样性分析与聚类 利用Reviewer software USB软件进行图像分析与数据收集。利用PowerMarker v3.25软件分析统计每对引物的等位基因个数、基因型个数、等位基因频率和多态性信息量(PIC),标记索引系数(Marker Index,MI),按Senior等提供的公式MI=Allele×PIC计算,Allele为该引物的等位基因数。利用NTSYS-pc2.1软件按照UPGMA方法对各材料进行聚类分析。
1.3.2 杂优类群内自交系差异位点比较 将杂优类群内自交系的40个位点的扩增带型进行比较,找出相互之间位点差异数及具体位点。
1.3.3 杂优组合的杂合位点数比较 将每个组配的杂优组合的2个亲本及每个源品种的2个亲本的40个位点的扩增带型组合成杂交种带型,统计杂合位点数,每一位点是2条带的是杂合位点。比较杂优组合与源品种的杂合位点数。
2 结果与分析
2.1 87个自交系的遗传变异分析
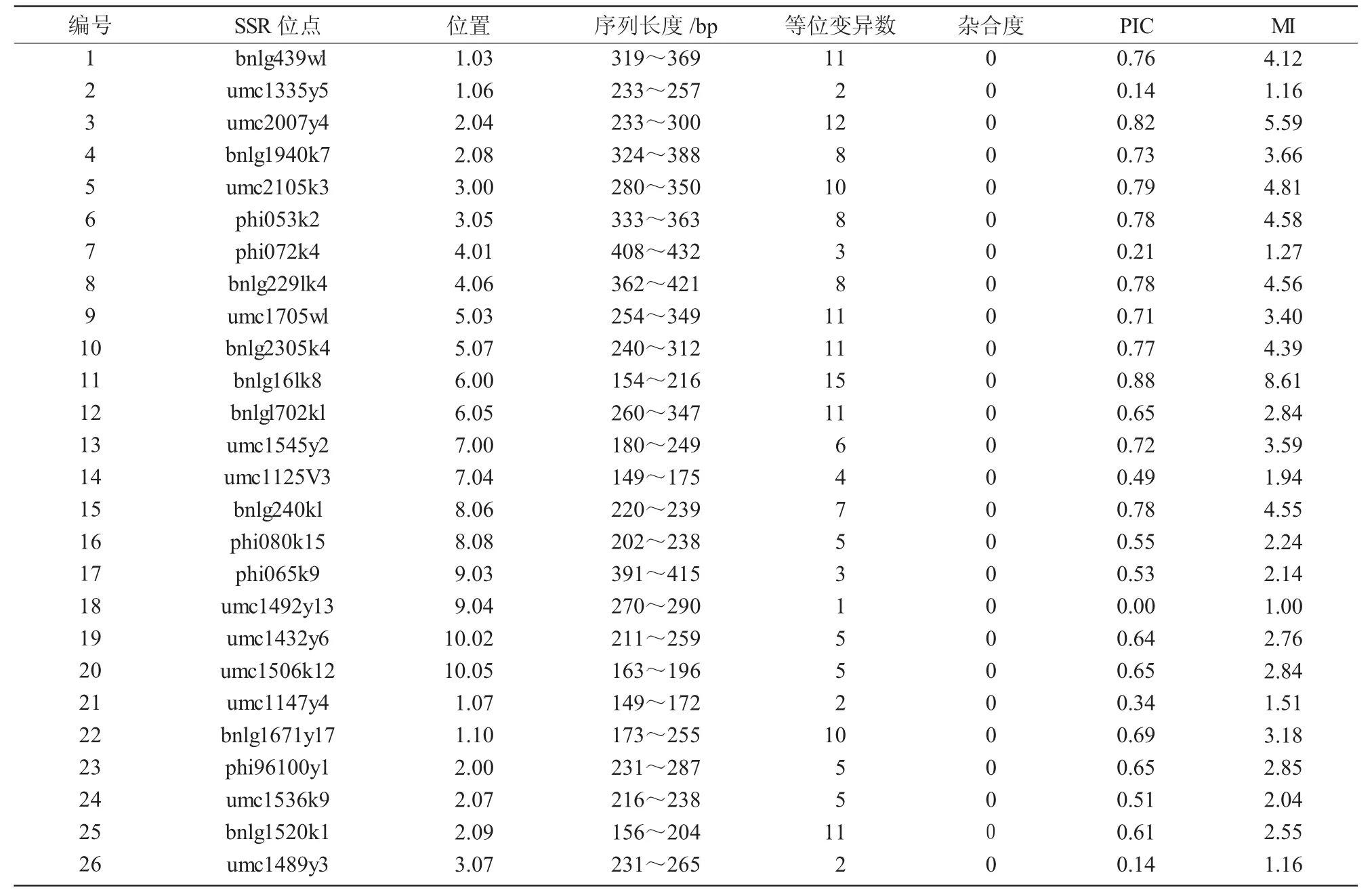
表2 SSR引物在87个玉米自交系中的等位变异及多态信息量
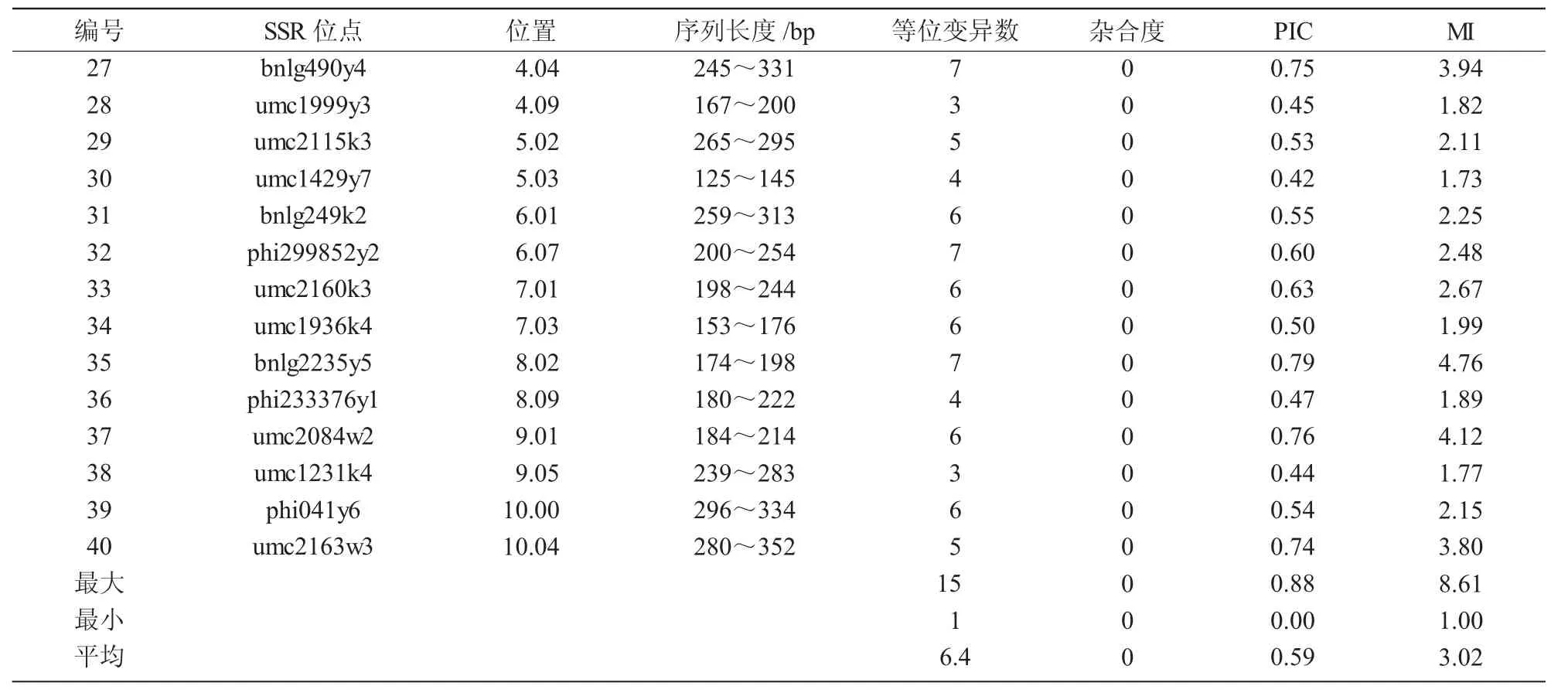
续表2
87个自交系在40个SSR位点检测到的遗传变异汇总如表2所示。共检测出256个等位变异,每对引物检测出1~15个等位变异,平均6.4个;其中,bnlg16lk8位点有15个等位变异,9个位点的等位变异数在10个以上,反映了在这些位点有高度的等位变异性;有7个位点只检测出3个及以下等位变异,其中,umc1492y13是一态,表明这些材料在这些位点高度纯合。40个位点的多态性信息量PIC值变化范围在0.00~0.88,平均为0.59;PIC值在0.8以上有2个位点。标记索引指数MI变化范围在1.00~8.61,平均为3.02。PIC值和标记索引指数MI值表现的趋势和等位变异数的趋势是类似的。这些材料的杂合度均为0,表明参试自交系都为纯系。
2.2 聚类分析
聚类分析结果显示(图1),大部分材料之间的遗传相似系数在0.77~0.98。在遗传相似系数大约为0.80水平上可以将所有材料分为4大类。其中,第Ⅰ类包括4小类,第1小类包括82(PH6WC),表明此群为Reid种群;第2小类包括郑58;第3小类和第4小类来源不清。第Ⅱ类分为2组,组1包括2个材料;组2分为2个组,其中一组包括83(PH4CV为335的父本)、84(PHB1M为508的父本)和85(PH5AD为696的父本),表明此类属于Lancaster种群。第Ⅲ类包括9个材料,其中包括87(昌7-2),本群为塘四平头群材料;第Ⅳ类群包括4个材料,可能属于新的优势类群。
2.3 杂优模式中亲本群的选择
从图1可以看出有许多类群以及它们的亲缘关系,但是它们组成的杂种模式是否是杂种优势模式,还需要进行许多研究。但是,我们知道先玉品种类的亲本组合是杂种优势模式之一,即Reid×Lancaster模式,但它们有共同的母本和各自的父本。实际育种工作中,很多育种家也是通过用本地种质改良先玉号的亲本获得改良自交系,以保持该模式的杂种优势。从图1可以看出,42,43,44和82(先玉母本) 构成了先玉母本群;13,14,15,16,17,18 和84(508父本)构成了 508父本群;19和 83(335父本)构成了335父本群;30和85(696父本)构成了696父本群。
2.4 先玉模式强优势组合的亲本预测
在选择了改良亲本群后,将对亲本进行组合。从表3可以看出,先玉335、先玉508和先玉696的杂合位点数分别为28,27,25个。82与各类父本组合后,杂合位点数少于42与各类父本组合的杂合位点数;43分别与各类父本组合后,组合杂合位点数明显少于相应的对照,44分别与各类父本组合后,组合杂合位点数明显多于相应的对照。因此,选择母本为 42 和 44,父本选择 13,14,15,16,17,18,19,30 及 83(335 父本)、84(508 父本)、85(696父本)进行组配。42与16,17组配后的杂合位点数是25,都比508杂合位点少,从组合表中去除;42与83(335父本)、84(508父本)、85(696父本)组合后,分别与先玉335、先玉508、先玉696差异仅为2个位点,视为同质品种,也去除。42与19组配后的杂合位点数是27,虽然没有335杂合位点多,但也属于多位点,应考虑其中。经比较后形成了推荐杂优组合(表4)。
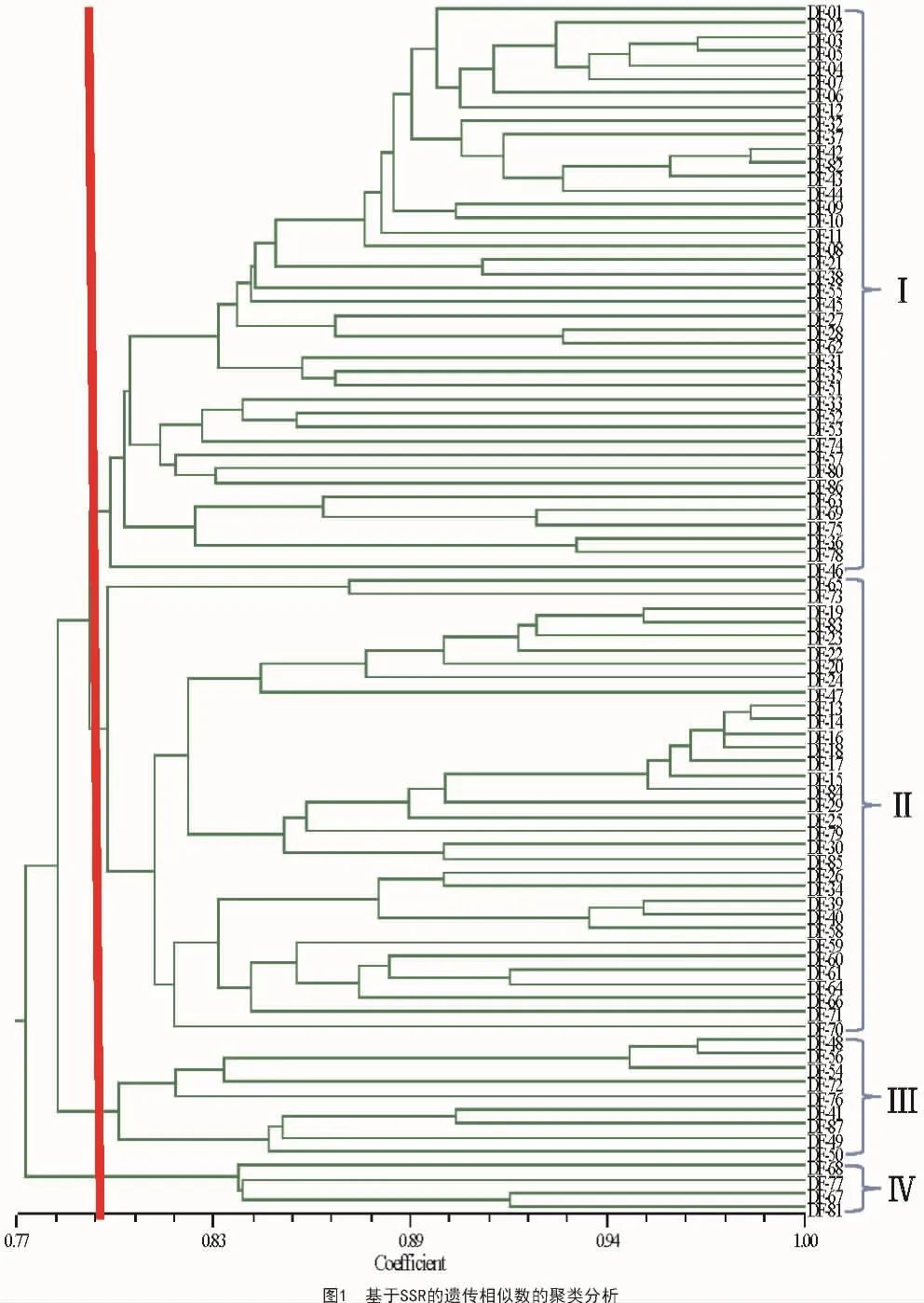
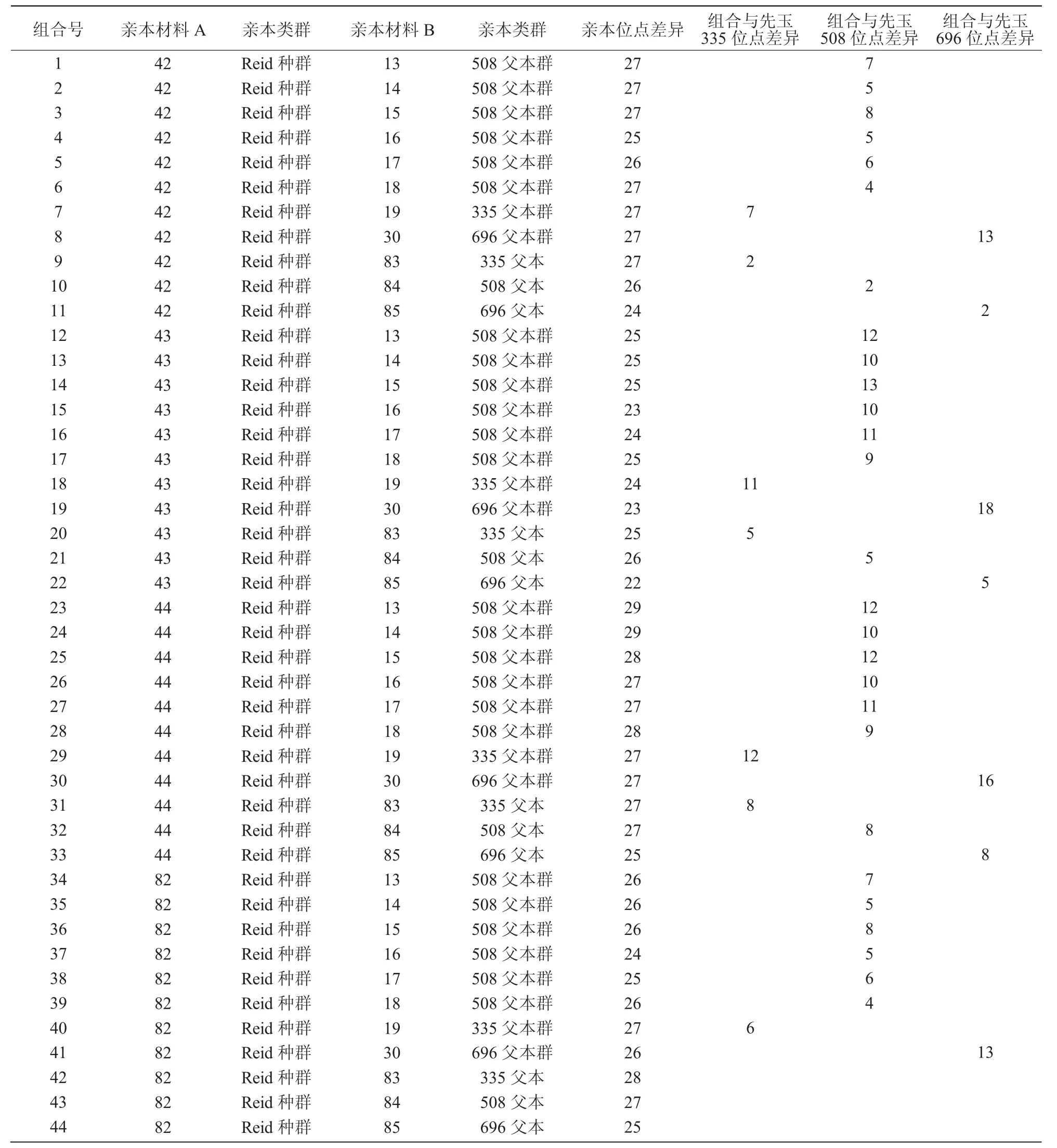
表3 玉米组合杂合位点分析
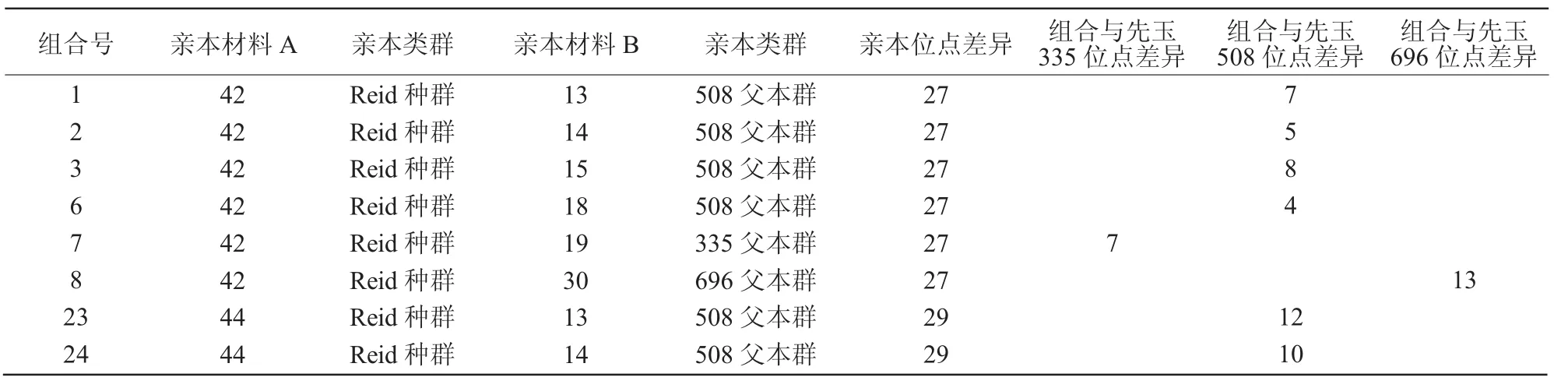
表4 玉米候选强优组合
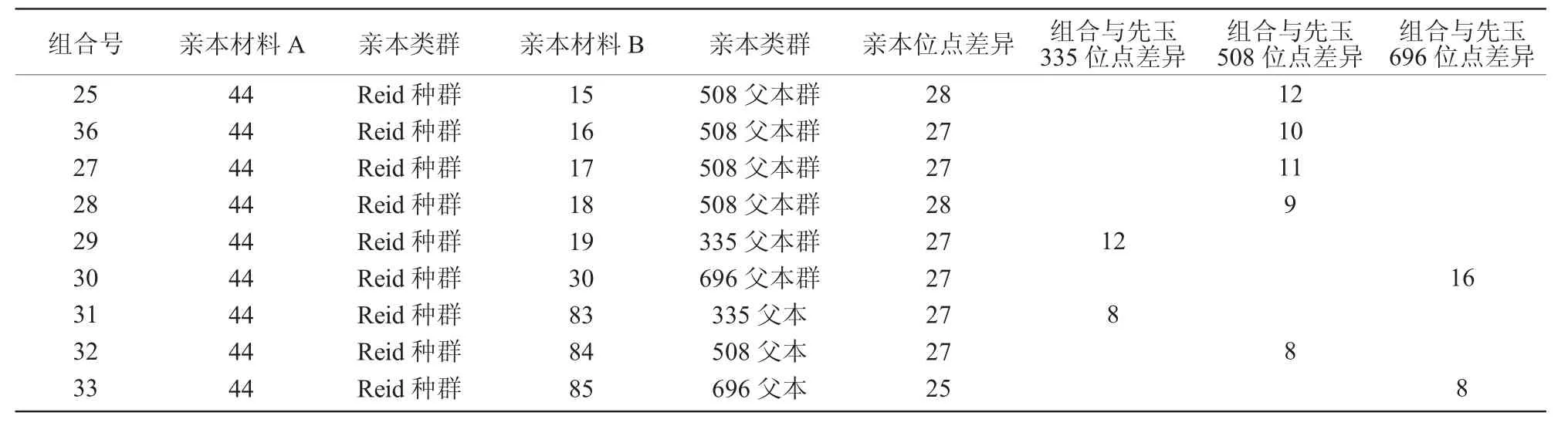
续表4
3 讨论
杂种优势利用是玉米育种的基础。为此,育种家们从实践中总结出了许多杂种优势类群与各种杂优模式。虽然,国内杂优类群按照标准种质划分为6大类[1],但近年来,由于先玉品种和郑单958在生产上的大面积推广,大家普遍接受了这2种杂优模式,很多育种家就是基于这2种模式进行大规模种质改良与组配杂交组合。因此,在育种中,主要按先玉母本群、先玉父本群、郑58群和昌7-2群划分杂优类群。本研究按这样思路将所有自交系进行了杂种优势类群的划分,使育种家们知道各自交系的归群及亲缘关系,指导他们有目标地按照杂优模式进行组配。本研究也显示还有一个新的类群,有可能是一个新的杂优类群,建议加强对它的研究。
按照杂优模式改良自交系,能方便应用成熟的杂交模式培育新品种,近年来审定的很多品种就是这样培育出来的,它们经过改良有了一些新的特性如抗性或适应性,又保持了原杂优模式的强杂种优势。但是,改良系很多,能组配的组合更多,如何产生有效的组合在实践中产生了困难。本研究比较了改良系与来源品种亲本的位点数,筛选出了有效亲本自交系。但将所有组合与对照品种的杂合位点数进行比较后发现,不是所有组合的杂合位点数等于或大于对照品种。研究表明,如果仅简单地按目标杂优类群进行组配是不够的,有必要进一步把组合与对照品种的杂合位点数进行比较后选择。本研究建议的杂优组合的分子预测方法在研究中已被证明。当然,推荐的组合要经过田间验证。
参考文献:
[1]李新海,傅骏骅,张世煌,等.利用SSR标记研究玉米自交系的遗传变异[J].中国农业科学,2003,33(2):1-9.
[2]XIE C,ZHANG S,LI M,et al.Inferring genome ancestry and estimatingmolecular relatedness among187 Chinese maize inbred lines[J].Journal ofGenetics and Genomics,2007,34(8):738-748.
[3]肖木辑,李明顺,李新海,等.黄淮海地区主要玉米自交系的SSR 遗传多样性分析[J].玉米科学,2008,16(2):1-7.
[4]聂永心,张丽,潘光堂,等.四川省常用玉米自交系SSR遗传多样性分析[J].分子植物育种,2005,3(1):43-51.
[5]郑淑云,王守才,刘东占.利用SSR标记划分玉米自交系杂种优势群的研究[J].玉米科学,2006,14(5):26-29.
[6]李锐,白建荣,程宇坤,等.山西省审定玉米品种亲本自交系的遗传多样性及杂优类群分析[J].作物杂志,2015(5):55-62.
[7]李锐,白建荣.山西中晚熟玉米区玉米自交系的遗传多样性及杂种优势群分析[J].山西农业科学,2013,41(4):311-316.
[8]赵久然,王凤格,郭景伦,等.中国玉米新品种DNA指纹库建立系列研究Ⅱ.适于玉米自交系和杂交种指纹图谱绘制的核心引物的确定[J].玉米科学,2003,11(2):3-5.
[9]陈发波,杨克诚,荣廷昭,等.西南及四川区试玉米组合遗传多样性分析[J].作物学报,2007,33(6):991-998.
[10]WANG F G,TIAN H L,ZHAO J R,et al.Development and characterization of a core set of SSR markers for fingerprinting analysis ofChinese maize varieties[J].Maydica,2011,56(1):7-17.
[11]吴毅歆.云南省222个玉米新品种DNA指纹图谱构建和遗传多样性分析[J].分子植物育种,2012,10(28):1199-1205.
[12]王凤格,田红丽,赵久然.中国328个玉米品种(组合)SSR标记遗传多样性分析[J].中国农业科学,2014,47(5):856-864.
[13]赵静,孙娟,张仁和,等.基于SSR技术分析陕西省玉米主栽品种的遗传多样[J].西北农业学报,2008,17(3):124-128.
[14]李俊芳,孙世贤.国家玉米主产区预试品种的SSR分析.Ⅱ.预试品种的遗传多样性[J].玉米科学,2007,15(1):16-20.
[15]葛建镕,刘晓鑫,易红梅,等.吉林省42份主推玉米杂交种的遗传分析[J].农业与技术,2008,28(5):37-42.
[16]高玉倩,田红丽,王凤格,等.吉林省玉米新品种SSR标准指纹数据库的构建及其分析[J].玉米科学,2012,20(3):43-47.
[17]朱志成.不同玉米品种SSR遗传多样性分析的研究[J].种子世界,2013(8):30-31.
[18]高永钢,高军,贾晋,等.内蒙古地区主要推广玉米品种遗传多样性研究[J].宁夏大学学报,2014,35(1):78-81.
[19]吴明生,贾希海,律宝春,等.杂交玉米种子纯度SSR技术鉴定结果的分析[J].种子科技,2006(6):47-48.
[20]薛艳颖,陈兴奎,樊妍,等.SSR分子标记技术在杂交玉米种子纯度鉴定中的应用[J].杂粮作物,2007,27(1):6-7.
[21]刘云婷,张秋兰,王倩,等.河北省玉米种子质量评价及遗传多样性分析[J].河北农业大学学报,2015,38(4):8-12.
猜你喜欢
蔬菜(2021年7期)2021-11-27 03:45:25
农业与技术(2021年20期)2021-11-04 08:10:00
种子(2021年3期)2021-04-12 01:42:22
农业科技与信息(2020年16期)2020-09-24 01:53:14
新农民(2019年11期)2019-09-16 08:38:30
中国奶牛(2019年2期)2019-03-08 08:10:08
外语教学理论与实践(2016年1期)2016-06-11 05:51:48
山西农经(2016年5期)2016-02-28 14:24:37
华东理工大学学报(自然科学版)(2014年1期)2014-02-27 13:48:29
华东理工大学学报(自然科学版)(2014年1期)2014-02-27 13:48:29
