
从生理生态角度解析毛竹爆发式生长的原因
2018-03-19 03:18王海湘温国胜梁谢恩黄皓南朱丽娜白尚斌张汝民
中南林业科技大学学报 2018年2期
徐 超,王海湘,温国胜,胡 策,梁谢恩,黄皓南,朱丽娜,白尚斌,张汝民
(浙江农林大学 林业与生物技术学院,浙江 临安 311300)
毛竹Phyllostachys edulis是隶属于禾本科刚竹属的一类植物。毛竹林被誉为“世界第二大森林”,是我国南方比较重要的森林资源,具有广泛的分布范围、较长栽培和利用历史以及较高的经济价值,在我国竹类资源里具有重要地位[1-2],其“爆发式生长”特性有别于其他类型的森林系统,成为我国森林固碳重要组成树种[3]。因此对于研究毛竹“爆发式生长”这一特定的生理时期具有重要意义。
毛竹林为“异龄林”,即具有典型的 “大小年”之分,大年主要完成竹笋的生长发育,小年主要完成竹鞭的生长以及更换竹叶[4]。毛竹迅速生长期是毛竹最重要生理阶段,指从春季竹笋出土、经历竹笋的高生长、竹枝抽出、竹叶的伸展直至高生长结束和秆形成的这个时间段。该时期跨度 40~60 d左右,竹笋就可完成其高生长,在此时期竹林生态系统的生物量快速增加。目前对于毛竹的研究主要集中在毛竹林生态系统生物量以及其潜在的固碳潜力[5-6]、各种胁迫对毛竹枝叶以及种子的影响[7-8]、竹林生态系统在叶片尺度上的光合固碳特性以及光能利用效率[9-12]、毛竹林生态系统的碳通量和水汽通量的变化特征[13-14]、毛竹林土壤微生物变化、各种酶活性、土壤呼吸、土壤理化性质以及土壤肥力[15-18]、毛竹林物质循环以及能量流动[19-20],以及不同经营方式对毛竹的生长和林地的影响等方面[21-22]。对于毛竹为什么会在短期内“爆发式生长”而产生如此神奇的生长速度?这一科学问题,至今仍未见报道。本文重点研究了毛竹快速生长期不同龄级毛竹的光合固碳能力、叶片水势变化的时空差异、以及地上器官生物量的分配时空格局,同时与小年期相应值比较,不仅可以从生理生态角度解释毛竹“爆发式生长”的原因,甚至可以评价现有经营模式的增汇潜力、研发毛竹林固碳增汇技术,为毛竹林生态经营管理方式创新提供科学依据。
1 材料和方法
1.1 试验地基本概况
试验地在位于浙江省杭州市临安市研口村(30°14′N,119°42′E)的毛竹林生理生态测量站内。此地气候类型属于亚热带季风。年均温约15.9 ℃,年降水量761~1 780 mm。土壤类型主要为黄壤与黄红壤,试验林是纯的毛竹林,林下植物很少,毛竹的竹龄分布主要在1~6 a,立竹度 2 400~3 000株/hm2,林相较好,大小年较为明显,经营措施为每年劈灌和小年采伐[12,23]。
1.2 试验方法
1.2.1 样地设置和采样
在立地条件基本一致的毛竹林设立3个样地,每一区域面积50 m ×50 m,分别统计此试验地毛竹的胸径,并分别统计2龄竹、4龄竹和6龄竹的数量,以便计算各龄竹的平均胸径,作为取样竹的标准即标准竹。根据幼竹的生长规律,将“爆发式生长期”分为生长初期(4月—5月)、生长中期(5月—6月)、生长后期(7月)3个阶段[13],并用SPAD-502、PAM-2100、PSYPRO多露点水势仪测定不同时期不同龄母竹的不同冠层(上层、中层、下层)且向阳外层叶片的相对叶绿素含量、光化学效率特征和水势。每次每样地随机选取2龄竹、4龄竹、6龄竹母竹标准竹各1株,共63株。
在试验地选取2龄竹、4龄竹、6龄竹的标准竹砍到,分叶、枝、秆3个器官称其鲜质量,各器官分别取2 000 g,在75℃下烘干至恒质量,测定其含水率,算出不同器官的干质量即生物量。
研究还分析了该试验林同一样地小年期母竹,为小年8月份采样和测定,方法同上。
1.2.2 数据处理
使用SPSS17.0进行相关数据分析,Sigma plot 12.5进行作图。
2 结果和分析
2.1 快速生长期内不同龄竹的光合的能力
2.1.1 快速生长期内不同龄竹相对叶绿素含量
不同生长阶段各龄竹不同冠层叶片的相对叶绿素含量见表1,同一生长阶段,相对叶绿素含量在不同龄毛竹叶片表现出相同的变化规律:相对叶绿素含量随着竹龄的增大而增加即6龄竹>4龄竹>2龄竹,此结果与朱弘等[12]结果完全相反,这可能与研究时间、叶龄不一致及 SPAD 值处于动态变化有关;而相对叶绿素含量在同龄竹不同生长阶段(初期,中期,后期)也具有一定变化规律:即为生长后期>生长前期>生长中期,而不同冠层表现为中层>下层>上层。
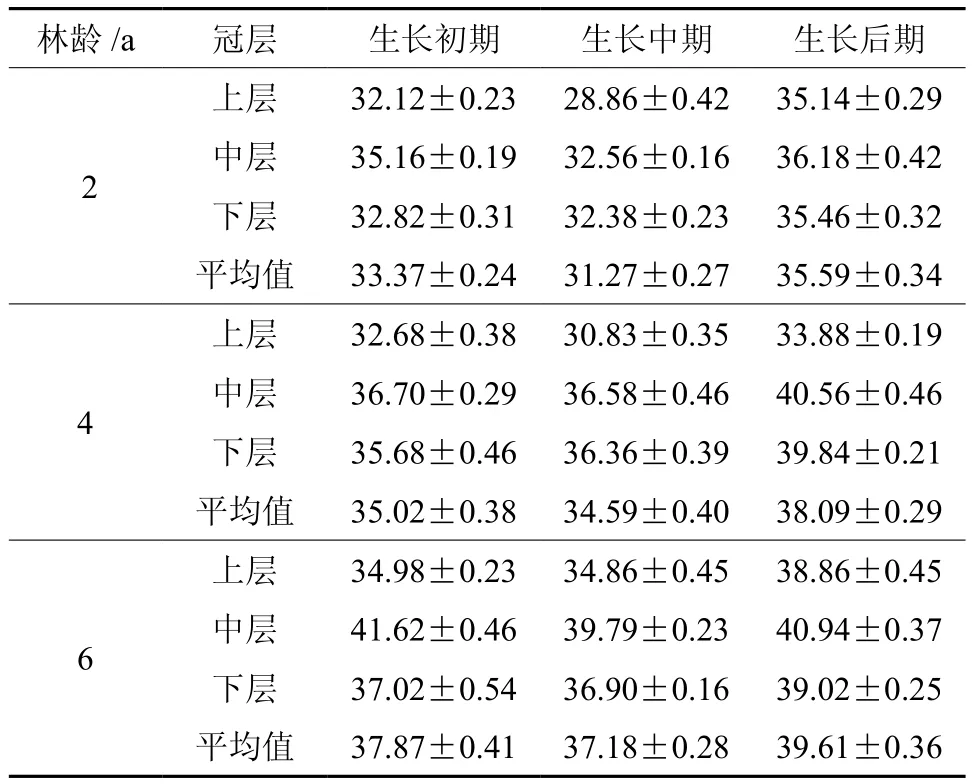
表 1 不同龄竹不同冠层毛竹叶片相对叶绿素含量†Table 1 Relative chlorophyll content in leaves of Moso bamboo from different layers at different ages
2.1.2 快速生长期与小年期的各龄竹的相对叶绿素含量比较
图1分析了快速生长期和小年期各龄级母竹相对叶绿素含量的差异。结果显示:不同竹龄毛竹叶片相对叶绿素含量值表现出很强的差异性,竹叶相对叶绿素含量值都随着竹龄的增大而增加,快速生长期母竹的相对叶绿素含量大于同期同龄小年的母竹,并且差异达到显著水平(P<0.05)。
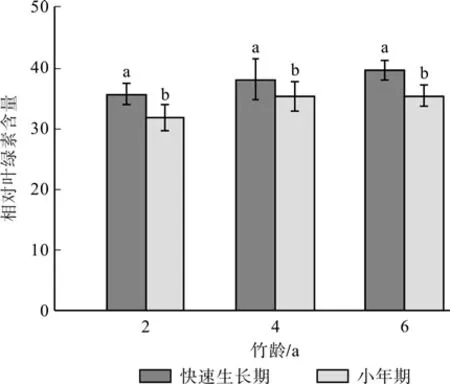
图1 各龄级毛竹相对叶绿素含量比较Fig.1 Comparison of relative chlorophyll content for Moso bamboo at different ages
2.1.3 快速生长期内不同龄竹的叶绿素荧光参数
不同生长阶段各龄竹叶绿素荧光参数见表2,不同生长阶段(初期、中期、后期),各竹龄PSⅡ实际光化学量子效率(actual photochemical ef fi ciency of PSⅡ,Yield)均表现为6龄竹>4龄竹>2龄竹,说明6龄竹PSⅡ的原初光能捕获效率高于2龄竹和4龄竹。与此同时,不同生长阶段6龄竹叶片的非光化学淬灭系数(non-photochemical quenching,NPQ)值均小于2龄竹和4龄竹即2龄竹>4龄竹>6龄竹,NPQ的值较小则表明叶片吸收的光能并没有大量的以热能的方式耗散掉。Yield和NPQ这两个参数表明在入射光照强度一定(相同生长阶段同一冠层同一方位)的条件下,竹龄越大利用光的能力越强,即6龄竹相对于2龄竹、4龄竹能更加高效的利用光能进行光合作用,初始荧光(minimal fl uorescence,Fo)和最大荧光(maximal fl uorescence,Fm)也都表现为6龄竹高于2龄和4龄竹,说明6龄竹电子传递较好;随着竹笋的高生长,各龄竹的PSⅡ潜在活性(potential activity,Fv/Fo)和PSⅡ最大光化学效率(maximal photochemical ef fi ciency of PSⅡ,Fv/Fm)均表现为生长前期最小,到了中后期Fv/Fm和Fv/Fo显著升高,其中生长中期Fv/Fm和Fv/Fo最高,温国胜等[11]从叶片尺度研究毛竹的快速固碳表明,毛竹林虽然在生长中期会换叶,但由于生态补偿作用,为了自身更好的生长,叶片光的化学效率和单位面积的光合固碳能力反而增加。
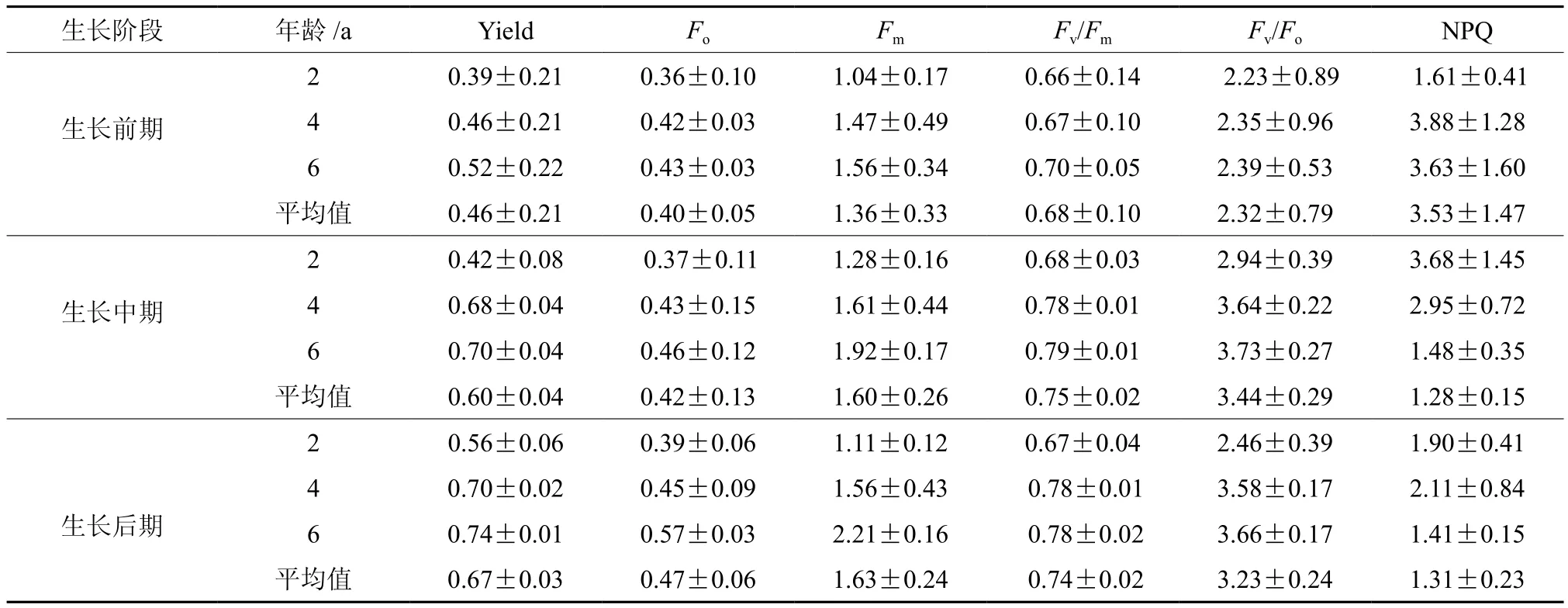
表 2 不同龄竹不同时期毛竹叶片叶绿素荧光参数Table 2 Chlorophyll fluorescence parameters of Moso bamboo at different ages in different growth stages
2.1.4 快速生长期与小年期的各龄竹叶绿素荧光参数比较
如图2快速生长期母竹叶绿素荧光参数与同期同样地小年期的相应值比较。结果表明:快速生长期2龄、4龄和6龄的Yield、Fo、Fm、Fv/Fo均大于同期同龄的小年值,但NPQ远远小于小年期。
2.2 快速生长期内不同龄竹叶片的水势
2.2.1 快速生长期内不同龄竹叶片的水势
不同生长阶段各龄竹不同冠层叶片的水势见表3。不同竹龄之间(2龄、4龄、6龄),在快速生长期内的叶水势均为竹龄越大水势越高,水分的输送和传输能力越强即6龄竹>4龄竹>2龄竹,从纵向水分传输上看,在快速生长期内不同林冠层(上层、中层、下层)的的叶水势均为下层水势>中层水势>上层水势。
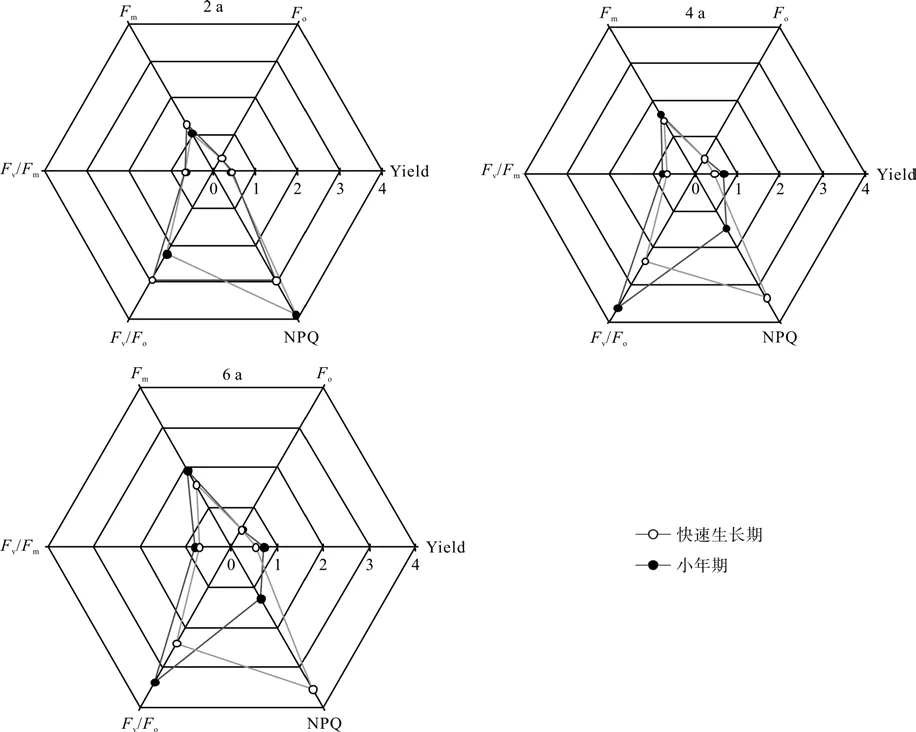
图2 各龄级毛竹叶绿素荧光参数的比较Fig.2 Comparison of chlorophyll fl uorescence parameters for Moso bamboo at different ages
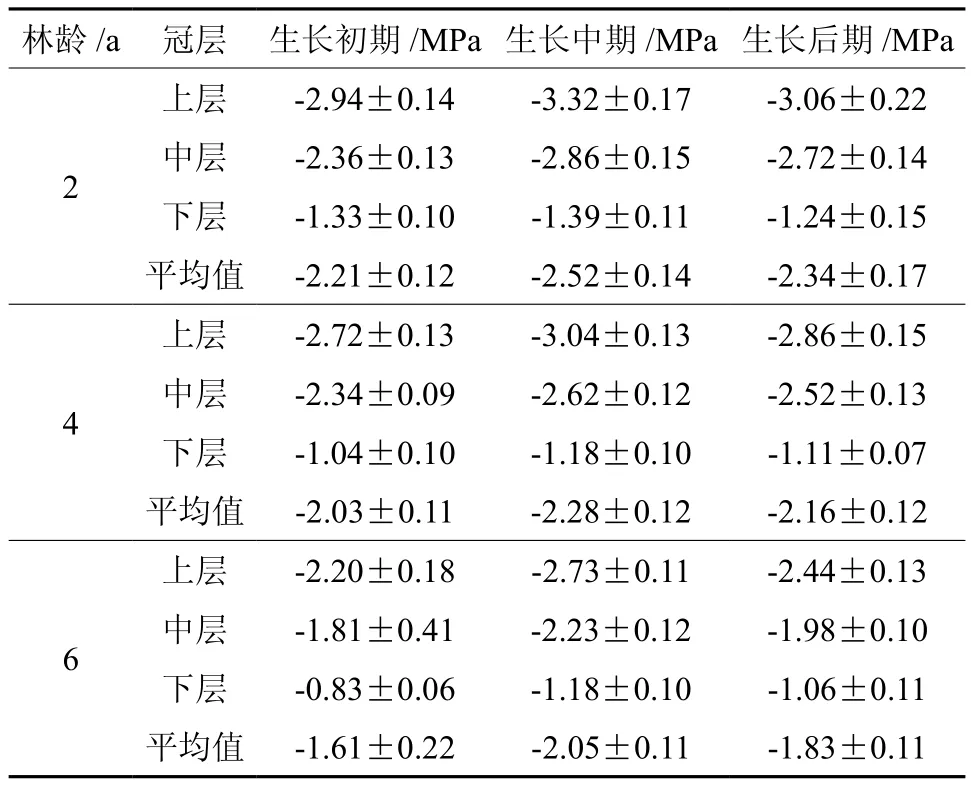
表 3 各龄竹不同时期毛竹叶片的水势Table 3 Leaf water potential of Moso bamboo at different ages in different growth stages
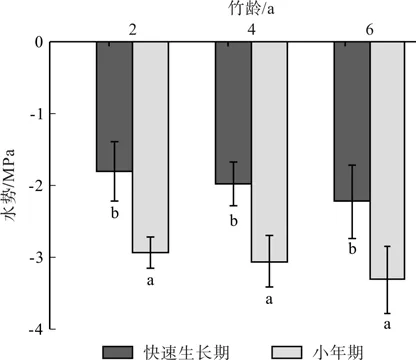
图3 各龄级毛竹叶水势的比较Fig.3 Comparison of leaf water potential for Moso bamboo at different ages
2.2.2 快速生长期与小年期的各竹龄叶片水势的比较
如图3快速生长期各龄竹的母竹叶片水势与同期同样地小年期的相应值比较。结果表明:快速生长期各龄竹的母竹的叶片水势均大于同期同样地小年期值,且差异达道显著水平(p<0.05)。
2.3 快速生长期不同龄竹地上器官的生物量
2.3.1 快速生长期内不同龄地上器官生物量的占比
不同生长阶段各龄竹地上器官生物量的占比见表4,在快速生长期内,叶片生物量占比范围是40.4%~69.4%,随着竹龄的增大,叶片生物量占比有变小趋势即6龄竹>4龄竹>2龄竹,同一竹龄在不同生长阶段叶生物量占比表现为生长后期>生长初期>生长中期;竹枝生物量占比变化范围为64.0%~75.3%,总体表现为:6龄竹>4龄竹>2龄竹,同一竹龄在不同生长阶段叶生物量占比表现为生长后期>生长中期>生长前期;竹秆生物量占比变化范围为57.2%~73.2%,总体表现为:6龄竹>4龄竹>2龄竹,同一竹龄在不同生长阶段叶生物量占比表现为生长后期>生长初期>生长后期,随竹龄的增长而渐渐增加。
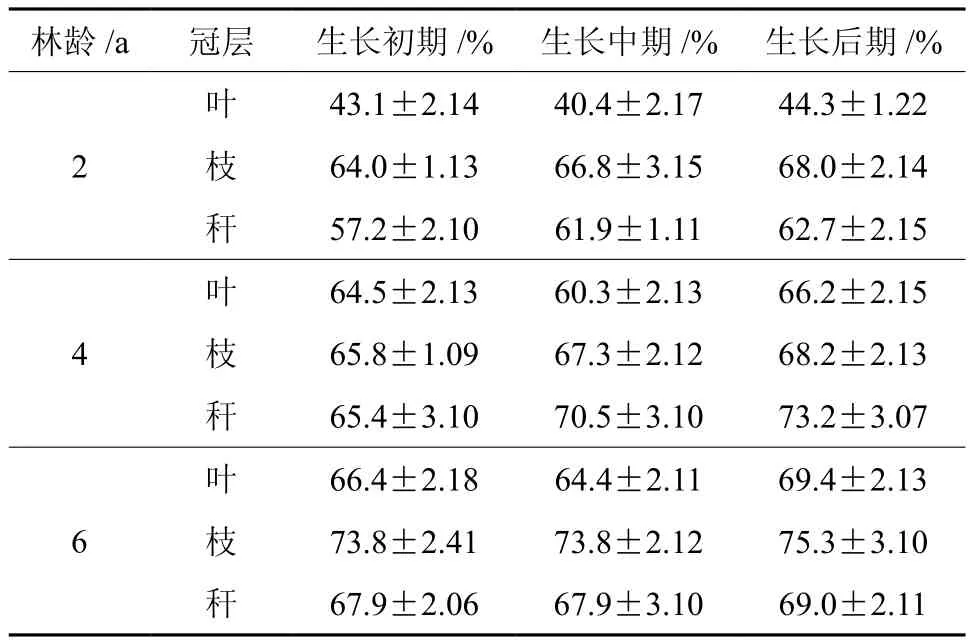
表 4 不同龄竹不同时期地上器官的生物量占比†Table 4 Percentage of aboveground biomass of Moso bamboo at different ages in different growth stages
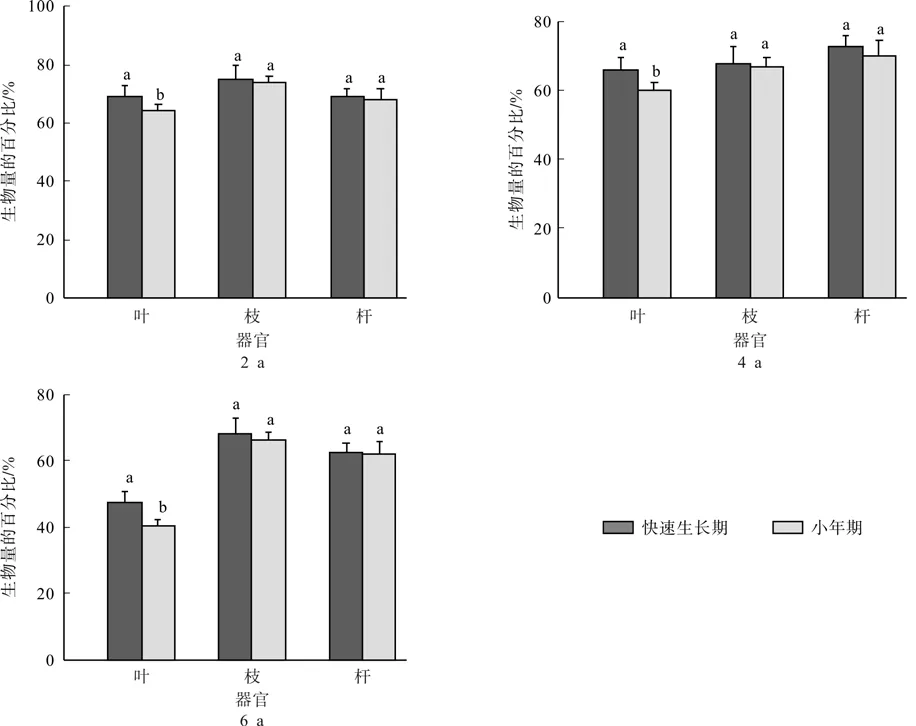
图4 各龄级毛竹地上器官生物量占比的比较Fig.4 Comparison of percentage of aboveground biomass for Moso bamboo at different ages
2.3.2 快速生长期与小年期的各竹龄地上器官生物量百分比的比较
如图4所示,快速生长期各龄竹地上器官生物量占比与同期同样地小年期的相应值比较,结果表明:快速生长期各龄竹地上器官生物量占比均大于同样地同期的小年期,并且叶片的的生物量占比与小年期相比达到显著差异(p<0.05)。
3 结论和讨论
绿色植物利用叶绿素等光合色素通过一系列的化学反应,固定有机物,叶绿素含量的多少不仅影响植物的光合作用,同时也是植物光合固定有机物能力一个重要指标[11]。毛竹相对叶绿素含量在同龄竹不同生长阶段(初期,中期,后期)表现为:即为生长后期>生长前期>生长中期。生长中期各龄级毛竹叶片的相对叶绿素含量有降低趋势,林琼影等[24]和孙少波等[25]研究表明生长中期(5—6月份)毛竹林处于更新换叶的时段,老的叶片即将凋落,叶片中叶绿素快速降解,含量急剧下降,所以此时期相对叶绿素含量最低;生长中期以后,随着毛竹叶片逐渐成熟,叶绿素迅速合成,其相对含量也渐渐增加,到生长后期毛竹完成换叶,相对叶绿素含量又逐渐升高。如图1快速生长期母竹的相对叶绿素含量大于同期同龄小年的母竹。并且差异达到显著水平(p<0.05)。这说明快速生长期各母竹叶片与小年期相比具有光合固定有机物的能力。如图2快速生长期2龄、4龄和6龄的Yield、Fo、Fm、Fv/Fo均大于同期同龄的小年值,但NPQ远远小于小年期,此结果与刘琳等[26]研究一致。这说明快速生长期各母竹与小年期相比利用光能的效率更大,光合作用更明显,光能以热能的形式被耗散掉的较少。
水势是指水的化学势,是推动水在生物体内移动的势能。水在土壤-植物-大气连续体中总是从水势较高处向水势较低处移动[27]。本文各龄竹在快速生长期内水势强弱表现为6龄竹>4龄竹>2龄竹,袁佳丽研究表明[23]在快速生长期的不同阶段(初期、中期、后期),水分的传输方向都是高龄竹将水分传输给低龄竹,与此文研究结果一致。如图3快速生长期各龄竹的母竹的叶片水势均大于同期同样地小年期值,且差异达道显著水平(p<0.05)。说明快速生长期竹笋的生长对母竹冠层叶片水势产生重要影响,此时期母竹水势较强,母竹和子竹之间有较高的水势差,有像幼竹传输更多的水分的能力。
生物量是森林生态系统生产力的最好的指标[28]。枝、秆生物量占比随着在不同龄竹之间变现为:6龄竹>4龄竹>2龄竹,不同生长阶段表现为生长后期>生长中期>生长前期。而叶的不同生长阶段表现为生长后期>生长前期>生长中期,研究结果与徐超等[13]研究一致,即毛竹林是异龄林,有大小年之分,生长中期毛竹林由于是换叶期,叶片发黄脱落,生物量转移。因此此时期毛竹叶片生物量占比小于前期和后期。如图4快速生长期各龄竹地上器官生物量占比均大于同样地同期的小年期,并且叶片的的生物量占比与小年期相比达到显著差异(p<0.05)。说明快速生长期竹笋的生长对母竹地上生物量格局产生重要影响,尤其母竹叶片影响最大,与小年期相比,快速生长期叶片进行光合作用积累有机物的能力大大增加,生物量分配更大,产生更多的有机物以供自身和子竹的生长发育。
综上所述,幼竹的快速生长依赖于母竹,即母竹对子竹的快速生长有促进作用。幼竹的快速生长影响了母竹叶片光合产物的分配,母竹分配光合产物给予子竹,使竹笋在短期内迅速完成高生长。在快速生长初期,母竹的相对叶绿素含量,而此时幼竹的快速生长需要较大的能量,因为在竹林管理中,此时可以在此时段合理地补充土壤氮、磷、钾养分,以维持幼竹的快速生长。在快速生长阶段,叶片各冠层的水势均大于同期的小年期,依靠水势,母竹将水分源源不断地供给新竹,以保证新竹健康快速生长。因此,水势成为了驱动毛竹快速生长的关键因素之一。幼竹快速生长期,母竹在各器官生物量分配上,竹叶所占比例提升,为满足新竹生长,在生物量分配上进行了调整与优化。竹叶生物量分配比例大于6龄竹,在竹林抚育时,应对2~4龄竹的管理加强,及时砍伐老竹。
毛竹之所以快速生长,除了母竹的促进作用外,也与自身的特定基因有关,对于控制毛竹快速生长的关键启动因子是什么?有待以后作进一步研究。本文从母竹在快速生长期和小年期叶片相对叶绿素含量,水势动态规律来看,快速生长期母竹叶片相对叶绿素含量和水势都小于同期的小年期,这与植物的不同生理期有关,同时还可能受到外界环境不同,如降雨、施肥、采伐等有密切关系,具体的动态变化规律还需要进一步研究。
[1]窦 营,余学军,岩松文代.中国竹子资源的开发利用现状与发展对策[J].中国农业资源与区划,2011,32(5):65-70.
[2]江泽慧.世界竹藤[M].沈阳:辽宁科学技术出版社,2002.
[3]李 翀,周国模,施拥军,等.毛竹林老竹水平和经营措施对新竹发育质量的影响[J].生态学报,2016,36(8):2243-2254.
[4]许庆标.春笋幼竹期毛竹林养分特征研究[D].北京:中国林业科学研究院,2013.
[5]周国模,刘恩斌,施拥军,等.基于最小尺度的浙江省毛竹生物量精确估算[J].林业科学,2011,47(1):1-5.
[6]肖复明,范少辉,汪思龙,等.毛竹Phyllostachy pubescens、杉木Cunninghamia lanceolata人工林生态系统碳贮量及其分配特征[J].生态学报,2007, 27(7): 2794-2801.
[7]朱强根,金爱武,唐世刚,等.毛竹枝叶生物量的冠层分布对钩梢和施肥的响应[J].中南林业科技大学学报,2015,35(1):24-29.
[8]陈俊任,柳 丹,吴家森,等.重金属胁迫对毛竹种子萌发及其富集效应的影响[J].生态学报,2014,34(22):6501-6509.
[9]施建敏,郭起荣,杨光耀.毛竹光合动态研究[J].林业科学研究,2005,18(5): 551-555.
[10]Xu Y, Wong M, Yang J, et al. Dynamics of Carbon accumulation during the fast growth period of bamboo plant[J]. The Botanical Review, 2011, 77(3): 287-295.
[11]Wen G, Zhang L, Zhang R, et al. Temporal and spatial dynamics of Carbon fi xation by MOSO bamboo (phyllostachys pubescens)in subtropical China[J]. Botanical Review, 2011, 77(3): 271-277.
[12]朱 弘,温国胜.不同竹龄毛竹冠层叶片PSⅡ叶绿素荧光特性的比较[J].中南林业科技大学学报,2017,37(1):12-19.
[13]徐 超,温国胜,王海湘,等.毛竹快速生长期的高生长与碳通量的变化规律[J].东北林业大学学报,2016,44(11):1-4.
[14]刘玉莉,江 洪,周国模,等.安吉毛竹林水汽通量变化特征及其与环境因子的关系[J].生态学报,2014,34(17):4900-4909.
[15]徐秋芳,姜培坤.毛竹竹根区土壤微生物数量与酶活性研究[J].林业科学研究,2001,14(6):648-652.
[16]夏根清,李国栋,周 燕,等.不同人工经营毛竹林土壤理化性质变化[J].竹子研究汇刊,2012,31(3):38-43.
[17]王奇赞.应用PCR-DGGE方法研究毛竹土壤细菌群落结构及其遗传多样性[D].临安:浙江林学院,2009.
[18]高志勤,傅懋毅.经营方式对毛竹林土壤肥力指数的影响[J].南京林业大学学报(自然科学版),2008,32(4):81-85.
[19]郭宝华.不同生产力水平毛竹林碳氮磷生态化学计量特征研究[D].北京:中国林业科学研究院,2014.
[20]Tang X, Xia M, Guan F, et al. Spatial distribution of soil Nitrogen, Phosphorus and Potassium stocks in MOSO bamboo forests in subtropical China[J]. Forests, 2016, 7(11): 267.
[21]余 林,李万和,徐海宁,等.江西安福不同类型毛竹林地土壤微生物量碳特征研究[J].经济林研究,2017,35(1):80-85.
[22]孟 勇,艾文胜,杨 明,等.上阔下竹复合经营模式对毛竹生长的影响[J].经济林研究,2016,34(3):135-141.
[23]袁佳丽,温国胜,张明如,等.毛竹快速生长期的水势变化特征[J].浙江农林大学学报,2015,32(5):722-728.
[24]林琼影,胡 剑,温国胜,等.天目山毛竹叶冬季光合作用日变化规律[J].福建林学院学报,2008,28(1):61-64.
[25]孙少波,杜华强,李平衡,等.基于小波变换的毛竹叶片净光合速率高光谱遥感反演[J].应用生态学报,2016,27(1):49-58.
[26]刘 琳.毛竹快速生长期茎秆反射光谱及叶绿素荧光特性研究[D].临安:浙江农林大学,2012.
[27]During H, Loveys B R. Diurnal changes in water relations and abscisic acid in field grown Vitis vinifera cvs. I. Leaf water potential components and leaf conductance under humid temperate and semiarid conditions[J]. VITIS-Journal of Grapevine Research, 2016, 21(3): 223.
[28]Yen T. Culm height development, biomass accumulation and Carbon storage in an initial growth stage for a fast-growing moso bamboo (Phyllostachy pubescens)[J]. Botanical Studies, 2016,57(1): 1-9.
猜你喜欢
天津农林科技(2022年2期)2022-04-19
干旱地区农业研究(2021年6期)2021-12-08
疯狂英语·新阅版(2021年7期)2021-08-27
农村实用技术(2020年9期)2020-11-19
浙江农林大学学报(2020年5期)2020-11-05
东坡赤壁诗词(2020年2期)2020-06-04
教育家(2020年14期)2020-04-26
湖北林业科技(2019年4期)2019-09-16
幼儿画刊(2018年1期)2018-01-04
农业与技术(2017年22期)2017-12-29
