
间伐对华南地区相思人工林土壤理化性质、微生物及酶活性的影响
2018-03-19 03:18刘颂颂陈葵仙沈德才苏纯兰卢琳超
中南林业科技大学学报 2018年2期
刘颂颂,陈葵仙,沈德才,张 浩,苏纯兰,卢琳超
(1. 东莞市林业科学研究所,广东 东莞 523106;2. 香港高等科技教育学院,中国 香港特别行政区 999077)
林木的生长与光照、能量、土壤养分和水分等环境因子联系密切,其中土壤作为森林生态系统的重要组分,是林木生长发育的基础[1]。森林土壤理化性质是反映森林土壤肥力的重要指标[2],在一定程度上影响着森林健康生长,其生产力水平直接决定着森林的生物产量和功能的发挥[3-5]。因此,植被和土壤是生态系统的重要组成部分。20世纪80年代后期,广东省在改造针叶林树种中种植了大面积相思林。这些相思树种具有生长快、耐干旱贫瘠和生产量高等特点,是广东省荒山造林绿化的优良先锋树种。但由于树种结构单一、大面积纯林会造成林地地力衰退和生产力下降,严重制约了人工林的可持续生产和发展。许多研究结果表明,长期经营纯林会引起林地土壤理化性质变差,土壤肥力下降,土壤质量严重退化[6-8]。间伐作为森林经营的一种手段,其通过调整林分密度结构,减小林木间竞争,从而促进林下植被和林木的更新与生长,提高林分的物种多样性,改变土壤肥力[9-13]。
土壤酶活性和土壤微生物是土壤生态系统的重要组成成分,在有机质分解、养分循环和植物营养等过程中发挥着关键作用,其数量变化与土壤的通气性、水分状况、养分状况及有机质含量等土壤理化指标密切相关,其活性可以在一定程度上反映土壤肥力[14-20]。目前,国内外相关研究大多集中在不同间伐强度对林木生长[13]、林下植被多样性[10,20]、林地土壤理化性质[21]等的影响,而间伐后对林地的土壤理化性质、土壤微生物和酶活性的影响研究仍较少。本研究比较研究东莞大岭山马占相思林不同间伐套种模式对林下土壤理化性质、土壤微生物和土壤酶活性的变化,揭示间伐套种与相思林土壤肥力的关系,为华南地区相思林林地的维持提供理论与实践依据。
1 试验地概况
东莞市林业科学园(新园)地处大岭山林场,该林场位于广东东莞虎门镇、长安镇和大岭山镇交界处(22°39′~ 23°09′ N,113°31′~114°15′ E),属南亚热带季风海洋气候,全年温暖多雨,年平均气温22.1 ℃,最热月(7月)平均温度28.2 ℃,极端最高温37.9 ℃,最冷月(1月)平均气温13.4 ℃,极端最低气温低于0 ℃;年降水量1 500~2 400 mm,降水集中在4—9月,降水量占全年的80%以上,并以台风雨居多。试验地为低丘陵,海拔约67 m,土壤为花岗岩发育而成的低丘赤红壤,有机质含量较低。地表植被以人工林为主,主要有马尾松Pinus massoniana、杉木Cunninghamia lanceolata、尾叶桉Eucalyptus urophylla及相思类树种。本研究所用马占相思人工林种植时间为1996年,种植密度2 500 株/hm2(株行距2 m × 2 m),林下植被主要有鸭脚木Schef fl era octophilla、毛叶冬青Ilex pubilimba、三叉苦Evodia lepta、乌毛蕨Blechnum orientale和九节Psychotria rubra等。
2004年选择立地条件相同的马占相思纯林进行疏伐试验,间伐改造区M1、M2、M3分别表示未间伐+套种、间伐30%+套种树种和间伐60%+套种树种;以未间伐的相思人工林为对照(CK);各个区的特征如表1所示。
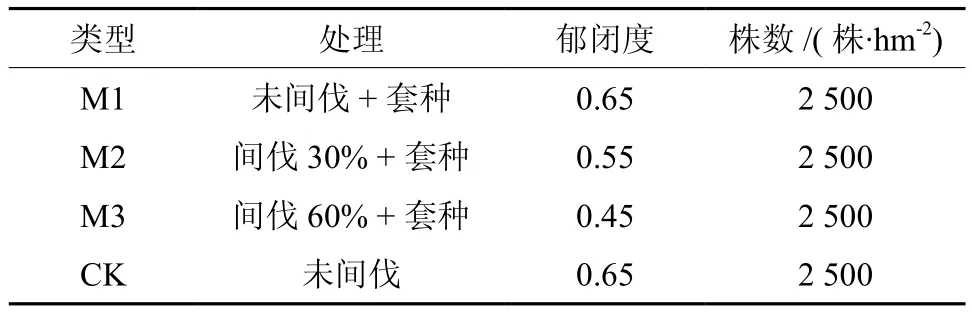
表 1 马占相思林分改造类型的基本特征Table 1 Basic modes characteristics of Acacia mangiumin plantation
2 研究方法
2.1 土壤采集
2016年1月(旱季)和7月(雨季),在每个林分20 m×20 m样方内,随机选取3处代表性20 m的样带,分别在0、10和20 m处取表层土壤0~10 cm采样,充分混合取土样,其中部分鲜土带回实验室后置于4 ℃保存供土壤酶活性与土壤微生物的测定;其余土样经过除去石块、根系等杂物后,磨碎过筛,装袋待测定土壤理化性质等。
2.2 土壤理化性质的测定
土壤自然含水率采用铝盒烘干法测定;pH采用水∶土=2.5∶1玻璃电极法;全氮含量采用半微量凯氏定氮法测定;铵态氮采用苯酚——次氯酸盐测定;硝态氮采用酚二磺酸比色法测定;全磷含量采用NaOH熔融——钼锑抗比色法测定;全钾含量采用氢氧化钠熔融-火焰原子吸收分光光度法;碱解氮含量采用1.0 mmol/L NaOH 碱解扩散法测定;速效磷含量采用0.5 mmol/L NaHCO3浸提——钼锑比色法;速效钾含量采用NH4OAC浸提——火焰光度法测定;交换性钙和交换性镁含量采用乙酸铵提取-火焰原子吸收分光光度法;交换性钠含量采用NH4OAC-NH4OH浸提——火焰光度法测定;具体测定方法参见参考文献[23]。
2.3 土壤微生物的测定
土壤微生物生物量碳和微生物生物量氮采用氯仿熏蒸浸提法测定。MBC和MBN(mg/kg)计算方法如下: MBC= 2.22 EC,MBN= 2.22 EN,式中,EC、EN分别为熏蒸和未熏蒸土样浸提液的有机碳、全氮的差值;2.22为校正系数。
2.4 土壤酶活性的测定
以容量法测定过氧化氢酶活性,以30 min后1 g土壤的0.1 N(即0.02 mol/L)高锰酸钾的毫升数表示;靛酚蓝比色法测定脲酶活性,以24 h后1 g土壤中NH3-N的毫克数表示;磷酸苯二钠比色法测定磷酸酶活性,以24 h后1 g土壤中消耗五氧化二磷的毫克数表示[24]。
2.5 数据处理与分析
采用模糊隶属函数法对不同改造模式土壤改良作用能力进行综合评定。X(u)=(X-Xmin)/(Xmax-Xmin);式中:X(u)为某一改造模式在某一土壤因子的隶属函数值;X为该植物在某一土壤因子的平均测定值;Xmax和Xmin分别为所有土壤因子在该改造模式下平均测定值中的最大值和最小值。
试验数据用Excel 2010进行初步处理,通过SPSS19.0软件的单因素方差分析(ANOVA)比较检验;以土壤理化性质、土壤微生物和土壤酶活性指标通过CANOCO4.5软件完成PCA分析。
3 结果与分析
3.1 土壤化学性质比较
3.1.1 土壤pH值和土壤电导率
试验结果显示,无论是旱季,还是雨季,林分土壤pH值均呈酸性。旱季时,土壤pH值以M3林分最高,M2和CK林分次之,M1林分最低;与旱季相比,雨季各林分土壤pH值呈增加趋势,以M3林分最高,M1林分次之,CK林分最低(表2)。旱季时,3种改造模式的土壤电导率显著高于CK林分,其土壤电导率在110.55 ~124.07 uS/cm;与旱季相比,雨季时改造对林分的土壤电导率呈显著下降趋势(表2)。

表 2 不同改造模式和季节对土壤pH值和土壤电导率的比较†Table 2 Comparison of pH and soil electrical conductivity in different thinning modes and seasons
3.1.2 土壤化学性质
3.1.2.1 全 N、NH4+-N和NO3--N
由表4可知,旱季时,M2林分的土壤全N含量显著高于其它林分和CK林分;与旱季相比,雨季时各林分的土壤全N含量显著增加,土壤全N含量在2.04~2.75 g/kg。旱季时,各林分的土壤NH4-N在16.10~26.57 mg/kg之间,但在雨季时M1、M2和CK林分的土壤NH4-N含量显著下降,M3林分含量变化不显著。各林分在旱季时土壤NO3-N含量在9.35~24.43 mg/kg之间;与旱季相比,改造显著促进林分的土壤NO3-N含量的增加,但未间伐CK林分含量变化不显著。
3.1.2.2 全P和有效P含量
季节的变化对土壤P含量影响呈显著影响(表3)。旱季时,林分全P含量以CK最高,为7.90 mg/kg,3种改造模式林分土壤全P含量在6.72~7.69 mg/kg之间。与旱季相比,雨季时各林分土壤全P含量显著低于旱季时,其含量在0.17~0.23 mg/kg之间。各林分在旱季时土壤有效P含量较低,为0.90~1.42 mg/kg之间;雨季促进了林分的土壤有效P含量的增加,以M3林分的土壤有效P含量最高。
3.1.2.3 交换性元素含量
季节的变化对土壤速效K、交换性Mg和交换性Na含量影响呈显著影响(表4)。与旱季相比,雨季时林分的土壤速效K、交换性Mg和交换性Na含量均低于旱季;而土壤交换性Ca含量在不同季节和林分变化不一致。旱季时,改造降低了林分的土壤速效钾含量、交换性Ca含量和交换性Mg含量,均表现出显著低于CK林分,交换性Na含量以M1和M2林分较高。雨季时,林分土壤速效K、交换性Mg和交换性Na含量以M3林分较高,交换性Ca含量以M2林分最高(87.13 mg/kg)。
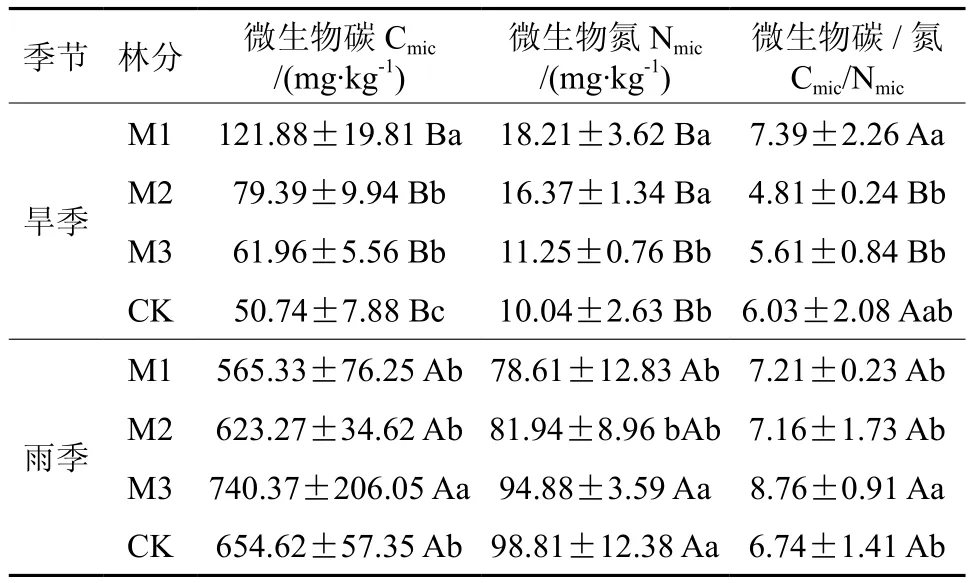
表 4 不同改造模式和季节对土壤微生物含量的比较†Table 4 Comparison of soil microorganism biomass in different thinning modes and seasons
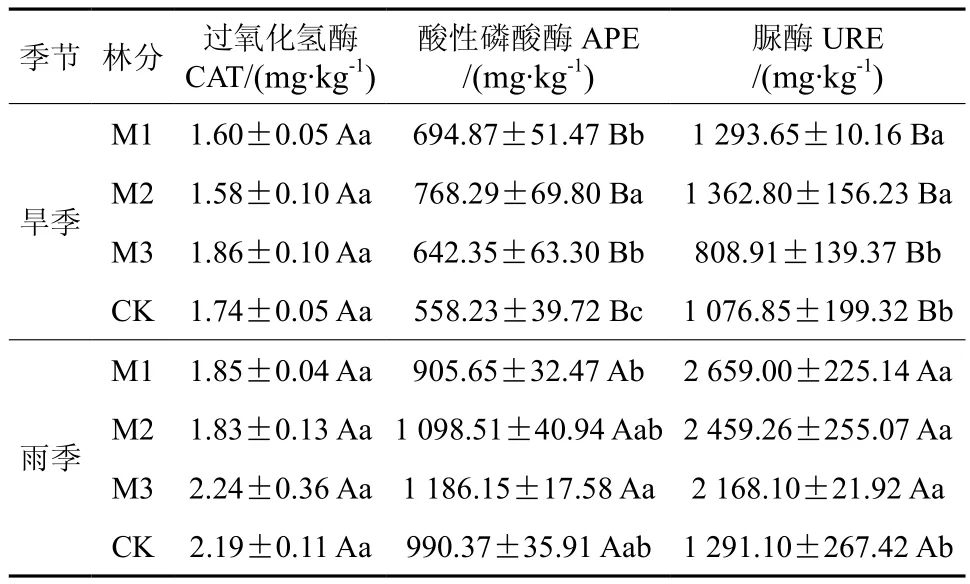
表 5 不同改造模式和不同季节对土壤3种酶活性的比较Table 5 Comparison of three soil enzyme activities in different thinning modes and seasons
3.2 土壤微生物碳和氮含量比较
由表4可以看出,雨季林分的土壤Cmic和Nmic含量显著高于旱季,反映了雨季时林分微生物较为活跃。旱季时,Cmic和Nmic含量以M1林分最高,分别为121.88 mg/kg和18.21 mg/kg,M2林分次之,CK林分最低;Cmic/Nmic比在4.81~7.39之间,以M1林分最高。雨季时,林分的土壤Cmic含量在565.33~740.37 mg/kg之间,土壤Nmic含量在78.61~98.81 mg/kg之间,Cmic/Nmic比值在6.74~8.76之间;M3林分土壤Cmic和微生物Cmic/Nmic均以M3林分最高,但微生物Nmic以CK最高。
3.3 土壤酶活性的比较
试验结果显示,季节对土壤过氧化氢酶活性影响不显著,对土壤酸性磷酸酶(APE)和脲酶(URE)影响显著(表5)。与旱季相比,雨季林分的CAT酶活性稍高于旱季。旱季时,M2林分的土壤APE酶活性(768.29 mg/kg)显著高于其它林分和CK,M1和M2林分的土壤URE酶活性(1 293.65~1 362.80 mg/kg)显著高于M3和CK。雨季, 土壤APE酶活性以M3林分最高,1 186.15 mg/kg,M2林分次之;改造促进林分土壤APE酶活性,均显著高于CK林分(1 291.10 mg/kg)。
3.4 土壤理化性质与土壤微生物、酶活性PCA分析
将旱季和雨季林分的土壤理化性质、土壤微生物和酶活性指标进行主成分分析(PCA),揭示了不同改造模式样点的分布。在旱季时,第一和第二主成分的累积贡献率为81.9%,其中PC1占总方差的69.3%,PC2占方差的12.6%(图1)。PC1主要反映的是pH值、全P、CAT酶、NH4-N、交换性Na、URE酶和全N因子,PC2主要反映的是电导率、APE酶、有效P、交换Mg、交换性Ca和速效K因子。第一排序(横)与Cmic、Nmic、交换性Na、APE酶、电导率、pH值、全P、CAT酶、Cmic/Nmic、有效P和速效K因子呈正相关,与其它因子间呈负相关;第二排序(纵)轴 与 NH4-N、NO3-N、URE酶、 全 N、Cmic、Nmic、APE酶和电导率因子呈正相关,与其它因子呈负相关。PCA将3种改造模式和CK林分明显分开,各个林分的3个样地均能各自聚为一类,但各林分的响应因子存在显著差异。
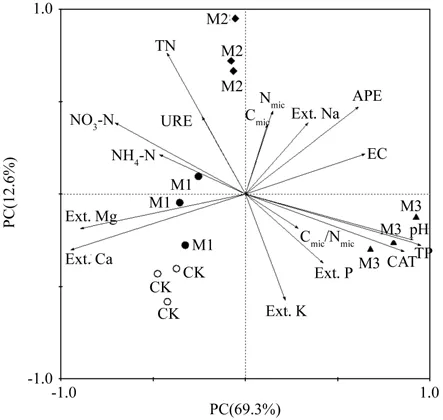
图1 不同改造模式旱季的土壤理化性质与土壤微生物、酶活性PCA分析Fig. 1 PCA biplot of soil physical and chemical characteristics,soil microorganism and soil enzyme activities in the dry season
在雨季时,第一和第二主成分的累积贡献率为64.6%(图2)。PC1主要反映的是pH值、电导率、全N、有效P、交换性Na、CAT酶、Cmic和Nmic因子,PC2主要反映的是NO3-N、速效K、全P、NH4-N、交换性Mg和Cmic/Nmic比因子。第一排序(横)与Cmic/Nmic比、全P、NH4-N、URE酶、pH值、交换性Ca和NO3-N呈正相关,与其它因子间呈负相关;第二排序(纵)轴与Cmic、Nmic、APE酶、交换性Mg、速效K、Cmic/Nmic比、全P、NH4-N和URE酶呈正相关,与其它因子呈负相关。Cmic、Nmic、交换性Na、APE酶、电导率、CAT酶、Cmic/Nmic比、有效P和速效K因子由图还可以看出,3种改造模式样地与CK林分样点分布差异较大,M1、M2和M3林分样地均能较好各聚为一类,但CK林分3个样地零星分布。
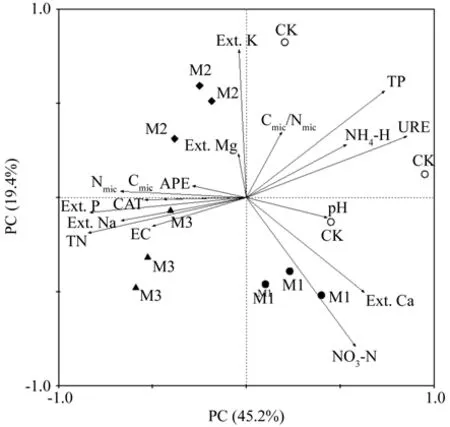
图2 不同改造模式在雨季时土壤理化性质与土壤微生物、酶活性PCA分析Fig. 2 PCA biplot of soil physical and chemical characteristics,soil microorganism and soil enzyme activities in the wet season
3.5 综合评价分析
基于土壤理化性质、土壤微生物和土壤酶活性因子进行的模糊隶函数值综合分析,比较不同改造模式在不同季节对林分土壤改良作用。结果表明,无论在旱季还是雨季,改造模式均能有效地改善林分土壤肥力,以M1改造模式(不间伐+套种)效果最佳,M2改造模式次之,M3改造模式较低(表6),反映了改造相思林的方式是可行的。
4 结论与讨论
4.1 间伐对相思林的土壤理化性质影响
不同改造模式后,林分由于林分郁闭度、物种多样性等有所不同,其土壤的理化性质差异明显。本研究发现,改造后林分的土壤全N含量差异不大,这可能是由于间伐套种刺激了土壤微生物的活性[25],加快了土壤有机氮的分解使其含量降低。同时,间伐后套种树种的生长消耗了土壤中大量的无机氮,因此土壤全氮含量没有显著增加[26]。NH4-N和NO3-N含量在不同季节差异显著,雨季时的土壤NH4-N和NO3-N含量高于旱季,这可能是因为在雨季时上层土壤中有机质含量较高,氮营养元素来源丰富缘故。全P 和有效 P 在旱季和雨季的差异不显著,表明了相思林整个林分均较为缺乏,这主要与华南地区土壤类型有关[27]。土壤速效 K 、交换性Mg和交换性Na含量在不同林型及不同季节变化较大,旱季时改造林分的速效K 、交换性Mg和交换性Na含量均显著高于雨季,这可能与林下营养物质分解速度慢及季节有关。
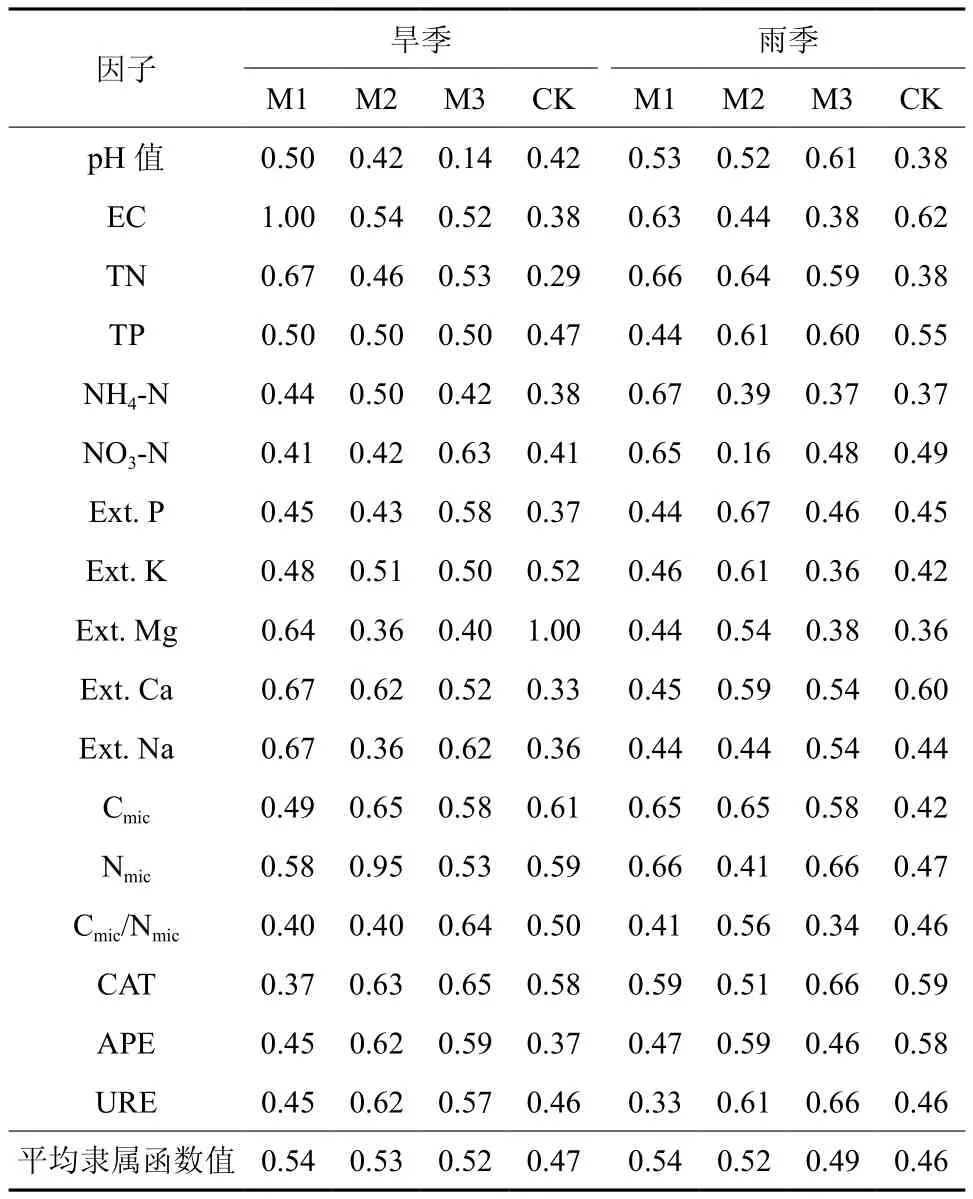
表 6 基于模糊隶属函数值对林分不同改造模式的综合评价Table 6 Comprehensive assessments of different modes based on subordinate function values
4.2 间伐对相思林的土壤微生物与土壤酶活性的影响
土壤微生物量的季节变化受森林凋落量节律的影响[28],然而季节的变化则主要是通过温度和水分等环境条件对土壤微生物过程产生了影响[29-30]。本研究结果初步显示,不同间伐改造林分的土壤微生物量碳和微生物生物量氮表现为雨季(夏季)较高,而旱季(冬季)较低,这与许多学者的研究结果相近[31-36]等人的研究结果较为相近,反映了雨季时的林分条件更利于土壤微生物的生长。本研究结果还显示,无论是雨季还是旱季,土壤微生物生物量均以M1林分最高。其可能原因未改造林分植物根系与土壤微生物共生关系受干扰较小,其土壤微生物活性和数量则明显增加,促进土壤有机质的矿化,从而反映了间伐套种模式可促进土壤微生物活跃度。
土壤酶是土壤中的生物催化剂,对维持土壤生态系统的稳定起着重要作用[33-34]。本研究结果表明,林分间伐套种11 a后,林分物种多样性呈增加趋势,林分的土层也形成大量的植物根系;改造后林分的土壤酶活性高于未间伐林分。这与林分植物根系代谢释放了大量的酶类物质有关,因为这些酶类物质活跃了土壤酶活性[37-38]。不同间伐套种改造模式在不同季节对土壤酶活性影响显著;旱季时土壤酸性磷酸酶和脲酶较低,雨季时土壤酶活性显著增加,与前人的研究结果基本一致[11]。杨宁等[39]和刘作云等[40]研究结果揭示林分的土壤酶活性的季节变化与林分中地被覆盖物的数量有关。因此,雨季时林分地被植物丰富度显著高于旱季,导致土层根系繁茂,土壤含水量较高,有利于土壤微生物的迁移与促进酶类物质的运动与活性 。
4.3 间伐对相思林的土壤因子的关系探讨
PCA分析结果表明,土壤理化性质、土壤微生物生物量和酶活性在旱季和雨季之间相关性显著,反映了不同间伐改造模式对林分土壤肥力、养分和能力转化有关,可以作为评价林分土壤质量的指标。本研究结果表明,3种间伐改造模式对土壤理化性质、微生物生物量和酶活性产生了积极影响,但在旱季和雨季的变化规律不同。未间伐与套种模式林分在旱季和雨季均与土壤NO3-N和交换性Ca相关;在旱季时,30%间伐林分与土壤全N、微生物C、微生物N和脲酶相关,60%间伐林分与土壤pH值、过氧化氢酶、全P 和微生物C/N比相关;在雨季时,前者与土壤速效K和交换性Mg,后者与土壤电导率、全N、有效P、微生物C、微生物N、交换性Na和磷酸酶相关。土壤理化性质、微生物和酶活性在不同季节变化反映了间伐改造影响着林分土壤质量,以未间伐与套种模式林分对土壤因子影响较为稳定。通过隶属函数值分析结果表明,无论是雨季,还是旱季,以未间伐与套种改造模式对同龄马占相思纯林的效果最佳。该研究结果将为相思林的土壤改良提供有价值的信息。
[1]Vieira S R, Pierre L H, Grego C R, et al. A geostatistical analysis of rubber tree growth characteristics and soil physical attributes[J]. Quantitative Geology and Geostatistics, 2010, 16:255-264.
[2]林培松,尚志海.韩江流域典型区不同森林类型土壤理化性质初步研究[J]. 水土保持研究, 2009, 16(3): 117-120.
[3]Rocha G N, Goncalves J L M, Moura I M. Changes in soil fertility and growth of an Eucalyptus grandis plantation fertilized with biosolid[J]. New Forests, 2004, 28(4):623-639.
[4]Souza J P D, Araújo G M, Haridasan M. In fl uence of soil fertility on the distribution of tree species in a deciduous forest in the Triêngulo Mineiro region of Brazil[J]. Plant Ecology, 2007,191(2): 253-263.
[5]姜 勇. 森林生态系统微量元素循环及其影响因素[J]. 应用生态学报, 2009, 20(1): 197-204.
[6]韩艺师,魏彦昌,欧阳志云,等. 连栽措施对桉树人工林结构及持水性能的影响[J]. 生态学报, 2008, 28(9): 4609-4617.
[7]唐万鹏,李吉跃,漆良华,等. 江汉平原连栽杨树人工林对土壤化学性质的影响[J]. 中南林业科技大学学报, 2009, 29(5):72 -76.
[8]魏 强,凌 雷,柴春山,等. 甘肃兴隆山森林演替过程中的土壤理化性质[J]. 生态学报, 2012, 32(15): 4700-4713.
[9]张鼎华, 叶章发, 范必有, 等. 抚育问伐对人工林土壤肥力的影响[J]. 应用生态学报, 2001, 12 (5): 672-676.
[10]Taki H, Inoue T, Tanaka H, et al. Responses of community structure, diversity, and abundance of understory plants and insect assemblages to thinning in plantations[J]. Forest Ecology and Management, 2010, 259(3): 607- 613.
[11]赵朝辉,方 晰,田大伦,等. 间伐对杉木林林下地被物生物量及土壤理化性质的影响[J]. 中南林业科技大学学报,2012,32(5): 102-107.
[12]Forrester D I, Medhurst J L, Wood M, et al. Growth and physiological responses to silviculture for producing solidwood products from Eucalyptus plantations: An Australian perspective[J]. Forest Ecology and Management, 2010, 259(9):1819- 1835.
[13]Saunders M, Tobin B, Black K, et al. Thinning effects on the net ecosystem exchange of a Sitka spruce forest are temperaturedependent[J]. Agricultural and Forest Meteorology, 2012, 157: 1-10.
[14]Steenwerth K L, Jackson L E, Calderon F J, et al. Soil microbial community composition and land use history in cultivated and grassland ecosystems of coastal California[J]. Soil Biology and Biochemistry, 2002, 34(1): 1599-1611.
[15]王金成,李海燕,梁 健.黄土高原子午岭不同林龄天然油松林土壤微生物组成研究[J]. 水土保持通报,2010,30(1):43-48.
[16]刘 曼,辛 颖,赵雨森. 氯氟氰菊酯对水源涵养林土壤微生物及酶活性的影响[J].东北林业大学学报, 2013,41(6):80-83.
[17]Li F, Liu M, Li Z, et al. Changes in soil microbial biomass and functional diversity with a nitrogen gradient in soil columns[J].Applied soil ecology, 2013, 64: 1-6.
[18]张雯雯,徐 军,董丰收,等. 苄嘧磺隆对水稻田土壤微生物群落功能多样性的影响[J]. 农业环境科学学报, 2014, 33(9):1749-1754.
[19]Timothy M, Veronica AM, Francisco C, et al. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensivelymanaged agricultural landscape[J]. Soil Biology & Biochemistry,2014, 68: 252-262.
[20]马履一,李春义, 王希群,等. 不同强度间伐对北京山区油松生长及其林下植物多样性的影响[J]. 林业科学, 2007, 43(5):1-9.
[21]杨媛媛,黎建强,陈奇伯,等. 滇中高原常绿阔叶林土壤生物学特性对土壤理化性质的影响[J]. 生态环境学报, 2016,25(3): 393-401.
[22]Tian D L, Peng Y Y, Yan W D, et al. Effects of thinning and litter fall removal on fi ne root production and soil organic carbon content in Masson pine plantations[J]. Pedosphere, 2010, 20(4):486-493.
[23]鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000.
[24]关松荫. 土壤酶及其研究法[M]. 北京: 北京农业出版社,1986.
[25]Garcia F O , Rice C W. Microbial biomass dynamics in tall grass prairie[J]. Soil Sci. Soc. Am. J., 1994, 58(3):816-823.
[26]魏金明,姜 勇,符明明,等. 水、肥添加对内蒙古典型草原土壤碳、氮、磷及pH的影响[J]. 生态学杂志, 2011, 30(8):1642-1646.
[27]李志安,翁 轰,余作岳. 人工植被对广东热带沿海侵蚀地土壤之影响[J]. 热带地理, 1996, 16(4): 310-318.
[28]吴艺雪,杨效东,余广彬. 两种热带雨林土壤微生物生物量碳季节动态及其影响因素[J].生态环境学报,2009,18(2):658-663.
[29]Moore-Kucera J, Dick R P. PLFA profiling of microbial community structure and seasonal shifts in soil of a Douglas- fi r chronosequence[J]. Microbial Ecology, 2008, 55(3): 500-511.
[30]何 斌,刘运华,陆志科,等. 肉桂人工林土壤速效养分与酶活性的季节变化[J]. 经济林研究, 2004, 22 (3): 1-4.
[31]王卫霞,史作民,罗 达,等. 南亚热带3 种人工林土壤微生物生物量和微生物群落结构特征[J].应用生态学报, 2013,24(7): 1784-1792.
[32]Bini D, Santas C A D, Bouillet J P, et al. Eucalyptus grandis and Acacia mangium in monoculture and intercropped plantations:Evolution of soil and litter microbial and chemical attributes during early stages of plant development[J]. Applied Soil Ecology, 2013, 63: 57-66.
[33]罗 达,史作民,唐敬超,等. 南亚热带乡土树种人工纯林及混交林土壤微生物群落结构[J]. 应用生态学报, 2014, 25(9):2543-2550.
[34]Hoogmoed M, Cunningham S C, Baker P, et al. N-fixing trees in restoration plantings: effects on nitrogen supply and soil microbial communities[J]. Soil Biology and Biochemistry,2014,77: 203-212.
[35]余 林,李万和,徐海宁,等.江西安福不同类型毛竹林地土壤微生物量碳特征研究[J]. 经济林研究, 2017, 35(1): 80-85.
[36]陈 瑕,杨 宁. 亚热带红壤丘陵区5种人工林对土壤性质的影响[J]. 西北农林科技大学学报(自然科学版), 2013, 41(12):167-173.
[37]付美云,杨 宁,杨满元,等. 衡阳紫色土丘陵坡地不同恢复阶段土壤微生物与养分的耦合关系[J]. 生态环境学报, 2015,24(1): 41-48.
[38]Bell T H, Klironomos J N, Henry H A L. Seasonal response of extracellular enzyme activity and microbial biomass to warming and nitrogen addition[J]. Soil Science Society of America Journal, 2010, 74(3): 820-828.
[39]杨 宁,杨满元,雷玉兰,等. 紫色土丘陵坡地土壤微生物群落的季节变化[J]. 生态环境学报, 2015, 24(1): 34-40.
[40]刘作云,杨 宁. 衡阳紫色土丘陵坡地不同植被恢复阶段土壤化学与微生物性质[J]. 生态环境学报, 2015, 24(4): 595-601.
猜你喜欢
今日农业(2022年16期)2022-11-09
今日农业(2022年14期)2022-09-15
河北果树(2021年4期)2021-12-02
中南林业科技大学学报(2021年3期)2021-03-23
——以杉木抚育间伐为例
绿色科技(2020年21期)2021-01-08
电脑迷(2020年5期)2020-12-18
防护林科技(2020年6期)2020-08-12
森林与环境学报(2020年4期)2020-07-27
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
