
豫东不同树龄梨园土壤微生物生态特征
2018-03-14 11:24马丽
河南农业科学 2018年1期
马 丽
(商丘师范学院生物与食品学院植物与微生物互作重点实验室,河南商丘476000)
土壤是生态系统不可或缺的重要组成部分,在植物生长发育过程中,不仅作为营养库供给营养,还具有涵养雨水、支撑以及稳定和缓冲环境变化等作用[1]。土壤微生物是土壤生态系统中最活跃的组分,其种类多、数量大,参与生物固氮、土壤有机质的分解和腐殖质的形成等过程,对土壤有机质的矿化分解和土壤养分的转化起着非常重要的作用[2-4]。果园土壤微生物的种类和数量在一定程度上反映了果园土壤肥力状况,与果树的生长及产量的形成有一定的关系,而不同树龄果园的根系空间分布不同[5-6],土壤有机碳储量[7]、土壤理化性状[8]、水肥营养状况[9-10]也不同,进而土壤微生物的分布也受到影响。因此,研究不同树龄果园土壤微生物状况有利于对不同树龄果园进行合理施肥及耕作管理,进而提高果树产量和品质。目前,对于果园土壤微生物的研究多集中于耕作措施[11-12]、水肥管理[13-14]、覆盖模式[15]等对土壤微生物的影响,且研究品种多为苹果[16]、葡萄[17]、脐橙[18]、蜜橘[19]等。梨是我国栽培历史悠久、种植面积大、产量多的主要果树之一,也是国内外栽培的大宗果树之一,而目前对不同树龄梨园土壤微生物和酶活性的研究鲜见报道。鉴于此,以豫东地区宁陵万亩梨园为研究对象,调查了不同树龄梨园不同土层深度土壤微生物包括细菌、真菌、放线菌数量分布特征,研究了土壤脲酶、蔗糖酶、过氧化氢酶活性空间变化,以期为评估不同树龄梨园的土壤质量以及指导不同树龄梨园的合理管理提供理论依据。
1 材料和方法
1.1 试验区概况
试验区位于河南省商丘市宁陵县石桥乡万亩梨园(34°45'N,115°32'E),该区地处豫东平原,年平均气温14.2℃,无霜期216 d,年平均降雨量720 mm。土壤类型以砂壤土为主,梨树品种以酥梨为主。
1.2 研究方法
选择树龄为 3、5、10、22、53、89 a 的梨园为研究对象,梨树特征见表1。在结果末期每个树龄的梨园选取3个样点,选择树体大小、树势基本一致的代表性梨树,沿2个行间方向选取距离树干100 cm处设点采样,每点选取3个土层深度(0~20、20~40、40~60 cm),每个样点2个行间方向同层土样采用四分法进行均匀混合,装入塑封保鲜袋,放入冰盒带回实验室,剔除土壤杂物后,将土样分为2份,1份新鲜土样立即测定土壤微生物数量,另1份风干后研磨过筛,用于测定土壤酶活性。
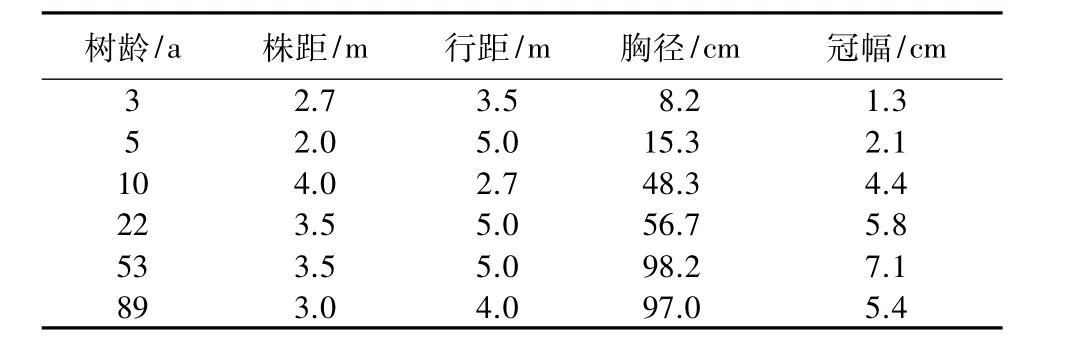
表1 不同树龄梨园梨树特征
1.3 指标测定
土壤蔗糖酶活性采用3,5–二硝基水杨酸比色法进行测定,用37℃条件下培养24 h后每克土壤中所含葡萄糖的质量表示;脲酶活性采用次氯酸钠-苯酚钠比色法进行测定,用37℃条件下培养24 h后每克土壤中所含NH3-N的质量表示;过氧化氢酶活性采用高锰酸钾滴定法进行测定,用每克土壤20 min所消耗的 0.01 mol/L KMnO4的量表示[20]。土壤微生物包括细菌、真菌、放线菌的测定采用平板稀释计数法,烘干法测定土壤含水率,用每克干土中的微生物数量表示[21]。土壤有机质含量测定采用重铬酸钾氧化法[22]。
1.4 数据分析
采用Excel 2003进行数据处理,采用SPSS 17.0进行Duncan氏差异显著性检验和Pearson相关性分析。
2 结果与分析
2.1 不同树龄梨园土壤有机质特征
由表2可知,0~20 cm土层内,3 a和5 a梨园土壤有机质含量最高(分别为 10.16、10.13 g/kg),分别较其他各树龄梨园高 28.28%~82.08%、27.90%~81.54%,且差异显著,而 89 a梨园最低(5.58 g/kg),显著低于其他各树龄梨园;20~40 cm土层内,3、5、10 a梨园显著高于 22、53、89 a 梨园;40~60 cm 土层也表现为3 a梨园最高(6.11 g/kg),而89 a梨园最低(3.58 g/kg)。总体上,各土层土壤有机质含量随树龄的增加而降低。不同土层间相比较,各树龄梨园均表现为0~20 cm >20~40 cm>40~60 cm,且 0~20 cm土层显著高于20~40 cm土层,而40~60 cm土层土壤有机质含量最低,表明梨园土壤有机质含量随土层深度增加而降低。
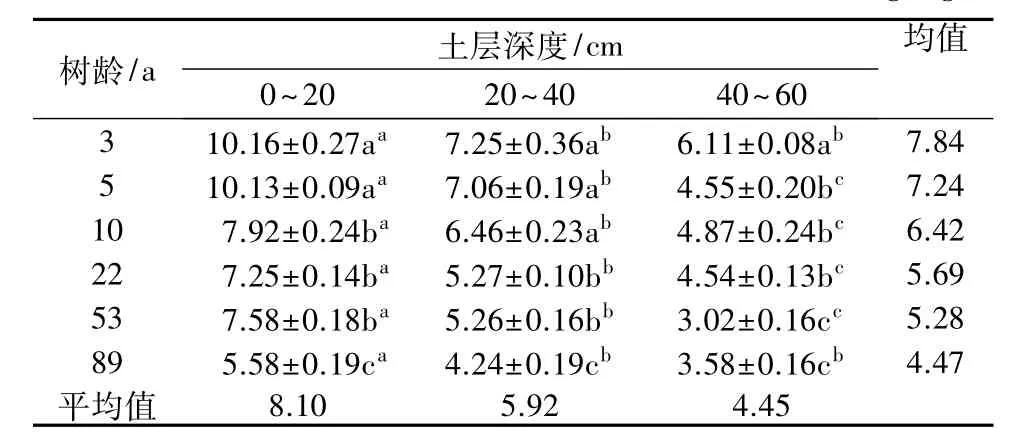
表2 不同树龄梨园土壤有机质含量变化 g/kg
2.2 不同树龄梨园土壤酶活性的变化
由表3可知,土壤蔗糖酶和脲酶活性均随土层深度增加而降低,而各土层间过氧化氢酶活性差异不显著。不同树龄梨园之间相比较,0~20 cm土层5 a梨园蔗糖酶活性最高,比其他梨园高5.73%~31.39%,20~40 cm和40~60 cm土层均以10 a梨园最高,分别比其他梨园高 7.10% ~30.69%和9.94% ~30.07%,蔗糖酶活性随树龄的增加呈先增加后降低趋势。总体上,土壤脲酶活性随梨园树龄的增加而逐渐降低。不同树龄梨园土壤过氧化氢酶活性差异不显著。
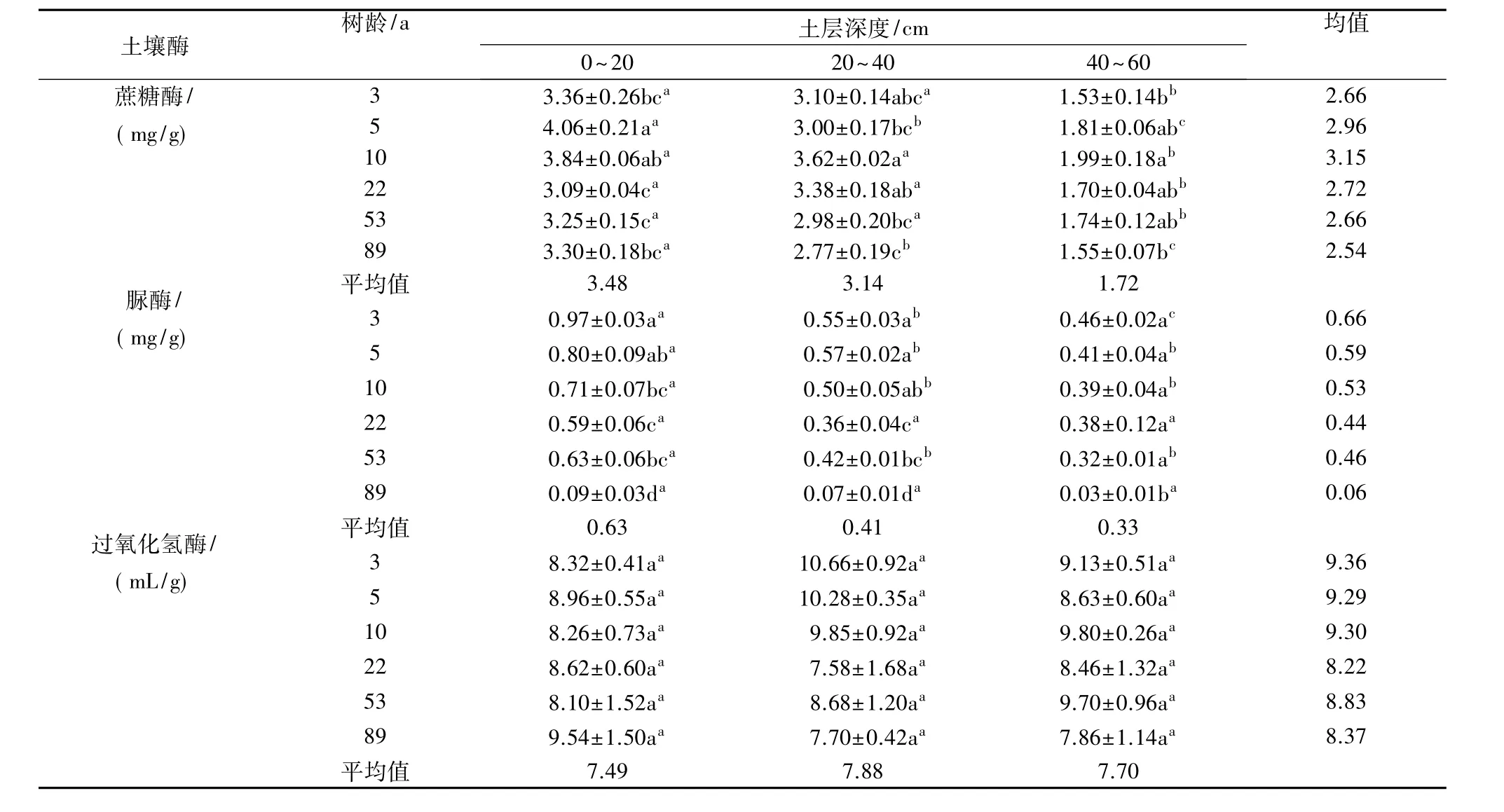
表3 不同树龄梨园不同土层土壤酶活性的变化
不同土层的土壤酶活性相比较,0~20 cm和20~40 cm土层蔗糖酶活性均显著高于40~60 cm土层,各树龄0~20 cm和20~40 cm土层平均土壤蔗糖酶活性分别比40~60 cm土层高102.52%和82.66%;土壤脲酶除22 a和89 a梨园,其他树龄梨园均表现为0~20 cm土层显著高于20~40 cm和40~60 cm土层,各树龄0~20 cm土层平均土壤脲酶活性分别比20~40 cm和40~60 cm土层高53.44%和90.45%,表明土壤蔗糖酶和脲酶活性随土层深度的增加而降低。过氧化氢酶活性在各树龄梨园不同土层间差异均未达到显著水平,表明0~60 cm土层内过氧化氢酶受土层深度的影响变化不大。
2.3 不同树龄梨园土壤微生物的数量特征
2.3.1 不同树龄梨园土壤微生物的数量分布 由表 4 可知,3、5、10、22、53、89 a 梨园在试验研究深度0~60 cm的土层范围内,土壤细菌数量为3.26×107~33.96×107cfu/g,细菌平均数量随树龄的增加逐渐减少,不同土层土壤细菌数量也大致表现出随树龄的增加而逐渐减少的趋势。其中,0~20 cm土层各树龄间差异均达到显著水平;20~40 cm土层3 a和5 a梨园显著高于其他树龄梨园,分别为32.79×107cfu/g和24.56×107cfu/g;40~60 cm 土层3 a 梨园细菌数量最高,为 22.37×107cfu/g,89 a梨园最低,为 3.26×107cfu/g,且均与其他梨园差异显著。不同土层细菌数量比较,除53 a和89 a梨园,其他各梨园均随土层深度的增加而减少,且0~20 cm土层与40~60 cm土层均达到差异显著水平;0~20、20~40、40~60 cm 土层各梨园细菌平均值 分 别 为 19.69× 107、17.76× 107、13.13×107cfu/g。
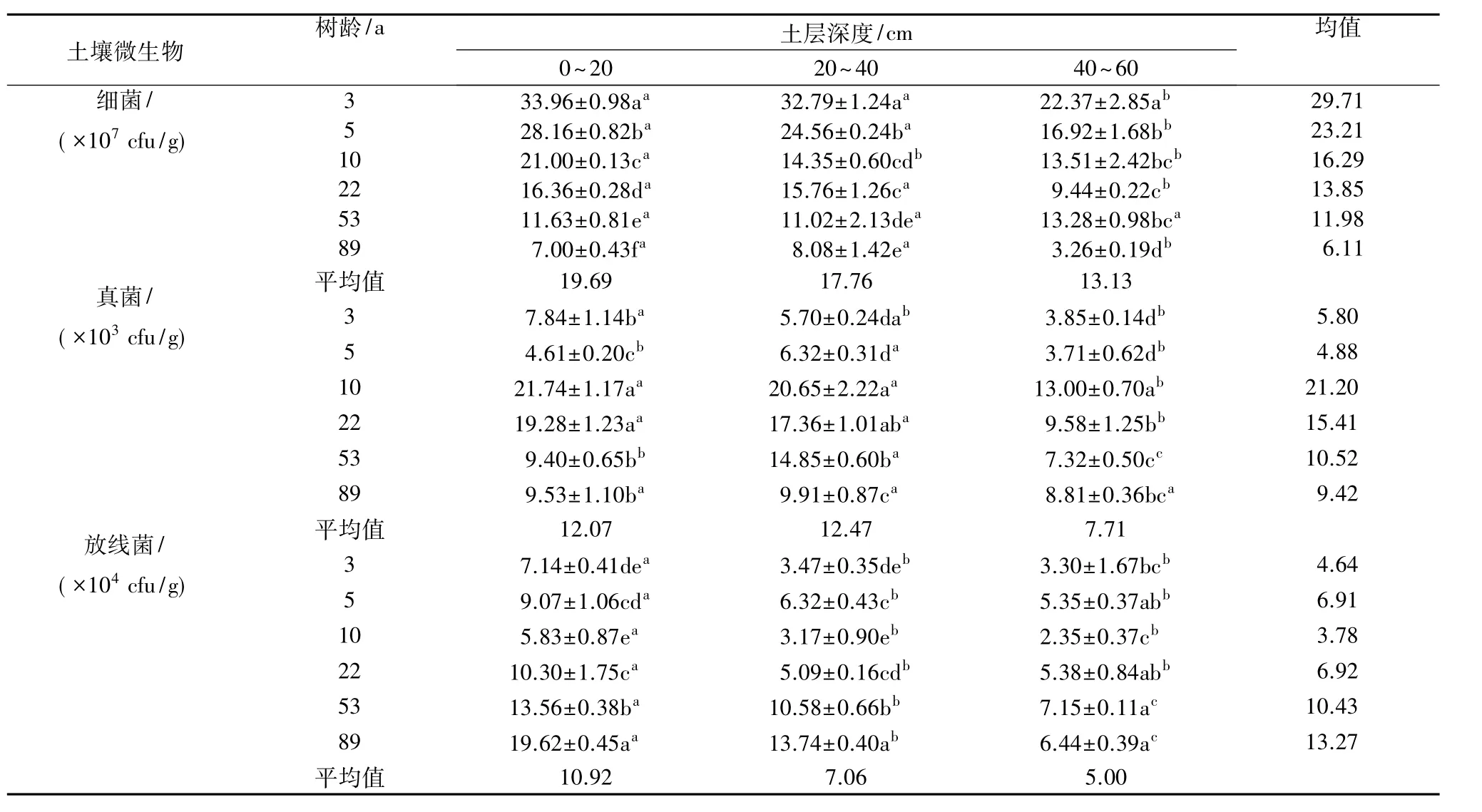
表4 不同树龄梨园不同土层土壤微生物数量的变化
土壤真菌数量在相同土层深度均表现为10 a和22 a梨园最高,显著高于其他树龄梨园,3 a和5 a梨园最低。不同土层相比较,3、10、22 a梨园0~20 cm土层分布最多,5、53、89 a梨园20~40 cm 土层分布最多,各树龄梨园0~20、20~40、40~60 cm 土层真菌数量平均值分别为 12.07×103、12.47×103、7.71×103cfu/g。
土壤放线菌数量在相同土层深度上均表现为10 a梨园最低,53 a和89 a梨园最高。0~20 cm土层89 a梨园土壤放线菌数量显著高于其他各树龄梨园,其次为53 a梨园,且与其他各树龄梨园差异显著;20~40 cm土层89 a梨园显著高于其他各树龄梨园,其次为53 a梨园,且与其他各树龄梨园差异显著,10 a和3 a梨园差异不显著;40~60 cm土层53 a和89 a梨园高于5 a和22 a梨园,但差异不显著,10 a梨园显著低于其他各树龄梨园(3 a除外)。总体上,不同土层放线菌数量随土层深度增加呈现减少的趋势,这与细菌和真菌分布类似。
2.3.2 不同树龄梨园土壤微生物的组成变化 由图1可知,梨园各土层土壤微生物总量总体上随树龄的增加而逐渐降低,而相同树龄不同土层间土壤微生物总量也表现为随土层深度的增加而减少,这与细菌变化趋势一致(表4)。梨园土壤三大微生物细菌、真菌、放线菌在数量组成上,细菌数量占绝对优势,分别为真菌和放线菌的104倍和103倍,因此,微生物总量的变化主要取决于细菌的变化。由表5可知,不同梨园土壤微生物组成中,细菌数量最多,占总微生物数量99%以上,其次为放线菌,真菌最少。随树龄的增加,不同微生物数量组成呈现出不同的变化趋势,细菌数量组成表现为随树龄的增加而逐渐减小,而真菌和放线菌则随树龄的增加而增加。随深度的增加,细菌、真菌、放线菌数量组成总体上变化不大。
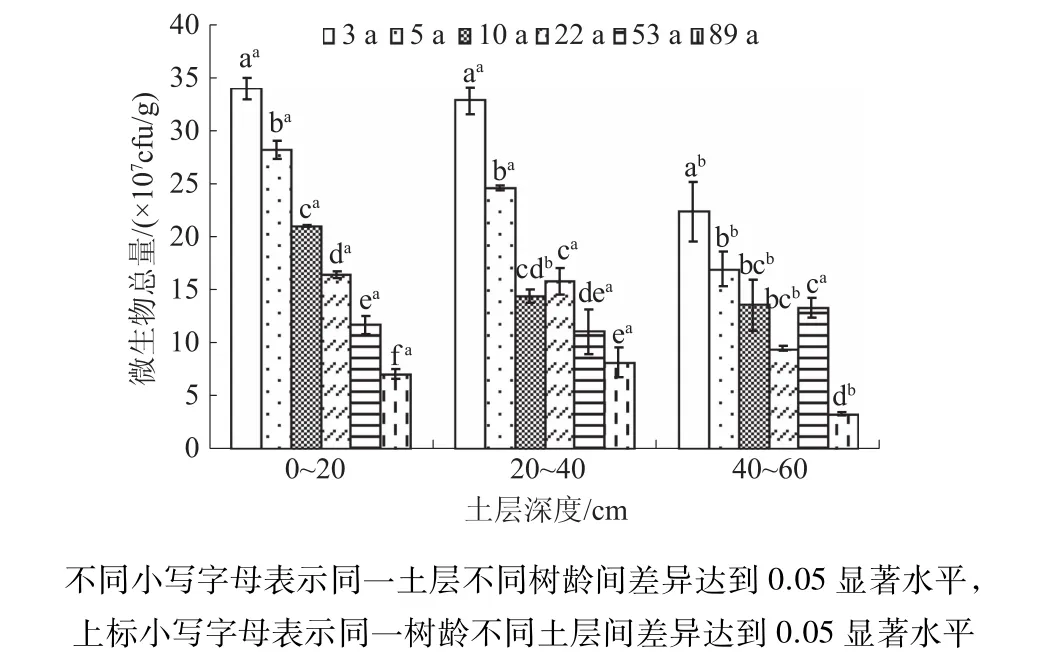
图1 不同树龄梨园土壤微生物总量的变化
2.4 梨园树龄、土壤酶活性与土壤微生物相关性分析
由表6可知,树龄与土壤有机质含量呈显著负相关,与蔗糖酶、过氧化氢酶、脲酶活性呈负相关,其中与土壤脲酶活性相关性达极显著水平,与土壤细菌和真菌数量呈负相关,其中与细菌相关性达显著水平,而与放线菌数量呈极显著正相关。土壤有机质含量与蔗糖酶、过氧化氢酶、脲酶活性呈正相关,其中与脲酶和过氧化氢酶活性相关性达到显著水平,与细菌数量呈极显著正相关,与放线菌数量呈显著负相关。此外,土壤脲酶活性与细菌数量呈显著 正相关,与放线菌数量呈显著负相关。
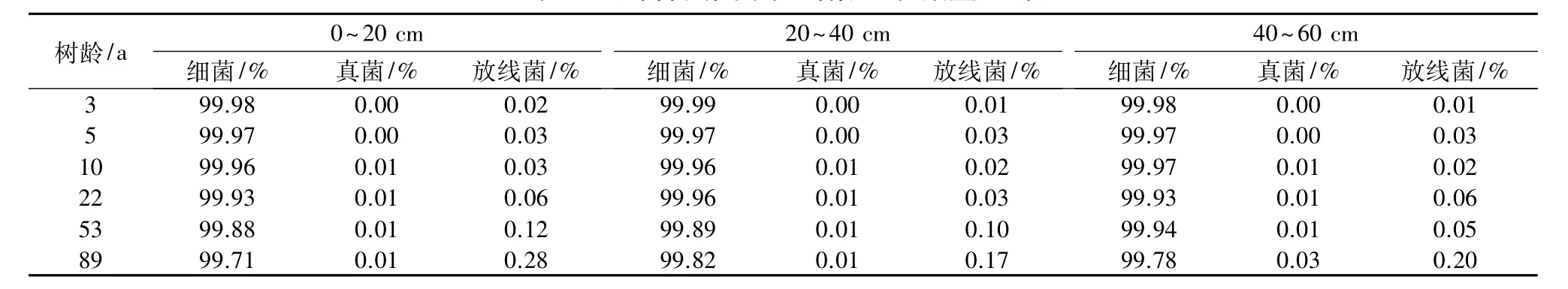
表5 不同树龄梨园土壤微生物数量组成
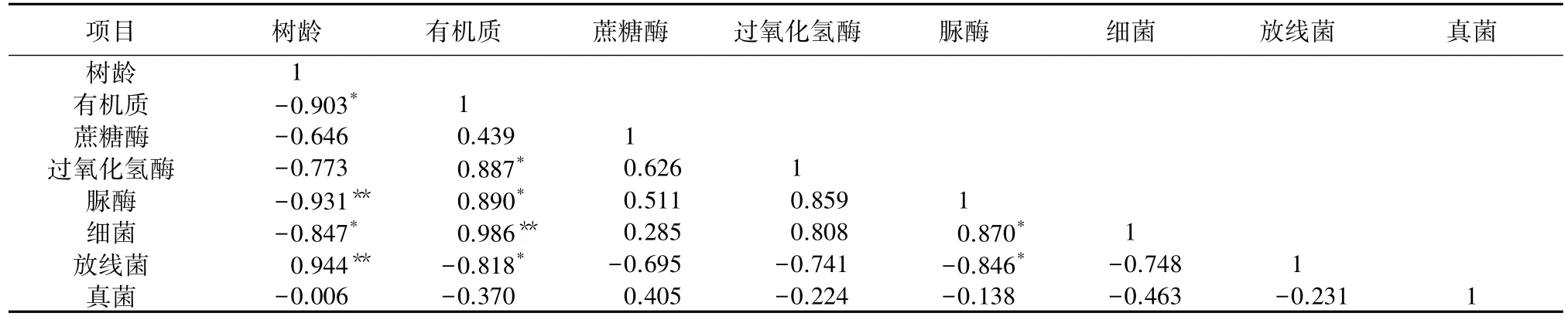
表6 梨园各指标相关性分析
3 结论与讨论
土壤酶是土壤中的生物催化剂,能够加速土壤生化反映速度,在土壤肥力评价中,酶活性是一项重要指标[23]。一般认为,树龄对果园土壤酶活性有一定的影响,本研究中,土壤蔗糖酶活性大致随树龄的增加呈先升高后降低的趋势。杜俊龙等[24]通过对枣园土壤酶活性的研究表明,3、5、10、15 a树龄枣园随种植年限的增加而增加,这与本研究结果一致。但随树龄的继续增加,土壤蔗糖酶活性反而降低。同一年限不同土壤土层间蔗糖酶活性表现为随土壤深度增加不断减小,这与杜俊龙等[24]、杜静静[25]研究结果一致。蔗糖酶是分解土壤腐殖质进行土壤能量转化的重要酶类[26],随土层深度的增加,土壤腐殖质减少,进而蔗糖酶活性降低。土壤脲酶活性随梨园树龄的增加而逐渐降低,同一树龄不同土壤深度表现为随土壤深度增加而减小,这与前人研究结果[27]相似。土壤脲酶活性与土壤肥力有一定的关系,土壤肥力越高,脲酶活性越强[28],土壤有机质含量在一定程度上反映了土壤肥力状况,而土壤脲酶的变化与土壤有机质变化一致。不同树龄、不同土层深度过氧化氢酶活性差异不显著。现阶段关于过氧化氢酶活性与树龄和土层深度的关系研究并未得出一致的结论。王静等[29]研究表明,过氧化氢酶活性与种植年限的相关性不显著,这与本研究结果相似。本研究中,0~60 cm土层过氧化氢酶活性受土层深度的影响变化不大。杨恒山等[30]研究认为,过氧化氢酶活性随土壤深度增加而增加。杜俊龙等[24]认为,过氧化氢酶活性与土层深度有关,但树龄不同,过氧化氢酶活性表现也不相同,具体机制有待进一步研究。
土壤细菌、真菌、放线菌作为土壤微生物的主要群体,影响着土壤的形成和发育,有利于土壤有机物质的分解、矿化和腐殖化,对养分的转化和循环有重要的作用。因此,微生物数量可以在一定程度上反映土壤的肥力状况[31]。本研究中,各梨园土壤微生物类群组成以细菌为主(细菌数量占总微生物的99%以上),且细菌数量随树龄的增加逐渐减少,同时随土层深度增加有减少的趋势;真菌数量表现为壮龄梨园(10 a和22 a)>老龄梨园(53 a和89 a)>幼龄梨园(3 a和5 a),不同土层深度真菌分布与细菌类似,表现出随土层深度增加而减少的趋势;放线菌数量分布则表现为老龄梨园(53 a和89 a)最高,而10 a梨园最低,不同土层深度放线菌分布与细菌和真菌类似,均表现为随土层深度增加而逐渐减少;但微生物总量变化与细菌保持一致,即随树龄的增加而逐渐减少,且随土层深度增加也表现为逐渐减少的趋势,这与前人研究结果相似[16]。微生物数量随树龄的增加而逐渐减少,原因可能是幼龄梨园表层土壤的耕作活动较为频繁,土壤孔隙度大,增加了土壤中氧含量,有利于土壤微生物的生长繁殖,而老龄梨园受人为干扰较大,土壤容重增加,孔隙度降低,土壤氧含量下降,抑制了好氧微生物的繁殖。此外,随土层深度增加,土壤中营养物质(主要为土壤速效养分和有机质)相应减少,土壤微生物数量与土壤的肥沃程度存在一定的相关性[16,19],这也是造成微生物数量减少的原因。
土壤微生物数量和酶活性能在一定程度上反映土壤的肥力状况,本研究初步分析了树龄、土壤酶活性与微生物数量之间的关系,结果显示,梨园树龄与土壤细菌数量呈显著负相关,即细菌数量随树龄增加而减少。但有研究指出盛果期细菌数量最大[16,31],原因可能是微生物分布除了受果园年龄影响外,还受土壤养分状况、孔隙度、温度和湿度等外部因素的影响,因此造成了研究结果不一致。另外,本研究表明,树龄与土壤脲酶活性呈极显著负相关,土壤脲酶活性在一定程度上反映了土壤氮素的情况[32],因此梨园随树龄的增加,氮素供应能力有所下降。
综上所述,土壤微生物生态特征与树龄存在着一定的关系,这种关系同时受到人类活动的影响,随梨园树龄增长,土壤微生物生态特征及土壤营养状况发生变化,可能会影响果树的产量和品质,因此,需要合理进行土壤调控管理以保证梨园土壤微生物生态平衡。
[1] 欧阳志云,王效科,苗鸿.中国陆地生态系统服务功能及其生态经济价值的初步研究[J].生态学报,1999,19(5):607-613.
[2] 杨海君,肖启明,刘安元.土壤微生物多样性及其作用研究进展[J].南华大学学报(自然科学版),2005,19(4):21-27.
[3] Caldwell B A.Enzyme activities as a component of soil biodiversity:A review [J].Pedobiologia,2005,49(6):637-644.
[4] Acosta-Martínez V,Cruz L,Sotomayor-Ramírez D,et al.Enzyme activities as affected by soil properties and land use in a tropical watershed[J].Applied Soil Ecology,2007,35(1):35-45.
[5] 马理辉,吴普特,汪有科.黄土丘陵半干旱区密植枣林随树龄变化的根系空间分布特征[J].植物生态学报,2012,36(4):292-301.
[6] 甘卓亭,刘文兆.渭北旱塬不同龄苹果细根空间分布特征[J].生态学报,2008,28(7):3401-3407.
[7] 刘维佳,李疆,时坤,等.不同树龄库尔勒香梨园土壤有机碳储量及其对树体生长的影响[J].新疆农业科学,2012,49(2):209-215.
[8] 马艳春,姚玉新,杜远鹏,等.葡萄设施栽培不同种植年限土壤理化性质的变化[J].果树学报,2015,32(2):225-231.
[9] 周怀平,关春林,杨治平,等.不同树龄仁用杏人工林地土壤水肥状况研究[J].水土保持学报,2007,20(6):137-140.
[10] 胥继东,王益权,刘军,等.渭北旱塬不同树龄果园土壤营养状况演化趋势[J].安徽农业科学,2008,36(31):13722-13724.
[11] 孙霞,陈新燕,柴仲平,等.不同土壤管理措施下南疆果园土壤微生物及酶活性特征[J].草业科学,2012,29(7):1023-1027.
[12] 徐凌飞,韩清芳,吴中营,等.清耕和生草梨园土壤酶活性的空间变化[J].中国农业科学,2010,43(23):4977-4982.
[13] 路超,李絮花,董静,等.渗灌条件下果园覆盖的保水效果及对根际土壤养分和微生物特性的影响[J].水土保持学报,2013,27(6):134-139.
[14] 李丙智,王桂芳,秦晓飞,等.沼液配施钾肥对果园土壤理化特性和微生物及果实品质影响[J].中国农业科学,2010,43(22):4671-4677.
[15] 温晓霞,殷瑞敬,高茂盛,等.不同覆盖模式下旱作苹果园土壤酶活性和微生物数量时空动态研究[J].西北农业学报,2011,20(11):82-88.
[16] 田稼,孙超,杨明琰,等.黄土高原不同树龄苹果园土壤微生物,养分及pH的相关性[J].西北农业学报,2012,21(7):138-141.
[17] 王瑛,艾训儒.连作对土壤养分、微生物特性及葡萄产质量的影响[J].河南农业科学,2016,45(7):49-55.
[18] 官雪芳,林碧芬,徐庆贤,等.种植年限对土壤性状、微生物群落及脐橙果实品质的影响[J].浙江农业学报,2012,24(1):105-113.
[19] 付学琴,黄文新.不同树龄南丰蜜橘根际土壤微生物群落多样性分析[J].园艺学报,2014,41(4):631-640.
[20] 关松荫.土壤酶及研究方法[M].北京:农业出版社,1986:274-332.
[21] 林先贵,王一明.土壤微生物研究原理与方法[M].北京:高等教育出版社,2010:30-47.
[22] 鲍士旦.土壤农化分析[M].3版.北京:中国农业出版社,2000:25-114.
[23] 张焱华,吴敏,何鹏,等.土壤酶活性与土壤肥力关系的研究进展[J].安徽农业科学,2007,35(34):11139-11142.
[24] 杜俊龙,孙霞,黄长福,等.种植年限对干旱区枣园土壤酶活性的影响——以新疆麦盖提县为例[J].天津农业科学,2015,21(2):89-92.
[25] 杜静静.不同种植年限果园土壤理化性质与酶活性研究[D].临汾:山西师范大学,2013.
[26] 陈继峰,孙会,夏阳,等.不同连作年限烟田土壤酶活性及养分含量变化[J].河南农业科学,2016,45(10):60-64.
[27] 马云华,魏珉,王秀峰.日光温室连作黄瓜根区微生物区系及酶活性的变化[J].应用生态学报,2004,15(6):1005-1008.
[28] 贾若凌,李丽,刘香玲,等.荔枝果园土壤脲酶活性与土壤肥力的关系研究[J].河南农业科学,2011,40(6):79-81.
[29] 王静,呼丽萍,李昶,等.种植年限对樱桃园土壤养分和酶活性的影响[J].水土保持通报,2013,33(4):155-158,165.
[30] 杨恒山,张庆国,邰继承,等.种植年限对紫花苜蓿地土壤pH值和磷酸酶活性的影响[J].中国草地学报,2009,31(1):32-35.
[31] 甘卓亭,张蓓蓓,张掌权,等.渭北塬区不同龄苹果园土壤微生物空间分布特征[J].生态学报,2015,35(21):6965-6973.
[32] 刘建国,卞新民,张伟,等.长期连作和秸秆还田对棉田土壤生物活性的影响[J].应用生态学报,2008,19(5):1027-1032.
猜你喜欢
现代园艺(2022年19期)2022-09-21
东南大学学报(自然科学版)(2022年4期)2022-08-12
小读者(2021年19期)2021-11-07
腐植酸(2020年1期)2020-11-29
果树资源学报(2020年3期)2020-05-13
西北农业学报(2020年1期)2020-02-07
新疆农业科学(2019年11期)2019-12-31
作文评点报·中考版(2019年4期)2019-02-18
课外语文·中(2018年10期)2018-11-16
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
