
蜜蜂RNA病毒在传粉昆虫中的分布及其病害诊断防治方法
2018-03-14 11:24史红霞刘永梅党晓群
河南农业科学 2018年1期
史红霞,刘永梅,党晓群
(重庆师范大学生命科学学院,重庆401331)
自然界中90%的被子植物有性繁殖直接或间接依赖于传粉昆虫。据统计,在全球范围内传粉昆虫带来的经济增益每年至少有2 250亿美元,非蜜蜂属膜翅目昆虫传粉的水果和蔬菜价值约30亿美元[1]。然而近10 a间,由于环境污染、农药的大量使用以及蜂群崩溃失调症(Colony collapse disorder,CCD)等,全球范围内蜜蜂种群数量正在不断减少。北美地区在1947—2005年蜂群数量减少了59%,欧美地区1985—2005年则下降了25%[2-3]。4种之前在美国本土数量十分充足的大黄蜂数量下降了约96%[3]。2012—2013 年美国蜜蜂损失比例高达45%[4],由此带来的对农业和生态系统的负面影响广受农业界及科学界的关注。
蜜蜂是集居的社会性昆虫,蜂群成员间密切接触使得它们更易受到各种传染性疾病感染。到目前为止,在蜜蜂的不同级型、不同生长阶段以及在其蜂巢、食物、寄生虫等样本中均能检测到蜜蜂病毒[5]。现已分离鉴定的蜜蜂病毒超过22种。由于蜜蜂和其他传粉昆虫如大黄蜂、独居蜂、胡蜂、苍蝇、蚂蚁、蝴蝶等所共有的采蜜行为,加速了它们之间病原体的传播[6]。蜜蜂病毒的广泛传播可能是传粉昆虫数量下降的原因之一。重点概述了蜜蜂RNA病毒在传粉昆虫中的分布及致病性,并对蜜蜂病毒的跨物种传播作一探讨,为蜜蜂病害综合防控体系的建设提供理论参考。
1 蜜蜂RNA病毒在传粉昆虫中的分布
1.1 蜜蜂卷翅病毒
蜜蜂卷翅病毒(Deformed wing virus,DWV)不仅是全球蜜蜂种群中最流行的病毒,也是寄主范围最广的蜜蜂病毒(表1)。DWV除通过瓦螨(Varroa destructor)传播至东方蜜蜂(Apis cerana)外,也可以在非人工饲养的大蜜蜂(Apis dorsata)、小蜜蜂(Apis florea)、普通黄胡蜂(Vespula vulgaris)、小木匠蜂(Ceratina dupla)、红壁蜂(Osmia bicornis)种群中广泛传播[7-9](表 1)。Gamboa 等[10]研究发现,在哥伦比亚的热带熊蜂(Bombus atratus)中也存在DWV。北美数量最多的几种熊蜂如东方熊蜂(Bombus impatiens)、亨特大黄蜂(Bombus huntii)和墨西哥熊蜂(Bombus vagans)中也检测到了DWV,DWV对欧洲熊蜂(Bombus terrestris)的致死率高达 50%[11-13]。
DWV的寄主域不仅局限于昆虫纲,越来越多的研究表明,DWV可以在瓦螨中复制增殖[14]。DWV可以诱导感染瓦螨的蜂蛹产生免疫抑制,促进瓦螨繁殖。Di等[15]在研究瓦螨与病毒共生对蜜蜂免疫系统的影响时提出,DWV和狄斯瓦螨是互惠共生的关系,DWV对蜜蜂个体和群体的致病性取决于寄生螨类的多少。狄斯瓦螨不仅可以影响DWV在蜂群内的多样性,而且还影响DWV在蜂群中的感染力和致病力。
1.2 蜜蜂黑蜂王台病毒
蜜蜂黑蜂王台病毒(Black queen cell virus,BQCV)于1977年由Bailey等[16]首次从西蜂蜂王前蛹期的黑色细胞中分离出,是引起蜂王幼虫死亡的主要原因。BQCV一度被认为是西蜂中最为流行的病毒之一。近几年来,在西双版纳野生小蜜蜂和大蜜蜂以及台湾、日本的Apis cerana indica、Apis cerana japonica 蜂群中也相继检测到了 BQCV[17-18],对BQCV的核衣壳蛋白进行系统进化树分析,推测BQCV有可能是从西蜂传播到大蜜蜂和小蜜蜂[18]。BQCV不仅可以在蜜蜂属内进行种间传播,还可以感染非膜翅目昆虫、蜂箱小甲虫(Aethina tumida)、欧洲球螋(Forficcula auricularia)、大蜡螟(Galleria mellonella)以及蜘蛛等[8,19-20]。Singh 等[6]采集了宾夕法尼亚州、纽约、伊利诺斯州等地区的蜜蜂以及一些非蜜蜂属膜翅目昆虫,并收集了它们携带的花粉,经研究发现,同一地区未患病蜜蜂和健康非蜜蜂属膜翅目昆虫及其携带的花粉中均可检测到BQCV,推测BQCV可能通过传粉昆虫共同的访花行为经由花粉传播至其他昆虫。
1.3 AKI复合体
蜜蜂急性麻痹病毒(Acute bee paralysis virus,ABPV)、蜜蜂克什米尔病毒(Kashmir bee virus,KBV)以及以色列急性麻痹病毒(Israeli acute paralysis virus,IAPV)称之为 AKI复合体[21],三者可复合感染蜜蜂的弱势群和健康群。该病主要在西蜂中暴发,在中蜂中比较少见。Bakonyi等[22]在西蜂瓦螨中检测到 ABPV,但到目前为止,没有关于ABPV在瓦螨中复制的报道。ABPV能感染热带熊蜂[10],而且对巴西蜜蜂的感染率为 100%[23]。
IAPV和KBV、ABPV基因组相似性分别为75%和 65%[24]。Berényi等[25]猜测 IAPV 可能是 KBV的亚型。KBV和ABPV能感染相同的蜂群,甚至同一只蜜蜂。Cox-Foster等[26]采用宏基因组的方法,对正常巢脾和出现CCD症状的巢脾中微生物群落进行RNA测序分析,发现KBV、IAPV与CCD关联度最大。Singh等[6]对11种非蜜蜂属膜翅目传粉昆虫的病毒进行检测发现,在欧洲熊蜂、普通黄胡蜂、墨西哥熊蜂、汗蜂(Augochlora pura)等中均能检测到 IAPV(表 1)。
1.4 蜜蜂囊状幼虫病毒
蜜蜂囊状幼虫病是1913年首种被发现的蜜蜂病毒病,其病原体为蜜蜂囊状幼虫病毒(Sacbrood virus,SBV)[27]。SBV 可感染成蜂和幼虫,患病幼虫体色会由乳白色逐渐变黄,不能正常化蛹,最后变成囊状,但对成蜂威胁不严重。目前,对于SBV已经分离出几种亚洲毒株如韩国株(Korea sacbrood virus,KSBV)[28]、中国株 (Chinese sacbrood virus,CSBV)[29]、泰国株(Thai sacbrood virus,TSBV)[30]。SBV与CSBV同源性很高,不交叉感染。但Gong等[31]发现,感染中华蜜蜂的SBV也可以感染西蜂,具有一定的致病性。瓦螨中虽能检测到SBV的存在,但其不能复制,这说明瓦螨可能作为SBV的载体去感染其他生物[32]。关于SBV在非蜜蜂属昆虫中寄生的报道很少(表1),仅在一些熊蜂和Andrena vaga中检测到[9],暗示SBV具有较窄的寄主域。
1.5 蜜蜂慢性麻痹病毒和缓慢性麻痹病毒
蜜蜂慢性麻痹病毒(Chronic bee paralysis virus,CBPV)是一种还未分类的病毒。该病毒为正义单链RNA病毒,基因组由5段组成。目前已完成测序的是由3 647个碱基组成的RNA1和由2 305个碱基组成的RNA2,其余为其卫星病毒所有[33]。CBPV多在西蜂蜂群中流行,但在东方蜜蜂、欧洲熊蜂、东方熊蜂、蚂蚁、瓦螨中均有分布,推测瓦螨可作为CBPV 的载体,加快CBPV侵染[34-35]。
蜜蜂缓慢性麻痹病毒(Slow bee paralysis virus,SBPV)属于传染性软腐病毒科。Mcmahon等[17]对从英国和比利时收集到的几种野生熊蜂属昆虫病毒进行检测发现,SBPV可感染熊蜂,但相对于蜜蜂中检测到的SBPV,其在熊蜂中的感染水平更高,推测SBPV可能由其他传粉昆虫传播至蜜蜂[36]。
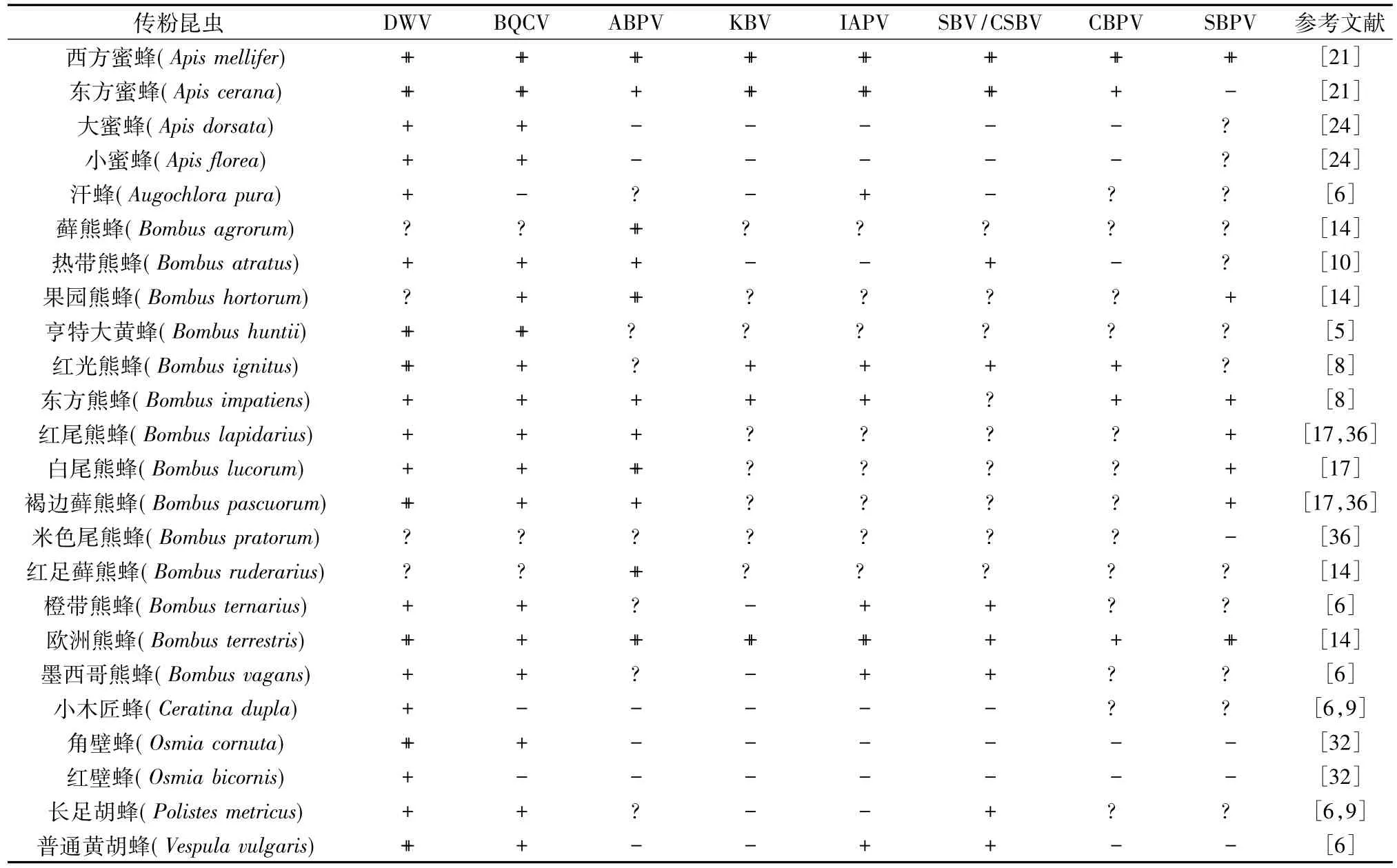
表1 蜜蜂病毒在传粉昆虫中的分布
2 蜜蜂病毒病的诊断与防治
2.1 蜜蜂病毒病的诊断
对于蜜蜂病毒病形态学的检测,一般都是基于蜜蜂的患病症状、流行病学初步作出判断,然后进行电镜观察,根据其病原特征,确定病毒类型(表2)。然而,对于致病症状类似的蜜蜂病毒,如以色列急性麻痹病毒、蜜蜂慢性麻痹病毒、缓慢性麻痹病毒均会诱发蜜蜂出现翅膀震颤、失去飞行能力、爬蜂等现象,并且病毒粒子多为直径20~30 nm的正二十面体,外形也比较相似,仅仅依靠症状和电镜观察很难将病原体进行区分,此时则可借助分子生物学诊断技术来鉴定。
目前,已有的蜜蜂病毒病检测方法主要有琼脂凝胶免疫扩散法、酶联免疫法、电子显微镜法、蛋白质印记法、寡核苷酸微列阵芯片技术、RT-PCR法、RT-qPCR法、多重连接依赖式探针扩增技术。酶联免疫法可高通量检测多种病毒。免疫胶体金试纸条检测法使用方便快速,便于基层和现场使用,所有反应能在15 min内完成,成本低,应用范围广。而分子检测的极高灵敏度也使其成为目前进行蜜蜂病毒学研究的主要工具。其中,寡核苷酸微列阵芯片和多重连接依赖式探针扩增技术可以同时检测9~10种病毒。RT-PCR、RT-qPCR可以定量和定性地检测病毒,目前应用范围最广[37-39]。

表2 几种常见蜜蜂病毒病的病原特征和症状
2.2 蜜蜂病毒病的防治
根据蜜蜂病毒病的病因、病源和传播途径,研究者进行了广泛的防治技术研究。概而言之,主要有物理防治和药物防治2种。蜂农常通过蜂场、蜂具及饲料的卫生与消毒来预防。另外,还可通过选育抗病蜂种、选择耐病性好的蜂种进行预防。药物防治包括抗生素的使用和中草药饲喂,这些仍是蜂农们赖以使用的有效方法,但药物残留对蜜蜂健康以及蜂产品质量都有不同程度的影响。
RNAi(RNA interference)技术是目前实现疾病治疗的有效方法之一。该技术具有特异性和高效性,可以特异地沉默或下调某个基因的表达水平,从分子水平上削弱或消除病毒的增殖。2016年,韩日畴课题组以中蜂SBV结构蛋白VP1基因为靶标基因,进行dsRNA干扰试验,发现蜜蜂幼虫的死亡率明显下降[40],为中蜂囊状幼虫病毒病乃至所有蜜蜂病毒病的防治提供了新的思路。
3 展望
随着全球化进程加剧,传粉昆虫的商业化使得本土传粉昆虫被引入到其他国家,如原产于欧洲、非洲和中东的西方蜜蜂已被引入世界各地。20世纪80年代后期,大黄蜂、熊蜂等也相继输入到不同国家,对当地传粉昆虫的健康造成严重威胁,也直接影响了当地生态系统的平衡。蜜蜂RNA病毒在传粉昆虫中的广泛分布可能是导致世界范围内传粉昆虫数量急剧下降的原因之一。RNA病毒的跨物种传播仍有很多问题亟待研究。第一,对于大部分传粉昆虫,是否存在蜜蜂RNA病毒还未展开研究,对传粉昆虫中病毒的全面调查分析将为病毒的跨物种传播研究提供更多线索,以便评估传粉昆虫的潜在健康威胁。第二,蜜蜂是否通过采集带病毒的花粉传播病毒,花粉或蜂蜜中的病毒是否具有感染性,均不清楚。第三,自然界中病毒与花粉植物之间存在怎样的互作关系,花粉在病毒的跨物种传播中有何作用,需要阐明。第四,蜜蜂病毒的寄主域如何,也需要明确。研究表明,DWV、BQCV和AKI具有较宽的寄主域,而SBV、CBPV和SBPV的寄主范围则相对较窄,提示DWV、BQCV和AKI的传粉昆虫应避免商业性输出。因此,应加强传粉昆虫病毒学的深入研究,评估这些病毒对传粉昆虫健康的影响,揭示不同传粉昆虫在病毒传播中扮演的角色,进而抑制具有广泛寄主域的病毒的流行与传播,缓解现今传粉昆虫数量急剧下降的问题。传粉昆虫病毒学的研究对在自然生态系统中建立病原体的综合防控监测体系也具有重要意义。
[1] Robinson W S,Nowogrodzki R,Morse R A.The value of honey bees as pollinators of U.S.crops[J].American Bee Journal,1989,129(13):411-423.
[2] Engelsdorp D V,Hayes J J,Underwood R M,et al.A survey of honey bee colony losses in the United States,fall 2008 to spring 2009[J].Journal of Apicultural Research,2010,49(1):7-14.
[3] National Research Council.Status of pollinators in North America[M].Washington,D C:The National Academies Press,2007:34-103.
[4] USDA.Beekeepers lost 44%of honey bee colonies last year[EB/OL].(2016-05-10) [2017-07-26].https://www.ecowatch.com/usda-beekeepers-lost-44-of-honeybee-colonies-last-year-1891129451.html.
[5] Chen Y P,Siede R.Honey bee viruses[J].Advances in Virus Research,2007,70(1):33-80.
[6] Singh R,Levitt A L,Rajotte E G,et al.RNA viruses in hymenopteran pollinators:Evidence of inter-taxa virus transmission via pollen and potential impact on non-Apis Hymenopteran species[J].PLoS ONE,2010,5(12):e14357.
[7] Reynaldi F J,Sguazza G H,Albicoro F J,et al.First molecular detection of co-infection of honey bee viruses in asymptomatic Bombus atratus in south America[J].Brazilian Journal of biology,2013,73(4):797-800.
[8] Levitt A L,Singh R,Coxfoster D L,et al.Cross-species transmission of honey bee viruses in associated arthropods[J].Virus Research,2013,176(1/2):232-240.
[9] Ravoet J,De S L,Meeus I,et al.Widespread occurrence of honey bee pathogens in solitary bees[J].Journal of Invertebrate Pathology,2014,122:55-58.
[10] Gamboa V,Ravoet J,Brunain M,et al.Bee pathogens found in Bombus atratus from Colombia:A case study[J].Journal of Invertebrate Pathology,2015,129:36-39.
[11] Genersch E,Yue C,Fries I,et al.Detection of deformed wing virus,a honey bee viral pathogen,in bumble bees(Bombus terrestris and Bombus pascuorum)with wing deformities[J].Journal of Invertebrate Pathology,2006,91(1):61-63.
[12] Graystock P,Meeus I,Smagghe G,et al.The effects of single and mixed infections of Apicystis bombi and deformed wing virus in Bombus terrestris[J].Parasitology,2016,143(3):358-365.
[13] Li J,Peng W,Wu J,et al.Cross-species infection of deformed wing virus poses a new threat to pollinator conservation[J].Journal of Economic Entomology,2011,104(3):732-739.
[14] Gisder S,Genersch E.Viruses of commercialized insect pollinators[J].Journal of Invertebrate Pathology,2016,147:51-59.
[15] Di P G,Annoscia D,Margiotta M,et al.A mutualistic symbiosis between a parasitic mite and a pathogenic virus undermines honey bee immunity and health[J].Proceedings of the National Academy of Sciences of the United States of America,2016,113(12):3203-3208.
[16] Bailey L,Woods R D.Two more small RNA viruses from honey bees and further observations on sacbrood and acute bee paralysis viruses[J].Journal of General Virology,1977,37(1):175-182.
[17] Mcmahon D P,Fürst M A,Caspar J,et al.A sting in the spit: Widespread cross-infection ofmultiple RNA viruses across wild and managed bees[J].Journal of Animal Ecology,2015,84(3):615-624.
[18] Zhang X,He S Y,Evans J D,et al.New evidence that deformed wing virus and black queen cell virus are multihost pathogens[J].Journal of Invertebrate Pathology,2012,109(1):156-159.
[19] Donaldson E F,Haskew A N,Gates J E.Metagenomic analysis of the viromes of three North American bat species:Viral diversity among different bat species that share a common habitat[J].Journal of Virology,2010,84(24):13004-13018.
[20] Peng W,Li J,Boncristiani H,et al.Host range expansion of honey bee black queen cell virus in the bumble bee,Bombus huntii[J].Apidologie,2011,42(5):650-658.
[21] Forsgren E,Wei S,Ding G L,et al.Preliminary observations on possible pathogen spill-over from Apis mellifera to Apis cerana[J].Apidologie,2014,46(3):1-11.
[22] Bakonyi T,Farkas R,Szendroi A,et al.Detection of acute bee paralysis virus by RT-PCR in honey bee and Varroa destructor field samples: Rapid screening of representative Hungarian apiaries[J].Apidologie,2002,33(1):63-74.
[23] Ueira-Vieira C,Almeida L O,Almeida F C D,et al.Scientific note on the first molecular detection of the acute bee paralysis virus in Brazilian stingless bees[J].Apidologie,2015,46(5):628-630.
[24] 张炫,周丹银,和绍禹.蜜蜂病毒病的检测与诊断[J].中国蜂业,2012,63(Z4):56-63.
[25] Berényi O,Bakonyi T,Derakhshifar I,et al.Occurrence of six honeybee viruses in diseased austrian apiaries[J].Applied & EnvironmentalMicrobiology,2006,72:2414-2420.
[26] Cox-Foster D L,Conlan S,Holmes E C,et al.A metagenomic survey of microbes in honey bee colony collapse disorder[J].Science,2007,318(5848):283-287.
[27] White G F.Sacbrood,a disease of bees[M].New York,USA:Bureau of Entomology,1913:1-5.
[28] Reddy K E,Yoo M S,Kim Y H,et al.Analysis of the RdRp,intergenic and structural polyprotein regions,and the complete genome sequence of Kashmir bee virus from infected honeybees(Apis mellifera)in Korea[J].Virus Genes,2014,49(1):137-144.
[29] Zhang Y,Huang X,Xu Z F,et al.Differential gene transcription in honeybee(Apis cerana)larvae challenged by Chinese sacbrood virus(CSBV)[J].Sociobiology,2013,60(4):413-420.
[30] Rana R,Rana B S,Kaushal N,et al.Identification of sacbrood virus disease in honeybee,Apis mellifera by using ELISA and RT-PCR techniques[J].Indian Journal of Biotechnology,2011,10(3):274-284.
[31] Gong H R,Chen X X,Chen Y P,et al.Evidence of Apis cerana sacbrood virus infection in Apis mellifera[J].Applied & Environmental Microbiology,2016,82(8):2256-2262.
[32] Mondet F,Miranda J R D,Kretzschmar A,et al.On the front line:Quantitative virus dynamics in honeybee(Apis mellifera)colonies along a new expansion front of the parasite Varroa destructor[J].PLoS Pathogens,2014,10(8):e1004323.
[33] 钟义,邵海英,傅丽君,等.慢性蜜蜂麻痹病毒半套式PCR检测方法的建立[J].病毒学报,2016,4(32):490-494.
[34] Celle O,Blanchard P,Olivier V,et al.Detection of chronic bee paralysis virus(CBPV)genome and its replicative RNA form in various hosts and possible ways of spread[J].Virus Research,2008,133(2):280-284.
[35] 汪天澍,施腾飞,刘芳,等.安徽省七种蜜蜂病毒的发生与流行研究[J].应用昆虫学报,2015,52(2):324-332.
[36] Parmentier L,Smagghe G,de Graaf D C,et al.Varroa destructor macula-like virus,lake sinai virus and other new RNA viruses in wild bumblebee hosts(Bombus pascuorum,Bombus lapidarius and Bombus pratorum) [J].Journal of Invertebrate Pathology,2016,134:6-11.
[37] Glover R H,Adams I P,Budge G,et al.Detection of honey bee (Apis mellifera) viruses with an oligonucleotide microarray[J].Journal of Invertebrate Pathology,2011,107(3):216-219.
[38] De Smet L,Ravoet J,De Miranda J R,et al.BeeDoctor,a versatile MLPA-based diagnostic tool for screening bee viruses[J].PLoS ONE,2012,7(10):e47953.
[39] 刘学录,童金凤,马振刚.蜜蜂主要病害及其病原PCR检测研究进展[J].南方农业学报,2016,47(1):147-152.
[40] Zhang J Q,Zhang Y,Han R.The high-throughput production of dsRNA against sacbrood virus for use in the honey bee Apis cerana(Hymenoptera:Apidae) [J].Virus Genes,2016,52(5):1-8.
猜你喜欢
中国科学探险(2022年8期)2022-12-31
今日农业(2021年6期)2021-06-09
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
小哥白尼(军事科学)(2020年4期)2020-07-25
河北省科学院学报(2020年1期)2020-05-25
大科技·百科新说(2018年3期)2018-04-11
儿童故事画报·发现号趣味百科(2017年10期)2018-03-13
儿童故事画报·发现号趣味百科(2017年9期)2018-03-13
儿童故事画报·发现号趣味百科(2016年4期)2016-06-24
现代计算机(2016年17期)2016-02-28
