
Proteomic analysis of trans-hemispheric motor cortex reorganization following contralateral C7 nerve transfer
2018-03-14 07:38:40YinYuanXiuyueXuJieLaoXinZhao
中国神经再生研究(英文版) 2018年2期
Yin Yuan, Xiu-yue Xu, Jie Lao, Xin Zhao,
1 Department of Hand Surgery, Huashan Hospital, Fudan University, Shanghai, China
2 Key Laboratory of Hand Reconstruction, Ministry of Health, Shanghai, China
3 Shanghai Key Laboratory of Peripheral Nerve & Microsurgery, Shanghai, China
Introduction
Nerve transfer is the most common treatment for total brachial plexus avulsion injury (TBPI). During the past several decades, many studies have been conducted to find new donor nerves, such as the intercostal nerve (Seddon, 1963;Narakas, 1978; Krakauer and Wood, 1994), accessory nerve(Allieu et al., 1984), phrenic nerve (Brunelli and Monini,1984; Nagano et al., 1995; Gu and Ma, 1996; Chuang et al.,2005; Liu et al., 2015b), and the contralateral C7(CC7) nerve(Gu et al., 1998; Chuang and Hernon, 2012; Tu et al., 2014;Mathews et al., 2017).
After TBPI and CC7nerve transfer, the movement of the injured limb will be activated by certain movements of the healthy limb at the early stage of recovery. This phenomenon is because the motor cortex for the CC7is located on the injured side, so the movement of the healthy limb, which was originally innervated by the transferred C7nerve, can activate the injured limb. Some patients can regain independent motion, but in others, synchronous motion of the healthy limb is maintained (Songcharoen et al., 2001). Many studies have shown that there is a trans-hemispheric reorganization process following CC7nerve transfer (Lou et al., 2006; Navarro et al., 2007; Jiang et al., 2010). Lundborg et al. (2006)found that after peripheral nerve injury, adjacent motor cortices encroach on the original motor cortical representation of the injured nerve and this process is referred to as phase-one reorganization, as described in previous studies(Donoghue et al., 1990; Kaas et al., 1990; Sanes et al., 1990;Merzenich and Jenkins, 1993; Qi et al., 2000). Pan et al. (2012)found that this phase-one reorganization was a harmful factor for recapture of brachial plexus motor cortices. Thus, if we can find a way to restrain or shorten this process, we may accelerate trans-hemispheric cortical functional remodeling.However, the mechanism of this process remains unclear.Previous studies have only focused on functional magnetic resonance imaging (Beaulieu et al., 2006; Sokki et al., 2012;Yu et al., 2017) and the proteins brain-derived neurotrophic factor (BDNF) and growth-associated protein 43 (GAP43)(Nicolelis et al., 1993; Lang et al., 2008; Zhao and Xu, 2008;Wei et al., 2011). There have been no studies using proteomics to analyze protein expression.
The purpose of our experiment was to screen for differential protein expression in the motor cortex after CC7nerve transfer for TBPI by using isobaric tags for relative and absolute quantitation (iTRAQ), and western blot assays. We identified differentially expressed proteins that might play a role in the motor cortex reorganization.
Materials and Methods
Animals
A total of 90, 2-week-old, female Sprague-Dawley rats weighing approximately 200 g were used for surgery. The rats were supplied and housed in the Laboratory Animal Room of the Medicine College of Fudan University of China(license number: SCXK 2014-0004) with free access to food and water. All animal protocols were performed in strict accordance with the Guide for the Care and Use of Laboratory Animals described by the U.S. National Institutes of Health and conformed to guidelines of the Society for Neuroscience and the Neurotrauma Society.
The rats were randomly and equally divided into three groups. The right forepaw was selected as the affected limb.CC7group: The C5–T1nerve roots for the affected limb were avulsed and the entire root of the CC7nerve was transected and transferred subcutaneously to the median nerve, bridged by the ulnar nerve of the affected limb. TBPI group: The C5–T1nerve roots were avulsed without any further operation on the affected limb. Control group: C5–T1nerve roots on both limbs were exposed as for the CC7group, but they were not avulsed and no further surgery was performed.
Model establishment of TBPI and CC7 nerve transfer
Rats were anesthetized with 40 mg/kg sodium pentobarbital and all had supraclavicular exposure of brachial plexus.The C5–T1nerve roots on the right side were avulsed from the spinal column using forceps and 1 cm portions of the avulsed roots were excised to prevent intraplexus nerve regeneration. In the CC7group, the CC7nerve (left side) was transected at the level of its division; the ulnar nerve of injury side (right side) was transected at the levels of the wrist and the axilla as the bridge between the CC7nerve and the median nerve of the injury side. The ulnar nerve was isolated together with the superior ulnar collateral artery, which was used as its nutrient artery. The distal stump was reverted and transferred to the contralateral neck through a subcutaneous tunnel under the chest, and coapted to the CC7root using 11-0 sutures. The proximal stump was coapted to the median nerve of the injury side. The wounds were closed with interrupted 5-0 nylon suture. In the TBPI group, the rats received C5–T1nerve root avulsion, as in the CC7group, without any further surgery on the affected limb. In the control group,the rats had C5–T1nerve exposure as in the CC7group, but no further surgery was performed.
The flexor digitorum superficialis muscles were selected as a representative muscle of the median nerve (Jiang et al.,2016). Electromyography was performed in five rats per group at 3 days, 1 week, 2 weeks, 1 month, 3 months and 5 months postoperatively using Neuromatic 2000M electrophysiological apparatus (Dantec, Les Ulis, France). The CC7nerve was stimulated using an electrode placedin situ(single square wave shocks, 2.0 mA super pulse current, 0.2 ms pulse width, 1 Hz stimulus frequency). Compound muscle action potentials (CMAPs) were recorded from the flexor digitorum superficialis muscle using a concentric needle recording electrode inserted vertically into the muscles.
Sample collection
The motor cortex specimens from both sides of cortex were taken from five rats per group at 3 days, 1 week, 2 weeks, 1 month, 3 months and 5 months postoperatively.
Anesthetized rats were secured in a stereotactic frame,and the scalp was incised along the midline. Both sides of the skull at the area of the motor cortex were removed (size of craniectomy, 15 mm2) using a drill and a rongeur. The coordinates of the three points from the Bregma were 4 mm rostral/1 mm lateral (coordinate A = +4, +1), 2 mm caudal/1 mm lateral (coordinate B = −2, +1), and 4 mm rostral/6 mm lateral (coordinate C = +4, +6) (Lee et al., 2009) (Figure 1).Avoiding deep structure injury, we removed the brain to a depth of 4 mm from the outer surface of the skull. We excluded rats with deep structure injury. The specimens were stored at −80°C until use.
iTRAQ quantitative proteomic analysis
At 3 days, 1 and 2 weeks, and 1, 3 and 5 months following surgery, five injured and normal control animals were sacrificed under sodium pentobarbital anesthesia (100 mg/kg,intraperitoneally). Motor cortex tissue from the same group(n= 5 rats) was pooled together for protein extraction.The mixed tissues were rinsed to remove blood, cut into pieces and ground into powder. The tissues were vortexed for 15 minutes after the addition of lysis buffer (7 M urea,2 M sulfourea, 0.1% phenylmethyl sulfonylfluoride, 65 M dithiothreitol) and incubated on ice for 30 minutes. After centrifugation at 12,000 revolutions per minute for 15 minutes, supernatants were collected and stored at −70°C until further use. The high-abundance proteins were removed by a Removal System Affinity Column (Agilent Technologies,Santa Clara, CA, USA) according to the manufacturer’s instructions. After depletion, the protein concentration was assessed by the Bradford method (Xie et al., 2011).
Proteins of each group at each time point were labeled with iTRAQ reagents (Bio-Rad, Hercules, CA, USA). Cysteine residues were then blocked for 10 minutes at room temperature by adding 1 μL iTRAQ cysteine blocking reagent and the samples were digested with trypsin (30 μg) at 37°C overnight.
The peptides were fractionated on a Waters ultra performance liquid chromatograph using a C18 column (Waters BEH C18 2.1 × 50 mm, 1.7 μm). The fraction was separated by nano-high performance liquid chromatography(Eksigent Technologies, California, USA) on a secondary reverse-phase analytical column (Eksigent, C18, 3 μm, 150 mm × 75 μm). A Triple TOF 4600 mass spectrometer was operated in information-dependent data acquisition mode to switch automatically between mass spectrometry (MS)and MS/MS acquisition. MS spectra were acquired across the mass range of 350–1,250 m/z using 250 ms accumulation time per spectrum. Tandem mass spectra were scanned from 100–1,250 m/z in high sensitivity mode with rolling collision energy. The 25 most intense precursors were selected for fragmentation per cycle, with a dynamic exclusion time of 25 seconds.
Tandem mass spectra were extracted and charge state deconvoluted by MS Data Converter from AB Sciex. Mascot(version 2.3.02; Matrix Science, London, UK) which was only used to interpret samples. Mascot was set up to search the rat database (Acanthamoeba castellanii from NCBI,30279 entries), assuming trypsin enzyme digestion. Mascot was searched with a fragment ion mass tolerance of 0.1 Da and a parent ion tolerance of 25 ppm. Methylthio of cysteine,iTRAQ8plex of lysine, and the N-terminus were specified in Mascot as fixed modifications. Oxidation of methionine and iTRAQ8plex of tyrosine were specified in Mascot as variable modifications.
Scaffold (version Scaffold_4.3.2; Proteome Software Inc.,Portland, OR, USA) was used to validate MS/MS based peptide and protein identifications. Peptide identifications were accepted, with a false discovery rate of less than 1.0% by the Scaffold Local false discovery rate algorithm.
A technical replication was performed in the iTRAQ analysis test and the medians of the two technical replicates were used in the normalization calculation.
Bioinformatics analysis
The control group was used as the baseline. Differentially expressed proteins in the CC7group and the TBPI group were analyzed separately at each time point. Proteins in the CC7group that showed differential expression of more than 1.5 or less than 0.67 relative to the control group at any time point were selected for further analysis. Proteins in the CC7group that showed differential expression of more than 1.2 or less than 0.83 (P< 0.05) compared to those in the TBPI group at each time point were also selected for further analysis. Differentially expressed proteins selected in step 2 and step 3 were input into the DAVID network database (https://david.ncifcrf.gov/) for further analysis, including functional classification, gene ontology analysis and functional annotation clustering (Huang da et al., 2009).
Western blot assay
Western blot assay of the same samples from the proteomic analysis was performed to validate proteomic quantitation of selected proteins; 40 μg of protein from each motor cortex sample was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane. The primary antibodies were mouse monoclonal antibodies against neurofilament (1:1,000; Abcam, Cambridge, UK), and CD90/Thy1 (1:1,000; Abcam).Secondary antibodies were goat anti-mouse IgG (H + L)(1:1,000; West Grove, PA, USA) and anti-rabbit IgG (H + L)(1:1,000; West Grove). Immunoreactive signals were detected by an enhanced chemiluminescence system (GE Healthcare, London, UK).
Statistical analysis
The results of electrophysiological study were expressed as the mean ± SD. All statistical analyses were made using SPSS version 19.0 software (IBM, Armonk, NY, USA). A twotailedt-test was applied to compare the amplitude and latency of CMAP. Statistical significance was set at 0.05.
Results
CC7 nerve transfer improves neurological function in rats with total brachial plexus avulsion injury
Electrophysiological testing showed that no nascent CMAPs from the flexor digitorum superficialis muscle could be recorded in the rats of the TBPI group at any time point postoperatively. In the CC7group, nascent CMAPs could be recorded at 3 and 5 months postoperatively, but their amplitude and latency were significantly worse than those in the control group (P< 0.05;Table 1).
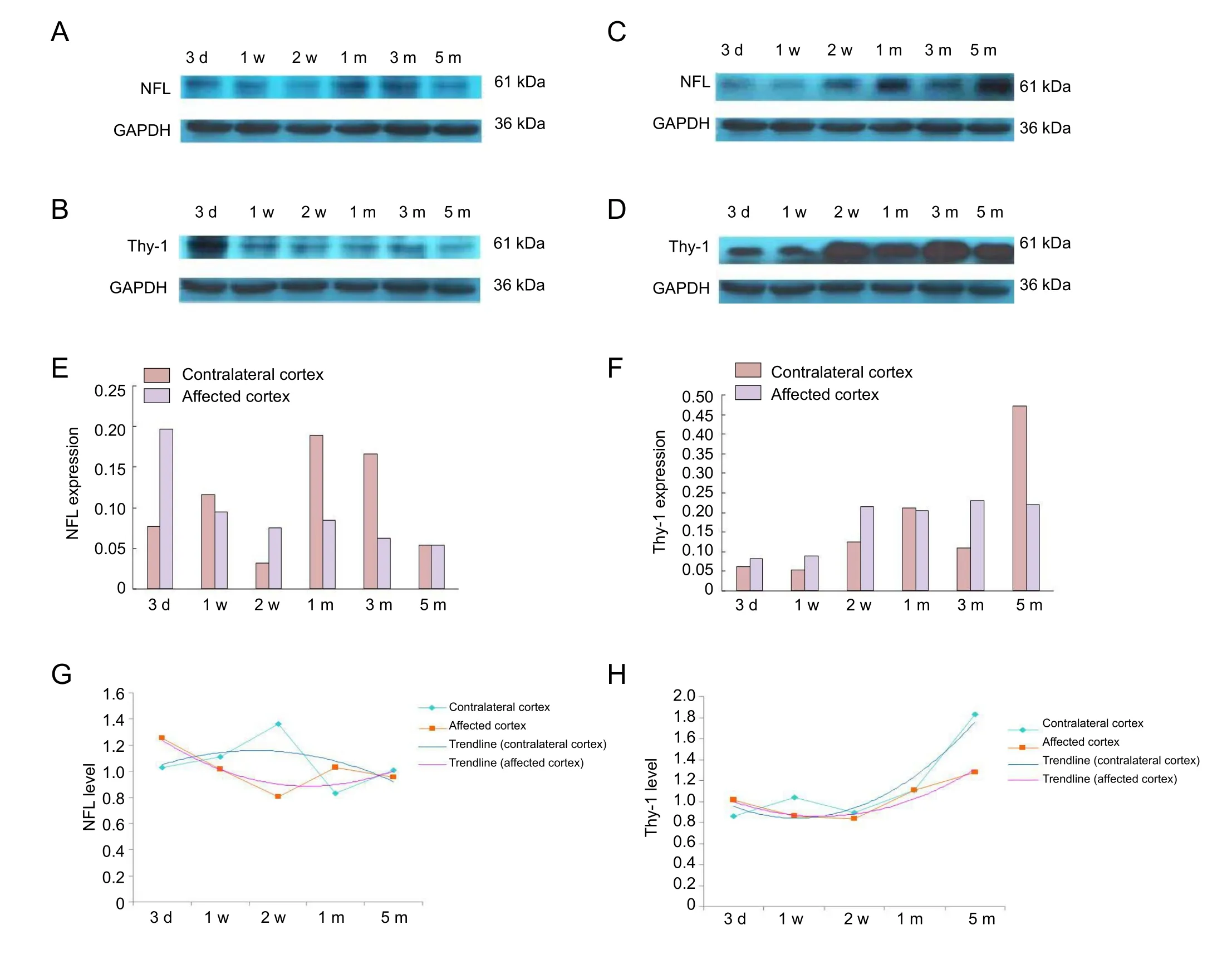
Figure 2 Proteomics results of Nfland Thy-1 in rat motor cortex after contralateral C7 nerve transfer (western blot assay).

Figure 1 Coordinates and exposure of rat motor cortex on both sides.
Changes in cortical protein expression and relationship to total brachial plexus avulsion injury with CC7 nerve transfer
Proteins in the CC7group that showed differential expression of more than 1.5 or less than 0.67 relative to control group at any time point were selected for further analysis. There were 265 differentially expressed proteins in the contralateral cortex and 264 in the affected cortex of the CC7group, as detected by iTRAQ proteome analysis. The functional classi-fication tool, gene ontology analysis and functional annotation cluster were used for further analysis.

Table 1 Effect of CC7 nerve transfer on the compound muscle action potential in the flexor digitorum superficialis muscle of rats after total brachial plexus avulsion injury
The functional classification tool of the DAVID network database can view functionally related genes together as a unit, to concentrate on the larger biological network. The differentially expressed proteins of each cortex could be classified into six function clusters (Table 2) in the gene functional classification. The differentially expressed proteins that were classified in the nervous system development and function cluster were chosen for gene ontology analysis and functional annotation clustering. We found 70 such proteins in the contralateral cortex and 71 proteins in the affected cortex in our study.
We focused on the biological process of the differentially expressed proteins to analyze their biological function. The top 10 processes in each cortex from the DAVID network database were listed according toPvalues (Table 3).
Functional annotation clustering can cluster highly similar annotated proteins into one functional group according to their co-association with genes. We listed the top three clusters of each cortex for the CC7group (Table 4).
Proteins in the CC7group which showed differential expression of more than 1.2 or less than 0.83 compared with those in the TBPI group at each time point were also selected for further analysis. We found 439 differentially expressed proteins in the contralateral cortex and 622 in the affected cortex of the CC7group, as quantified by iTRAQ proteome analysis (Table 5). Gene ontology analysis was used for further analysis.
According to electrophysiological testing, changes in proteins should be at earlier than 3 months if they participate in the nerve regeneration processes. Thus, the period before 1 month and after 1 month (including 1 month) was defined as the early stage and the late stage, respectively. The biological processes of differentially expressed proteins at each stage were analyzed together (Table 6).
According to the results of the above protein analysis, we chose differentially expressed proteins that were found in bilateral cortices and could also be classified in the nervous system development and major function cluster. Five key differentially expressed proteins were found: neurofilament light (NFL), Thy-1, Gap43, Cdc42 and Stmn1.
Among these five proteins, Nfland Thy-1 were identified for the first time by screening of peripheral nervous system injury. Compared to the TBPI group, in contralateral cortex,the Nfllevel was upregulated at 2 weeks after transfer then downregulated at 1 and 5 months. Conversely, the Thy-1 level was upregulated from 1 month to 5 months after transfer. In the affected cortex, the Nfllevel increased gradually from 1 month to 5 months after transfer.
Compared to the control group, in contralateral cortex,the Nfllevel tended to be upregulated at first, and then to be downregulated. Conversely, the Thy-1 level tended to be gradually upregulated. In the affected cortex, the Nfllevel tended to be gradually downregulated while the Thy-1 level still tended to be gradually upregulated.
Therefore, we selected these two key proteins to assess by western blot assay, the results of which were consistent with the proteomic findings (Figure 2).
Discussion
A well-functioning upper extremity requires not only a healthy peripheral nervous system, including muscular and sensory end-organs, but also a healthy central nervous system. TBPI is devastating for patients and poses substantial challenges for surgeons; nerve transfer procedures remain the primary option for nerve reconstruction after TBPI(Tung, 2014; Liu et al., 2015a). CC7is commonly transferred to the median nerve to restore hand function in TBPI patients. It provides 17,000 to 40,000 myelinated nerve fibers and contains both sensory and motor fibers, so the median nerve is a suitable recipient nerve for CC7(Gu, 2007; Tu et al., 2014). Although many variables can affect the success of repair, we wanted to study the role of the central nervous system. Multiple studies have demonstrated that remodeling of neural networks, termed brain plasticity, occurs after peripheral nerve injury and repair, especially after nerve transfer. Due to the trans-hemispheric reorganization process,choosing the CC7nerve is more challenging and requires more time for nerve regeneration (Kiper et al., 2007; Pour-rier et al., 2010; Beisteiner et al., 2011). New approaches for improving the functional results rely on fully understanding this undisclosed process. Identifying changes in proteins is helpful for this purpose.

Table 2 Gene functional classification in contralateral and affected motor cortices of total brachial plexus avulsion injury rats treated by contralateral C7 nerve transfer

Table 3 Top 10 biological processes of differentially expressed proteins in contralateral and affected motor cortex of total brachial plexus avulsion injury rats treated by contralateral C7 nerve transfer

Table 4 Functional annotation cluster analysis of contralateral and affected motor cortices of total brachial plexus avulsion injury rats treated by contralateral C7 nerve transfer
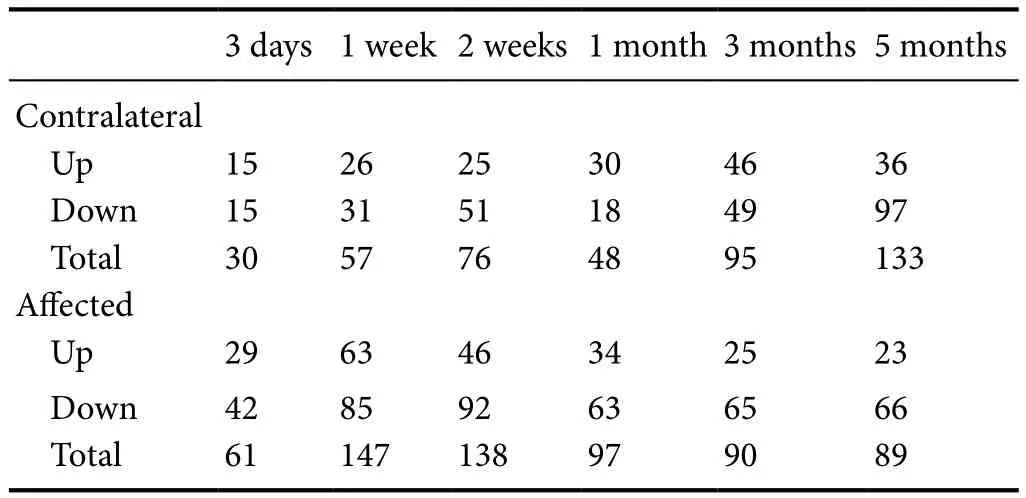
Table 5 Differential protein counts of contralateral and affected motor cortices of total brachial plexus avulsion injury rats treated by contralateral C7 nerve transfer
The functional analysis showed that most differentially expressed proteins in both cortices of the upper limb correlated with development and function of the nervous system (including neuron differentiation and development,axonogenesis and guidance), microtubule and cytoskeleton organization, synapse plasticity, and transmission of nerve impulses after CC7transfer. It implies that these processes were activated after CC7root transfer and that specific proteins were involved in the brain plasticity.
The peak of the differentially expressed protein counts appeared in the late stage in contralateral cortex and in the early stage in affected cortex. The function cluster showed that proteins involved in axonogenesis were upregulated in the early stage in contralateral cortex and the late stage in affect-ed cortex, and then downregulated in the late stage in contralateral cortex. Proteins involved in transmission of nerve impulses were upregulated in the late stage of contralateral cortex, while downregulated in affected cortex. There seemed to be a trend for alternative regulation. It has been reported that the postoperative dynamic changes for achieving functional independence of the injured limb after CC7transfer are substantially associated with the reorganization of neural control pathways (Lou et al., 2006) and perhaps these processes play an important role during brain plasticity.
Neurofilaments are major structural elements of neurons in the peripheral and central nervous systems and their major functions are to control radial growth of axons and maintain axon caliber (Hoffman et al., 1987; Kriz et al.,2000). They are composed of NFL, neurofilament medium(NFM) and neurofilament heavy (NFH), with Nflan essential component of the neurofilament core (Yuan et al., 2012).It encodes a 62 kDa structural protein, which is a major cytoskeletal component of neurons. The radial growth of myelinated axons in animals lacking neurofilaments is severely attenuated (Kuchel et al., 1997; Jordanova et al., 2003). Zhu et al. (1997) showed that after targeted disruption of the Nflgene in mice, axonal caliber is diminished and maturation of regenerating myelinated axons is delayed. Some studies have also reported that neurofilament subunits can be detected in the cerebrospinal fluid and are associated with neuronal death and axonal degeneration in Alzheimer’s disease patients (Scherling et al., 2014; Skillbäck et al., 2014).
Some studies have suggested that the cortical plasticity process is actually an activation of existing cortical pathways that are normally suppressed under healthy conditions. This plasticity process is caused by newly formed axons. Several lines of evidence show that a unilateral lesion in the motor cortex in neonatal mice can cause recrossing of corticospinal tract axons in the white matter or the gray matter of the spinal cord (Barth and Stanfield, 1990; Kuang and Kalil,1990; Joosten et al., 1992). Peripheral nerve injury results in functional expansion of neighboring cortical areas to occupy the corresponding cortical representation (Lundborg et al.,2006). A study of trans-hemispheric cortical reorganization in rats with CC7transfer to the median nerve showed that this cortical plasticity underwent two processes: early stage bilateral control and late stage single-sided control from the contralateral hemisphere (Lou et al., 2006).
In this study, the Nfllevel in the contralateral cortex was upregulated 2 weeks after transfer and then downregulated at 1 and 5 months. It indicates that Nflmay have promoted axonogenesis due to the occupation of adjacent motor cortices at 2 weeks. To regain single-sided control from the contralateral hemisphere, Nfllevels were downregulated to restrain radial axonal growth and stop the encroachment of adjacent motor cortices at 1 and 5 months. In affected cortex, the Nfllevel increased gradually from 1 to 5 months after transfer. In a positron-emission tomography study of a macaque monkey model of spinal cord injury, enhanced activity (increased regional cerebral blood flow) was observed during the early recovery stage in bilateral
M1 cortex, and in the contralesional primary motor cortex and ipsilateral ventral premotor cortex during the late recovery stage(Nishimura and Isa, 2009). Positron-emission tomography studies showed the cortical hand area in patients with incomplete spinal cord injury at the thoracic or lumbar level expanded towards the cortical leg area, with enhanced bilateral activation of the thalamus and cerebellum (Buonomano and Merzenich, 1998). One month is also the time that nascent CMAPs could be recorded from the flexor digitorum superficialis muscle. Thus, Nflpossibly promotes axonogenesis in the affected cortex to expand the cortical C7area to gain the control of affected limb, and then activates the contralateral motor cortex to gradually regain control of the affected limb.
Thy-1 is a major glycoprotein on mammalian neurons but its role in the peripheral and central nervous systems is poorly understood. Nevertheless, it has been inferred to be a negative regulator of neurite outgrowth. Chen et al. (2005)found that the action of Thy-1 may be involved in synaptic modification induced by learning or long-term potentiation(LTP).
Cortical plasticity following peripheral nerve injury includes short-term and long-term plasticity (Bitter et al., 2011).Mechanisms of cortical plasticity include activation of previously inactive connections (Jacobs and Donoghue, 1991),strengthening or weakening activity at existing synapses (Hess et al., 1996), and changes in neuronal membrane excitability(Halter et al., 1995), all of which are mainly functional.
In this study, we found that the Thy-1 level in contralateral cortex was upregulated from 1 to 5 months, indicating that Thy-1 may restrain radial axonal growth and stop the encroachment of adjacent motor cortices, the same as NFL.It may also strengthen the activity at existing synapses to enhance the connections between bilateral hemispheres.
In summary, Nfland Thy-1 may play an important rolein trans-hemispheric organization following TBPI and CC7nerve transfer. Further study focused on these two proteins is still needed to determine how and when they affect motor cortical reorganization. Regulating their expression may improve the prognosis of CC7nerve transfer.

Table 6 The major biological processes of differentially expressed proteins in affected motor cortex of total brachial plexus avulsion injury rats treated by contralateral C7 nerve transfer
Author contributions:YY and XZ conceived and designed experiments.YY and XYX performed the experiments. YY and JL analyzed sequencing data. YY wrote the paper. YY, XYX, JL and XZ reviewed and edited the paper. All authors approved the final version of the paper.
Conflicts of interest:The author declare that they have no conflicts of interest.
Financial support:None.
Research ethics:All animal protocols were performed in strict accordance with the Guide for the Care and Use of Laboratory Animals described by the U.S. National Institutes of Health and conformed to guidelines of the Society for Neuroscience and the Neurotrauma Society.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under identical terms.
Open peer review report:
Reviewer:Melanie G. Urbanchek, University of Michigan, USA.
Comments to authors:The authors state that the purpose of the experiment is to screen a portion of the brain motor cortex for differential protein expression by using iTRAQ-based quantitative proteomic technique and western blot techniques after a CC7 nerve transfer. The CC7 nerve transfer is a surgical procedure to repair total brachial plexus avulsion injury. In a rat model, the experimental group had the C5–T1 nerve roots avulsed on the affected limb and the entire root of CC7 nerve was transected and attached to the affected side median nerve using the dissected ulnar nerve of the affected side as a connecting bridge between CC7 and the affected medial nerve. Appropriate positive and negative control groups were also studied. Organizing software modes are used to classify the effects of differential proteins. It is assumed by the reviewer that historical data are used to infer the roles of protein differentiation during the motor cortex reorganization. These historical data are recalled in the discussion and used without statistical treatment to infer correlation relationship with the protein data found in the motor cortexes of the rats studied. These described correlations need to be explained and documented more thoroughly in the results section of the manuscript. Some measurements indicate that the CC7 nerve transfers successfully restored nerve connections to the affected limb.
Allieu Y, Privat JM, Bonnel F (1984) Paralysis in root avulsion of the brachial plexus. Neurotization by the spinal accessory nerve. Clin Plast Surg 11:133-136.
Barth TM, Stanfield BB (1990) The recovery of forelimb-placing behavior in rats with neonatal unilateral cortical damage involves the remaining hemisphere. J Neurosci 10:3449-3459.
Beaulieu JY, Blustajn J, Teboul F, Baud P, De Schonen S, Thiebaud JB,Oberlin C (2006) Cerebral plasticity in crossed C7grafts of the brachial plexus: an fMRI study. Microsurgery 26:303-310.
Beisteiner R, Hollinger I, Rath J, Wurnig M, Hilbert M, Klinger N,Geissler A, Fischmeister F, Wober C, Klosch G, Millesi H, Grisold W,Auff E, Schmidhammer R (2011) New type of cortical neuroplasticity after nerve repair in brachial plexus lesions. Arch Neurol 68:1467-1470.
Bitter T, Sorger B, Hesselmann V, Krug B, Lackner K, Guntinas-Lichius O (2011) Cortical representation sites of mimic movements after facial nerve reconstruction: a functional magnetic resonance imaging study. Laryngoscope 121:699-706.
Brunelli G, Monini L (1984) Neurotization of avulsed roots of brachial plexus by means of anterior nerves of cervical plexus. Clin Plast Surg 11:149-152.
Buonomano DV, Merzenich MM (1998) Cortical plasticity: from synapses to maps. Annu Rev Neurosci 21:149-186.
Chen CH, Wang SM, Yang SH, Jeng CJ (2005) Role of Thy-1 in in vivo and in vitro neural development and regeneration of dorsal root ganglionic neurons. J Cell Biochem 94:684-694.
Chuang DC, Hernon C (2012) Minimum 4-year follow-up on contralateral C7 nerve transfers for brachial plexus injuries. J Hand Surg Am 37:270-276.
Chuang ML, Chuang DC, Lin IF, Vintch JR, Ker JJ, Tsao TC (2005)Ventilation and exercise performance after phrenic nerve and multiple intercostal nerve transfers for avulsed brachial plexus injury.Chest 128:3434-3439.
Donoghue JP, Suner S, Sanes JN (1990) Dynamic organization of primary motor cortex output to target muscles in adult rats. II. Rapid reorganization following motor nerve lesions. Exp Brain Res 79:492-503.
Gu YD (2007) Contralateral C7 root transfer over the last 20 years in China. Chin Med J (Engl) 120:1123-1126.
Gu YD, Ma MK (1996) Use of the phrenic nerve for brachial plexus reconstruction. Clin Orthop Relat Res:119-121.
Gu YD, Chen DS, Zhang GM, Cheng XM, Xu JG, Zhang LY, Cai PQ,Chen L (1998) Long-term functional results of contralateral C7 transfer. J Reconstr Microsurg 14:57-59.
Halter JA, Carp JS, Wolpaw JR (1995) Operantly conditioned motoneuron plasticity: possible role of sodium channels. J Neurophysiol 73:867-871.
Hess G, Aizenman CD, Donoghue JP (1996) Conditions for the induction of long-term potentiation in layer II/III horizontal connections of the rat motor cortex. J Neurophysiol 75:1765-1778.
Hoffman PN, Cleveland DW, Griffin JW, Landes PW, Cowan NJ, Price DL (1987) Neurofilament gene expression: a major determinant of axonal caliber. Proc Natl Acad Sci U S A 84:3472-3476.
Huang da W, Sherman BT, Lempicki RA (2009) Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc 4:44-57.
Jacobs KM, Donoghue JP (1991) Reshaping the cortical motor map by unmasking latent intracortical connections. Science 251:944-947.
Jiang S, Li ZY, Hua XY, Xu WD, Xu JG, Gu YD (2010) Reorganization in motor cortex after brachial plexus avulsion injury and repair with the contralateral C7 root transfer in rats. Microsurgery 30:314-320.
Jiang Y, Wang L, Lao J, Zhao X (2016) Comparative study of intercostal nerve transfer to lower trunk and contralateral C7root transfer in repair of total brachial plexus injury in rats. J Plast Reconstr Aesthet Surg 69:623-628.
Joosten EA, Schuitman RL, Vermelis ME, Dederen PJ (1992) Postnatal development of the ipsilateral corticospinal component in rat spinal cord: a light and electron microscopic anterograde HRP study. J Comp Neurol 326:133-146.
Jordanova A, De Jonghe P, Boerkoel CF, Takashima H, De Vriendt E, Ceuterick C, Martin JJ, Butler IJ, Mancias P, Papasozomenos S,Terespolsky D, Potocki L, Brown CW, Shy M, Rita DA, Tournev I,Kremensky I, Lupski JR, Timmerman V (2003) Mutations in the neurofilament light chain gene (NEFL) cause early onset severe Charcot-Marie-Tooth disease. Brain 126:590-597.
Kaas J, Krubitzer L, Chino Y, Langston A, Polley E, Blair N (1990)Reorganization of retinotopic cortical maps in adult mammals after lesions of the retina. Science 248:229-231.
Kiper D, Martin KAC, Scherberger H (2007) Cortical plasticity: a view from nonhuman primates. Neurodegener Dis 4:34-42.
Krakauer JD, Wood MB (1994) Intercostal nerve transfer for brachial plexopathy. J Hand Surg Am 19:829-835.
Kriz J, Zhu Q, Julien JP, Padjen AL (2000) Electrophysiological properties of axons in mice lacking neurofilament subunit genes: disparity between conduction velocity and axon diameter in absence of NF-H.Brain Res 885:32-44.
Kuang RZ, Kalil K (1990) Specificity of corticospinal axon arbors sprouting into denervated contralateral spinal cord. J Comp Neurol 302:461-472.
Kuchel GA, Poon T, Irshad K, Richard C, Julien JP, Cowen T (1997) Decreased neurofilament gene expression is an index of selective axonal hypotrophy in ageing. Neuroreport 8:799-805.
Lang EM, Schlegel N, Reiners K, Hofmann GO, Sendtner M, Asan E(2008) Single-dose application of CNTF and BDNF improves remyelination of regenerating nerve fibers after C7ventral root avulsion and replantation. J Neurotrauma 25:384-400.
Lee DH, Hong SH, Kim SK, Lee CS, Phi JH, Cho BK, Wang KC (2009)Reproducible and persistent weakness in adult rats after surgical resection of motor cortex: evaluation with limb placement test. Childs Nerv Syst 25:1547-1553.
Liu GY, Jin Y, Zhang Q, Li R (2015a) Peripheral nerve repair: a hot spot analysis on treatment methods from 2010 to 2014. Neural Regen Res 10:996-1002.
Liu Y, Xu XC, Zou Y, Li SR, Zhang B, Wang Y (2015b) Phrenic nerve transfer to the musculocutaneous nerve for the repair of brachial plexus injury: electrophysiological characteristics. Neural Regen Res 10:328-333.
Lou L, Shou T, Li Z, Li W, Gu Y (2006) Transhemispheric functional reorganization of the motor cortex induced by the peripheral contralateral nerve transfer to the injured arm. Neuroscience 138:1225-1231.
Lundborg G, Waites A, Bjorkman A, Rosen B, Larsson EM (2006)Functional magnetic resonance imaging shows cortical activation on sensory stimulation of an osseointegrated prosthetic thumb. Scand J Plast Reconstr Surg Hand Surg 40:234-239.
Mathews AL, Yang G, Chang KW, Chung KC (2017) A systematic review of outcomes of contralateral C7transfer for the treatment of traumatic brachial plexus injury: an international comparison. J Neurosurg 126:922-932.
Merzenich MM, Jenkins WM (1993) Reorganization of cortical representations of the hand following alterations of skin inputs induced by nerve injury, skin island transfers, and experience. J Hand Ther 6:89-104.
Nagano A, Yamamoto S, Mikami Y (1995) Intercostal nerve transfer to restore upper extremity functions after brachial plexus injury. Ann Acad Med Singapore 24:42-45.
Narakas A (1978) Surgical treatment of traction injuries of the brachial plexus. Clin Orthop Relat Res:71-90.
Navarro X, Vivó M, Valero-Cabré A (2007) Neural plasticity after peripheral nerve injury and regeneration. Prog Neurobiol 82:163-201.
Nicolelis MA, Lin RC, Woodward DJ, Chapin JK (1993) Induction of immediate spatiotemporal changes in thalamic networks by peripheral block of ascending cutaneous information. Nature 361:533-536.
Nishimura Y, Isa T (2009) Compensatory changes at the cerebral cortical level after spinal cord injury. Neuroscientist 15:436-444.
Pan F, Wei HF, Chen L, Gu YD (2012) Different functional reorganization of motor cortex after transfer of the contralateral C7to different recipient nerves in young rats with total brachial plexus root avulsion. Neurosci Lett 531:188-192.
Pourrier SD, Nieuwstraten W, Van Cranenburgh B, Schreuders TA,Stam HJ, Selles RW (2010) Three cases of referred sensation in traumatic nerve injury of the hand: implications for understanding central nervous system reorganization. J Rehabil Med 42:357-361.
Qi HX, Stepniewska I, Kaas JH (2000) Reorganization of primary motor cortex in adult macaque monkeys with long-standing amputations. J Neurophysiol 84:2133-2147.
Sanes JN, Suner S, Donoghue JP (1990) Dynamic organization of primary motor cortex output to target muscles in adult rats. I. Longterm patterns of reorganization following motor or mixed peripheral nerve lesions. Exp Brain Res 79:479-491.
Scherling CS, Hall T, Berisha F, Klepac K, Karydas A, Coppola G,Kramer JH, Rabinovici G, Ahlijanian M, Miller BL, Seeley W, Grinberg LT, Rosen H, Meredith J Jr, Boxer AL (2014) Cerebrospinal fluid neurofilament concentration reflects disease severity in frontotemporal degeneration. Ann Neurol 75:116-126.
Seddon HJ (1963) Nerve grafting. J Bone Joint Surg Br 45:447-461.
Skillbäck T, Farahmand B, Bartlett JW, Rosen C, Mattsson N, Nagga K,Kilander L, Religa D, Wimo A, Winblad B, Rosengren L, Schott JM,Blennow K, Eriksdotter M, Zetterberg H (2014) CSF neurofilament light differs in neurodegenerative diseases and predicts severity and survival. Neurology 83:1945-1953.
Sokki AM, Bhat DI, Devi BI (2012) Cortical reorganization following neurotization: a diffusion tensor imaging and functional magnetic resonance imaging study. Neurosurgery 70:1305-1311.
Songcharoen P, Wongtrakul S, Mahaisavariya B, Spinner RJ (2001)Hemi-contralateral C7transfer to median nerve in the treatment of root avulsion brachial plexus injury. J Hand Surg Am 26:1058-1064.
Tu YK, Tsai YJ, Chang CH, Su FC, Hsiao CK, Tan JS (2014) Surgical treatment for total root avulsion type brachial plexus injuries by neurotization: a prospective comparison study between total and hemicontralateral C7nerve root transfer. Microsurgery 34:91-101.
Tung TH (2014) Nerve transfers. Clin Plast Surg 41:551-559.
Wei HF, Zeng BF, Chen YF, Chen L, Gu YD (2011) BDNF and GAP43 contribute to dynamic transhemispheric functional reorganization in rat brain after contralateral C7root transfer following brachial plexus avulsion injuries. Neurosci Lett 500:187-191.
Xie XZ, Wang X, Liu LH, Dong SL, Pi XE, Liu W (2011) ITRAQ Technology and Its Application in Proteomics. Zhongguo Shengwu Huaxue yu Fenzi Shengwu Xuebao 27:616-621.
Yu A, Wang S, Cheng X, Liang W, Bai R, Xue Y, Li W (2017) Functional connectivity of motor cortical network in patients with brachial plexus avulsion injury after contralateral cervical nerve transfer: a resting-state fMRI study. Neuroradiology 59:247-253.
Yuan A, Rao MV, Veeranna, Nixon RA (2012) Neurofilaments at a glance. J Cell Sci 125:3257-3263.
Zhao J, Xu Q (2008) Over-expression of gap-43 in mn9d cells induces spontaneous sprouting and causes susceptibility to semaphorin3a.Int J Dev Neurosci 26:866.
Zhu Q, Couillard-Despres S, Julien JP (1997) Delayed maturation of regenerating myelinated axons in mice lacking neurofilaments. Exp Neurol 148:299-316.

- 中国神经再生研究(英文版)的其它文章
- Detection of thinned corticospinal tract and corticoreticular pathway in a patient with a calf circumference discrepancy
- Relationship of distraction rate with inferior alveolar nerve degeneration-regeneration shift
- The effect of increased intra-abdominal pressure on orbital subarachnoid space width and intraocular pressure
- Voltage adjustment improves rigidity and tremor in Parkinson’s disease patients receiving deep brain stimulation
- Effect of electrical stimulation on neural regeneration via the p38-RhoA and ERK1/2-Bcl-2 pathways in spinal cord-injured rats
- GSK3β inhibitor promotes myelination and mitigates muscle atrophy after peripheral nerve injury