
云南松天然群体球果表型变异研究
2018-03-12 07:00:48,,,,,
种子 2018年1期
,, ,,,
(1.西南林业大学,云南省高校林木遗传改良与繁育重点实验室,昆明 650224;2.北京林业大学,林木育种国家工程实验室,北京 100083)
云南松(PinusyunnanensisFranch.)是我国云贵高原的主要乡土树种[1-3],具有适应性强、耐干旱瘠薄、木材用途广等特点,在分布区域林业生产和生态经济建设中占有重要的地位[3-5],天然分布于东经96°~108°、北纬23°~30°之间,分布范围广,生境复杂多样[6],其中云南省是云南松的集中分布区[3,7-8]。目前云南松现存林分低矮、弯曲、扭曲等不良个体的比例逐渐增加,表现出林分衰退问题日益突出现象,亟需对云南松种质资源开展保护工作,而遗传多样性研究是开展种质资源保护的基础[9]。
表型多样性是遗传多样性与环境多样性的综合体现,主要研究群体在其分布区内各种环境下的表型变异,是遗传多样性和生物系统学研究的重要内容[10-11],其表型性状形态学结构特征也是植物遗传多样性研究形态学标记的重要内容[12-16],同时通过表型来检测群体的变异和分化又是一种简便、快捷、有效的方法[11,17],特别是在短期内对种质资源进行评价时,常作为一种重要的评价手段。球果作为裸子植物繁殖系统的重要特征[18],在选择压力下会发生适应性演化[19-20],且果实形态往往是较稳定的遗传特征[20],其表型性状不仅决定物种的扩散能力,也影响到物种的萌发和幼苗定居,进而影响到种群的分布格局[21]。云南松种实性状在不同海拔梯度群体或不同茎干类型群体的相关研究中初步表明存在一定的变异[22-24],但针对不同分布区域群体间的球果表型变异未见研究报道。鉴于此,本研究在前期针叶表型变异研究基础上[25],以云南松主分布区不同分布区域天然群体的球果为材料,分析云南松群体间和群体内球果性状的表型变异程度与变异规律,以期为云南松资源保护与合理利用提供依据。
1 材料与方法
1.1 群体的分布及球果的采集
在云南松自然分布区内选择天然群体9个(表1),样株的年龄均在25年生以上[14]。为了最大化地代表各群体的遗传变异状况,采集的样品均来自于不同植株,且各样株不相邻,间隔5倍以上树高[23],从树冠南向中上部采集当年成熟的球果,用于表型性状测定。
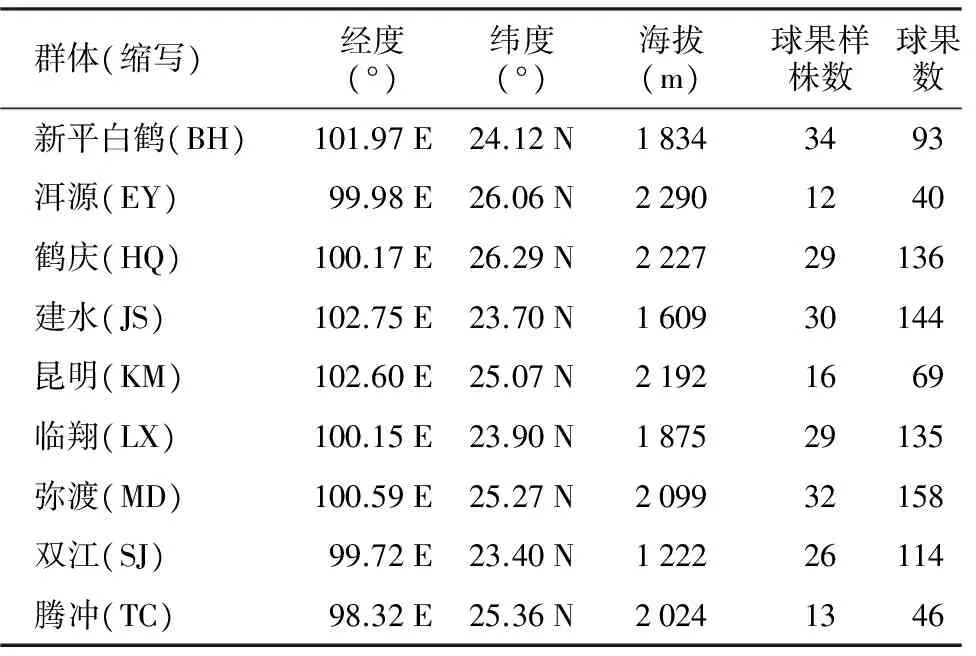
表1 云南松天然群体信息[23]
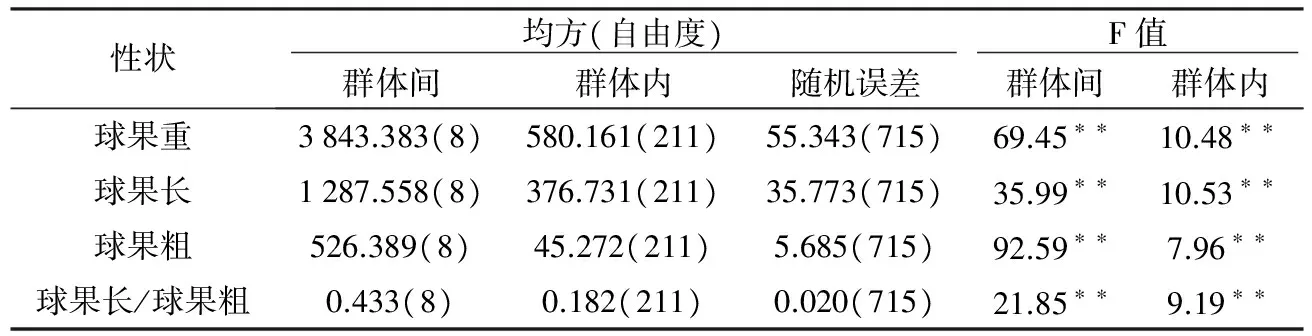
表2 云南松球果性状的方差分析
注:**表示在0.01%水平上差异显著。
1.2 球果性状的测定
包括球果长、球果粗、球果重量、球果长/球果粗等4个性状。用电子游标卡尺测量球果长(两端的直线长度)和球果粗(最宽处直径)[15],精确到0.01 mm,用电子天平测量球果重,精确到0.01 g,计算球果长/球果粗,即球果的形状指数,精确到0.01,每单株测定5~10个球果。
1.3 土壤样品的采集与测定
土壤样品的采集与测定同前期研究所述[25],分别测定含水量、pH值、有机质、水解氮、速效磷、速效钾、全氮、全磷、全钾及质地共10个指标,其中质地采用定性描述,未用于后续的分析中。
1.4 数据分析
对球果性状采用巢式方差分析[10,26-27]计算群体间和群体内的方差分量,线性模型为:
Yijk=μ+τi+δj(i)+εk(ij),
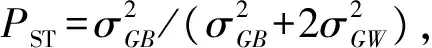
2 结果与分析
2.1 群体间和群体内的球果性状变异特征
将4个球果表型性状进行巢式方差分析,各性状的均方及F值见表2。云南松4个球果性状在群体间和群体内均呈极显著差异(p<0.01),表明云南松的球果表型性状在群体间和群体内存在广泛的变异[23]。
2.2 球果性状群体间的形态分化
对各个性状成对群体间进一步分析形态分化PST(表3和图1)。球果重、球果长、球果粗和球果长/球果粗在各成对群体间的形态分化波动于0.000 0~0.462 9、0.000 0~0.213 1、0.000 0~0.605 7和0.000 0~0.145 5,平均分别为0.124 1、0.051 5、0.162 1和0.026 4,以球果长/球果粗有形态分化最低,即该性状最稳定。总体来看,球果性状在各群体间的分化均较低,其中仅有一对群体间在球果粗方面表现出分化系数大于0.5。
2.3 球果性状的变异特征
对同一群体的不同性状或同一性状的不同群体进行比较[23],9个群体球果表型性状的相对极差见表4。云南松球果重、球果长、球果粗和球果长/球果粗的相对极差波动于0.42~0.96、0.43~0.90、0.46~1.00和0.31~0.96,平均分别为0.66、0.71、0.61和0.62。综合各群体的相对极差来看,BH和MD群体变异程度比较高,而TC群体的变异比较低。
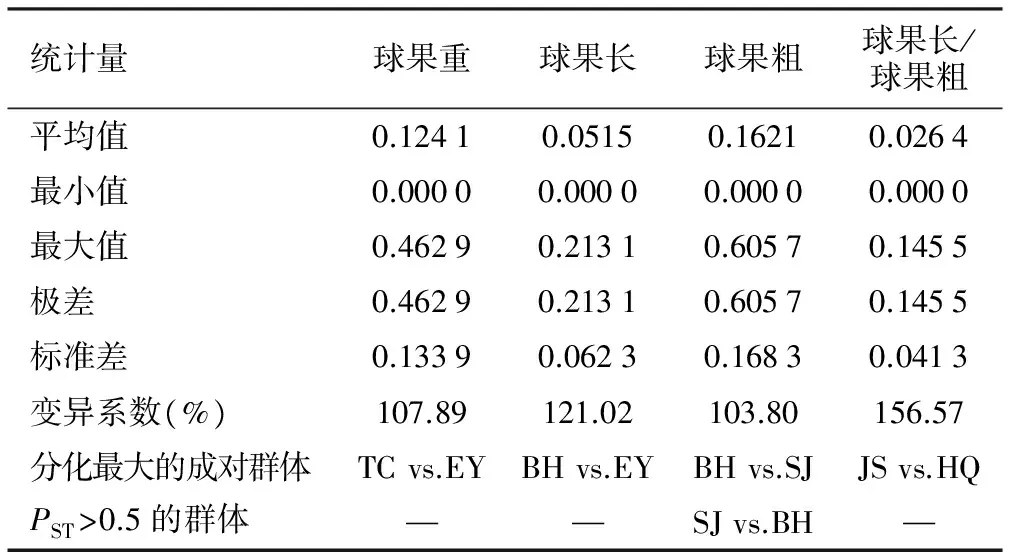
表3 云南松天然群体球果性状的形态分化(PST)
表4 云南松天然群体球果性状的相对极差
注:群体名称的缩写见表1。
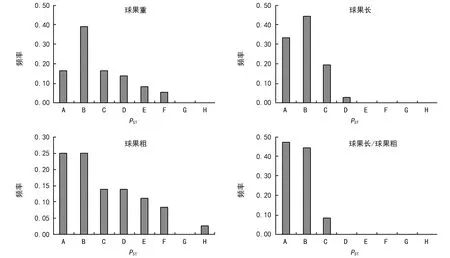
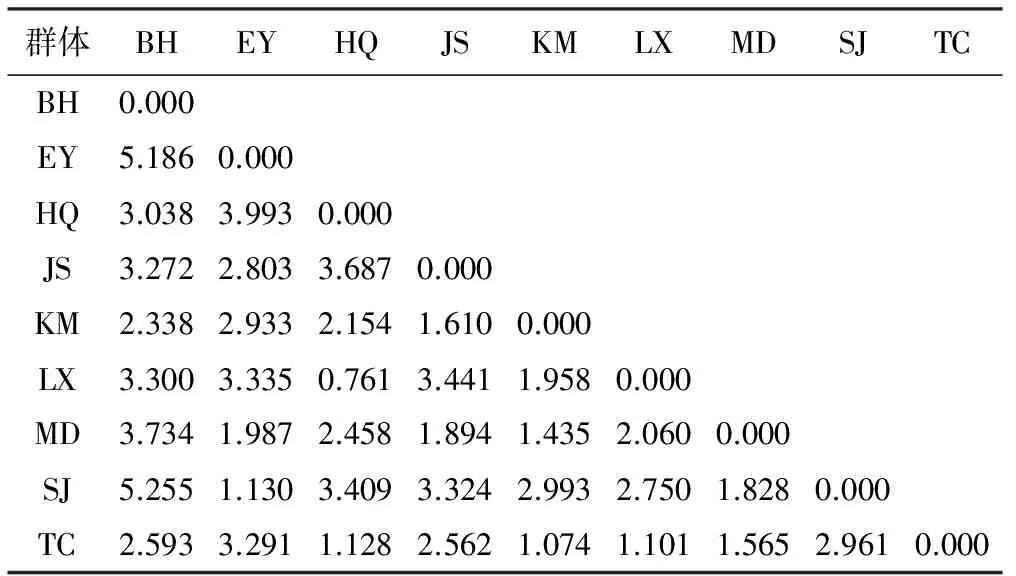
注:A:PST=0;B:0 对云南松球果性状与土壤因子间相关分析表明(表5),除球果重与有机质含量、全氮含量,及球果长/球果粗与全钾含量之间的相关性为显著正相关关系外(p<0.05),其它因子间的相关性均未达到显著水平,相关性弱,变异可能是随机的分布[23]。 表5 云南松天然群体球果性状与土壤因子间的相关分析 (Spearman’s) 注:*表示在0.05%水平上相关性显著。 基于云南松4个球果性状,对9个群体进行主成分分析,结果见表6和图2。第一主成分主要解释球果长、球果粗和球果重,即球果的大小及其重量;第二主成分主要解释球果长/球果粗,即球果的形状。其中第一主成分将BH群体明显区分于其它群体,第三主成分使MD与其它群体区分开来[23]。 将4个球果性状在各群体的均值标准化后,计算各群体间的欧式距离,结果见表7。各距离波动于0.761~5.255之间,以BH群体和SJ群体间的距离最大,而以HQ群体和LX群体间的距离最小。 表6 云南松天然群体球果性状的主成分分析 表7 基于球果性状云南松天然群体间的欧式距离 注:群体名称的缩写见表1。 图2 基于球果性状云南松天然群体的主成分分析三维图 图3 基于球果性状云南松天然群体的UPGMA聚类图 基于球果性状各成对群体间的欧氏距离,绘制云南松9个群体的UPGMA聚类图(图3),结果表明,当以欧氏距离2.8为阈值时,9个群体被分为3大类:其中BH单独形成一类(Ⅰ),SJ和EY聚成另一类(Ⅱ),其余6个群体形成第三类(Ⅲ)。当以欧氏距离1.5为阈值时,可将第三大类再分为3个亚类:LX和HQ 2个群体形成的第1亚类(Ⅲ-1),KM、TC和MD 3个群体构成的第2亚类(Ⅲ-2),JS独自形成一个亚类(Ⅲ-3)[23]。 遗传变异是群体生物逐渐适应环境和不断进化的产物,是遗传信息的重要表征[35-36]。云南松球果表型性状在群体间和群体内均存在极显著差异,在白皮松[10]、川西云杉[37]、云杉[38-40]、油松[41]、红松[42]、华北落叶松[43]等研究中均揭示表型性状变异的广泛性,广泛的形态变异能适应不同环境[44]。不同海拔梯度云南松群体针叶、种实等性状多样性指数分析表明,云南松群体表现出丰富的表型多样性[22,45]。此外,在不同地理分布区域群体间、不同茎干类型表型多样性的研究均揭示云南松表型变异丰富,各表型性状表现出不一样的变异式样[23-25,46-48]。丰富的表型变异是云南松适应性广、抗逆性强的基础[25]。云南松分布范围广,且为异交的多年生植物,这些特性都有利于产生丰富的变异,从而适应不同环境条件[23,25]。 各球果性状在群体间存在极显著差异,存在一定的形态分化,球果重、球果长、球果粗和球果长/球果重的形态分化分别为0.124 1、0.051 5、0.162 1和0.026 4,以球果粗和球果重的较高,总体来看,球果性状形态分化都较低,群体内变异是云南松球果变异的主要来源[23,25,49],表型变异分析与遗传多样性分析一致[49]。在白皮松[10]、云杉[40]、川西云杉[37]、油松[41]、青海云杉[38-39]等研究中均揭示变异主要存在于群体内。这可能与繁育系统有关,云南松为风媒异花授粉植物,种子具翅,花粉具气囊,可远距离扩散[23],从而导致群体间的基因流较大[49],削弱群体间的遗传分化,从而使群体间的变异降低。 不同性状群体内的变异程度有所差异,球果重的变异大于球果形状大小的变异,其中又以球果长的变异高于球果粗的变异,云南松球果大小在纵向上的变异较横向上的变异大,以球果粗最为稳定,而球果重的遗传变异最为丰富。对红松的研究也报道相似的结果,张恒庆等对不同纬度和海拔下3个生境的天然红松林研究表明,3个群体的变异系数均表现为球果重>球果长>球果粗,平均变异系数分别为30.77%、14.94%和10.60%,群体内以球果粗最为稳定[42]。研究还表明,云南松各球果性状比针叶性状变异幅度小[25],即球果性状比针叶性状稳定,这可能是因为叶子性状更容易受环境条件的影响[14],相反,繁殖性状更保守不易受环境条件的影响[50],果实大小的性状更为稳定[20]。此外,不同群体的各个球果性状的变异也不一样,在9个群体中,BH和MD群体变异程度比较高,而TC群体的变异比较低。不同群体的相同性状变异程度存在差异,这可能是由环境异质性导致的[20]。 球果性状与各土壤因子相关不显著或弱相关,变异可能是随机的分布。类似的如油松[41]、白皮松[10]、川西云杉[37]、华北落叶松[43]等研究中发现,球果性状地理变异趋势不明显。聚类分析表明,有些地理上相近的群体并没有优先聚合成一类,在针叶性状的分析中也表明,针叶性状随地理分布的变异趋势不明显,而与其生态因子分异间的关系更明显[25],这可能与群体间立地条件类型存在差异有关[20],即基于球果性状的云南松群体并不完全按地理分布进行聚类,不宜利用地理分布区域对云南松球果表型进行划分。 综上所述,云南松9个群体球果性状在群体间和群体内均存在极显著差异,表明变异存在的广泛性。各个性状的变异主要存在于群体内,开展种质资源收集与保护时,可适当增加群体内个体数、减少群体取样数。由各个球果性状形态分化较低,两两群体间的形态分化说明群体间的分化较弱。相关分析表明,各球果性状与土壤因子间的相关性较弱,变异可能是随机的。基于球果性状的主成分分析和聚类分析表明,不同群体间的聚类并不完全按照地理分布聚类。 [1]中国科学院昆明植物研究所编著.云南植物志(第四卷):种子植物[M].北京:科学出版社,1986:54-57. [2]中国科学院中国植物志编辑委员会.中国植物志(第7卷)[M].北京:科学出版社,1978:122-282. [3]金振洲,彭鉴.云南松[M].昆明:云南科技出版社,2004:1-66. [4]邓喜庆,皇宝林,温庆忠,等.云南松林资源动态研究[J].自然资源学报,2014,29(8):1 411-1 419. [5]Zhang L,Xu WH,Ouyang ZY,et al.Determination of priority nature conservation areas and human disturbances in the Yangtze River Basin,China[J].Journal for Nature Conservation,2014,22(4):326-336. [6]李本德,刘中天.云南松(Pinusyunnanensis)林的分布及其分区[J].云南大学学报,1984(1):33-46. [7]陈飞,王健敏,陈晓鸣,等.基于Kira指标的云南松气候适宜性分析[J].林业科学研究,2012,25(5):576-581. [8]陈飞,王健敏,孙宝刚,等.云南松的地理分布与气候关系[J].林业科学研究,2012,25(2):163-168. [9]SevΙk H,Ayan S,Turna I,et al.Genetic diversity among populations in Scotch pine (PinussilvestrisL.)seed stands of Western Black Sea Region in Turkey[J].African Journal of Biotechnology,2010,9(43):7 266-7 272. [10]李斌,顾万春,卢宝明.白皮松天然群体种实性状表型多样性研究[J].生物多样性,2002,10(2):181-188. [11]贾子瑞,张守攻,王军辉.林芝云杉天然群体针叶与种实的变异及其地理趋势[J].林业科学研究,2011,24(4):428-436. [12]廖柏勇,陈丽君,王芳,等.苦楝种源果核和种子性状地理变异的趋势面分析[J].林业科学研究,2016,29(5):784-792. [13]Boulli A,Baaziz M,M’Hirit O.Polymorphismof natural populations ofPinushalepensisMill.in Morocco as revealed by morphological characters[J].Euphytica,2001,119(3):309-316. [14]Dangasuk OG,Panetsos KP.Altitudinal and longitudinal variations inPinusbrutia(Ten.)of Crete Island,Greece:some needle,cone and seed traits under natural habitats[J].New Forest,2004,27(3):269-284. [15]Donahue JK,Upton JL.Geographical variation in leaves,cone and seeds of Pinus gregii in native forests[J].Forest Ecology and Management,1996,82(1):145-157. [16]Wahid N,González-Martínez SC,El Hadrami I,et al.Variation of morphological traits in natural populations of maritime pine (PinuspinasterAit.) in Morocco[J].Annals of forest Science,2006,63(1):83-92. [17]尚帅斌,郭俊杰,王春胜,等.海南岛青梅天然居群表型变异[J].林业科学,2015,51(2):154-162. [18]刘贵峰,臧润国,刘华,等.天山云杉种子形态性状的地理变异[J].应用生态学报,2012,23(6):1 455-1 461. [19]杨玲,沈海龙,梁立东,等.不同产区野生花楸果实和种子的表型多样性[J].东北林业大学学报,2009,37(2):8-10. [20]董昕,王磊,鲁仪增,等.山东稀有植物小果白刺天然群体表型变异研究[J].林业科学研究,2017,30(2):293-299. [21]李帅锋,苏建荣,刘万德,等.思茅松天然群体种实表型变异[J].植物生态学报,2013,37(11):998-1 009. [22]徐杨,邓丽丽,周丽,等.云南松不同海拔天然群体种实性状表型多样性研究[J].种子,2015,34(11):70-74,79. [23]许玉兰.云南松天然群体遗传变异研究[D].北京林业大学,2015. [24]邓丽丽,朱霞,和润喜,等.云南松不同茎干类型种实性状表型多样性比较[J].种子,2017,36(3):4-9. [25]许玉兰,蔡年辉,陈诗,等.基于针叶性状云南松天然群体表型分化研究[J].西南林业大学学报,2016,36(5):1-9. [26]葛颂,王明麻,陈岳武.用同工酶研究马尾松群体的遗传结构[J].林业科学,1998,24(11):399-409. [27]刘永红,高桂琴,崔嵬,等.油松天然群体种实性状表型多样性分析[J].种子,2010,29(9):44-49. [28]López R,López de Heredia U,Collada C,et al.Vulnerability to cavitation,hydraulic efficiency,growth and survival in an insular pine (Pinuscanariensis)[J].Annals of Botany,2013,111(6):1 167-1 179. [29]Raeymaekers JAM,Van Houdt JKJ,Larmuseau MHD,et al.Divergent selection as revealed by PST and QTL-based FST in three-spined stickleback(Gasterosteus aculeatus)populations along a coastal-inland gradient[J].Molecular Ecology,2007,16(4):891-905. [30]Leinonen T,O'hara RB,Cano JM,et al.Comparative studies of quantitative trait and neutral marker divergence:a meta-analysis[J].Journal of Evolutionary Biology,2008,21(1):1-17. [31]Spitze K.Population-structure in Daphnia obtusa:quantitative genetic and allozymic variation[J].Genetics,1993,135(2):367-374. [32]Boratyńska K,Jasińska AK,Ciepuch E.Effect of tree age on needle morphology and anatomy of Pinus uliginosa and Pinus silvestris-species-specific character separation during ontogenesis[J].Flora,2008,203(8):617-626. [33]Steel RG,Torrie JH,Dickey DA.Principles and procedures of statistics.New York-Toronto-London:McGrow-Hill Book Company Inc.,1960:481. [34]Rohlf F J.NTSYS-PC:Numerical Taxonomy and Multivariate Analysis System Version 1.80.Setauket New York:Distribution by Exeter SoftWare.1994. [35]张振,张含国,周宇,等.红松多无性系群体的种实性状变异研究[J].北京林业大学学报,2015,37(2):67-78. [36]柴春山,芦娟,蔡国军,等.文冠果人工种群的果实表型多样性及其变异[J].林业科学研究,2013,26(2):181-191. [37]辜云杰,罗建勋,吴远传,等.川西云杉天然种群表型多样性[J].植物生态学报,2009,33(2):291-301. [38]王娅丽,李毅,陈晓阳.祁连山青海云杉天然群体的种实性状遗传多样性研究[J].林业科学,2008,44(2):70-77. [39]王娅丽,李毅.祁连山青海云杉天然群体的种实性状表型多样性[J].植物生态学报,2008,32(2):355-362. [40]罗建勋,顾万春.云杉天然群体表型多样性研究[J].林业科学,2005,41(2):66-83. [41]刘永红,高桂琴,崔嵬,等.油松天然群体种实性状表型多样性分析[J].种子,2010,29(9):44-49. [42]张恒庆,安利佳,祖元刚.天然红松种群形态特征地理变异的研究[J].生态学报,1999,19(6):932-938. [43]李文荣,齐力旺,韩有志.山西华北落叶松天然林的地理分布和种群变异规律的研究[J].林业科学,1992,28(6):493-501. [44]Ji M,Zhang Q,Deng J,et al.Intra-versus inter-population variation of cone and seed morphological traits ofPinustabulaeformisCarr.in northern China:impact of climate-related conditions[J].Polish Journal of Ecology,2011,59(4):717-727. [45]徐杨,周丽,蔡年辉,等.云南松不同海拔群体的针叶性状表型多样性研究[J].云南农业大学学报(自然科学),2016,31(1):109-114. [46]Xu YL,Woeste K,Cai NH,et al.Variation in needle and cone traits in natural populations ofPinusyunnanensis[J].Journal of Forestry Research,2016,27(1):41-49. [47]邓丽丽,孙琪,许玉兰,等.云南松不同茎干类型群体针叶性状表型多样性比较[J].西南林业大学学报,2016,36(3):30-37. [48]邓丽丽,周丽,蔡年辉,等.基于针叶性状的云南松不同茎干类型遗传变异分析[J].西南农业学报,2017,30(3):530-534. [49]Xu YL,Cai NH,Woeste K,et al.Genetic diversity and population structure ofPinusyunnanensisby simple sequence repeat markers[J].Forest Science,2016,62(1):38-47. [50]Vasudeva R,Hanumantha M,Gunaga RP.Genetic variation for floral traits among teak (TectonagrandisLinn.f.) clones:Implications to seed orchard fertility[J].Current Science,2004,87(3):358-362.2.4 球果性状与土壤因子间的相关性
2.5 基于球果性状云南松天然群体的聚类



3 讨论与结论
猜你喜欢
广西植物(2023年7期)2023-08-20 06:03:03
新农业(2022年7期)2022-05-03 10:05:58
浙江农林大学学报(2022年1期)2022-03-01 07:12:16
凯里学院学报(2021年3期)2021-07-16 06:29:42
林产工业(2020年8期)2020-08-31 06:58:26
广西林业科学(2019年2期)2019-07-25 02:28:46
乡村科技(2019年9期)2019-02-22 06:09:22
山西农业科学(2019年10期)2019-02-12 04:15:51
中国林副特产(2019年3期)2019-01-10 17:17:11
福建林业(2018年3期)2018-11-07 05:34:36
