
禽呼肠病毒在鸡胚成纤维细胞系中的增殖规律
2018-02-10 02:13沈文康谢芝勋谢丽基黄娇玲曾婷婷张艳芳张民秀罗思思谢志勤邓显文
动物医学进展 2018年1期
沈文康,谢芝勋,王 盛,黄 莉,谢丽基,黄娇玲,曾婷婷,张艳芳,张民秀,罗思思,范 晴,谢志勤,邓显文
(1.广西大学动物科学技术学院,广西南宁 530004;2.广西壮族自治区兽医研究所/广西兽医生物技术重点实验室,广西南宁 530001)
禽呼肠病毒(Avian reovirus,ARV)是呼肠病毒科、正呼肠病毒属的成员,为双链RNA病毒,无囊膜,含10个节段,双衣壳结构,可感染鸡、火鸡、鹅、鸭及一些鸟类[1],临床主要症状包括病毒性关节炎、腱鞘炎、肠道病、矮小综合征、吸收不良综合征及免疫抑制等,是家禽养殖业中一类重要的传染病,严重危害着养殖业的健康发展[2]。
ARV于1954年由Fahey和Crawley从病鸡中分离得到[3];Olson等于1957年报道鸡群中有滑膜炎并分离出病原,1972年被确定为禽呼肠病毒[4]。自20世纪80年代以来,我国各省份及地区陆续报道了ARV感染的疫情,对ARV的研究也日渐深入,Vanderheide等用ARV S1133株生产出了第一代的商品化活疫苗应用于生产[5];2012年起,我国ARV感染疫情逐渐扩大,主要症状也呈现复杂化、多样化。目前,养殖场普遍存在ARV感染,给养禽业带来了不可估量的经济损失,严重阻碍养禽业的发展。研究表明,ARV的细胞亲嗜性较广,能在绿猴肾细胞(Vero)、犬肾细胞(MDCK)、鸡胚成纤维细胞(CEF)、乳地鼠细胞(BHK-21/13)等多种细胞系中增殖,且产生稳定典型细胞病变[6]。丁明洋等[7]用鸭呼肠病毒TH11感染BHK-21细胞后可有效增殖,出现了稳定病变;黄莉等[8]用ARV强、弱毒株分别接种Vero细胞,并分析了接毒后细胞蛋白表达的差异;余爱花等[9]研究了3株鸭源ARV对鸭胚成纤维细胞的致病性,分析了接毒后细胞TNF-α表达水平的差异性。
有关ARV的研究已经较为深入,但研究ARV在DF1细胞上的增殖特性还未见报道,本文就此展开研究。通过将ARV S1133毒株接种DF1细胞,研究其增殖特性,旨在为下一步ARV致病机理的研究和细胞灭活苗的研制奠定技术和理论基础。
1 材料与方法
1.1 材料
DMEM培养基、胎牛血清(FBS)、2.5 g/L胰酶,Gibco公司产品;PBS,Solarbio公司产品;禽呼肠S1133标准强毒株,中国兽医药品监察所提供;DF1细胞,由广西兽医生物技术重点实验室保存提供。
1.2 方法
1.2.1 引物设计及合成 参考NCBI上的ARV S1133株S2基因的核苷酸序列(登录号:KF741763.1),用Primer 5.0软件设计引物,并由华大基因公司合成,引物序列见表1。
1.2.2 ARV S1133接种DF1细胞 将ARV S1133接种至生长良好的DF1细胞上,37℃孵育2h,更换维持液(含20 mL/L FBS的DMEM培养基),在体积分数为5%的CO2中37℃培养,观察CPE。培养3 d后收细胞培养物,置-80℃冻存备用。取上述病毒液再接种细胞,培养3 d后收获细胞培养物,重复接毒3次。置-80℃冻存备用,测定第3代细胞培养物中的病毒滴度。
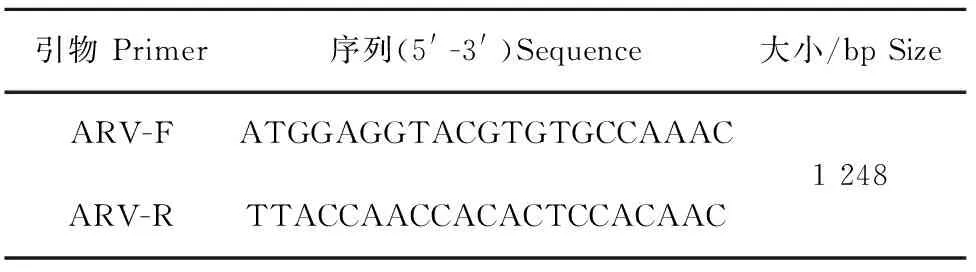
表1 ARV S2基因的核苷酸序列
1.2.3 RT-PCR检测 收获接毒后培养48 h的第3代细胞培养物,提取病毒RNA,用ARV通用引物反转录得到cDNA,将其作为模板用表1中的特异性引物进行PCR扩增,预期扩增片段大小为1 248 bp。PCR程序为94℃ 5 min;94℃ 1 min,55℃ 1 min,72℃ 70 s,循环数为35;72℃ 10 min。PCR产物用10 g/L的琼脂糖凝胶电泳检测。
1.2.4 一步生长曲线的绘制 将上述收获的第3代病毒液接种DF1细胞,分别收取接毒后12、24、36、48、72、96、120 h的病毒液,置-80℃冻存备用。将DF1细胞接种在96孔细胞板上,在体积分数为5%的CO2中37℃过夜培养,观察CPE及记录CPE孔数(重复此过程3次)。按Reed-Meunch法计算7个时间段的半数组织感染量(TCID50)(取3次重复试验的平均值),绘制ARV在DF1细胞上的一步生长曲线。
2 结果
2.1 细胞病变
接毒12 h后即可观察到CPE,在48 h时最明显,主要细胞病变为细胞萎缩、破碎,脱落,成团,边缘不清晰,个别胞体出现变大,死亡等特征,对照组细胞生长良好(图1)。

A.正常;B.接毒后24 h;C.接毒后48 h;D.接毒后72 h
A.Normal;B.Post-infection 24 h;C.Post-infection 48 h;D.Post-infection 72 h
图1 DF1细胞形态(400×)
Fig.1 Morpha of DF1 cells(400×)
2.2 RT-PCR检测结果
收取接种ARV S1133株的第3代DF1细胞培养物,用RT-PCR检测。结果扩增到一条1 248 bp大小片段(图2)。对片段序列与所选的参考株同源性在99.99%以上,说明ARV S1133毒株在DF1细胞上增殖。
2.3 ARV S1133毒株在DF1细胞系上的增殖规律
ARV S1133接种DF1细胞后,分别收取12、24、36、48、72、96、120 h的细胞培养物,测其TCID50, 每个时间点分别做3次重复试验,取平均值。7个时间点的TCID50平均值分别为10-2.49、10-3.5、10-6.84、10-8.9、10-7.86、10-5.38、10-3.64,绘制出生长曲线(图3)。该曲线显示,ARV接种DF1细胞后,0~12 h为静止期,12 h~48 h为快速增长期,在48 h达到最大的病毒效价,TCID50为10-8.9/0.1 mL。
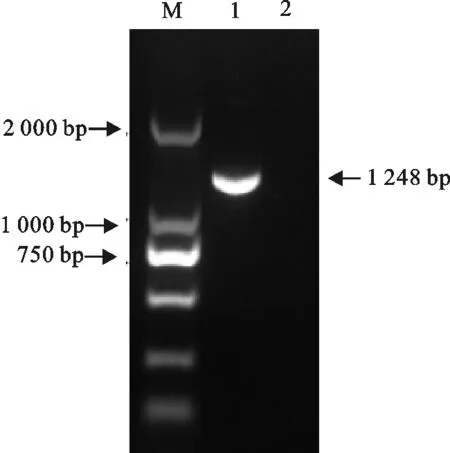
M.DNA 标准DL 2 000;1.PCR扩增产物;2.阴性对照
M.DNA Marker DL 2 000;1.PCR Product;2.Negative control
图2 ARV RT-PCR检测结果
Fig.2 Detection results of ARV by RT-PCR

图3 ARV S1133在DF1细胞上的生长曲线
3 讨论
随着养禽业的快速发展,禽病发生日益增多。近年来,多省份陆续有ARV感染疫情的发生,报道指出ARV有更广的宿主范围,更强的致病性,流行的疫区也日渐扩大,是一种严重危害养禽业健康发展的重要传染病[10]。目前,分离ARV一般采用鸡胚接种,此法虽然单次获毒较多,但需要的时间长,且操作繁琐,在快速获取大量病毒方面不具有时效性,在一定程度上反而限制了ARV的分离[11]。DF1细胞系来源于ELL鸡胚胎,该细胞无禽白血病病毒、肉瘤病毒内源性基因,同时在形态上呈纤维状,是一种稳定的、无肿瘤基因、自发无限增殖的细胞系。目前被广泛用于动物病毒研究、疫苗研制、癌症研究等诸多领域,是生命科学领域重要的病毒转染、培养生物材料。ARV基因组为RNA,而DF1细胞可用于反转录病毒的复制,DF1细胞系有助于ARV对细胞杀伤力的研究。为此,采用了细胞增殖病毒的方法,研究了ARV S1133毒株在DF1细胞上的增殖特性 ,大大缩短了获取大量病毒的时间,大幅提高了获毒效率。
定量描述病毒生长规律的实验曲线称为一步生长曲线,根据病毒的生长特点可划分为静止期、快速增长期、稳定期3个时期[12]。通常用来研究病毒在细胞上的增殖规律的方法主要有测定病毒的TCID50和病毒蚀斑试验[13-14]。本研究直接采取了测定病毒TCID50的方法,并用PCR检测、测序确定细胞仅有ARV感染,无其他病原污染。试验结果显示,ARV接种DF1细胞后,前24 h为静止期,24 h后进入快速增长期,并在48 h达到最大的病毒效价,即TCID50为10-8.9/0.1 mL。这一增殖规律可能与细胞的自身免疫机制有关,至于真正的原因,还待进一步的研究。本研究结果表明,ARV可在DF1细胞上有效地增殖,并伴有典型病变,在接毒48 h后出现病毒效价峰值,TCID50为10-8.9/0.1 mL。ARV感染DF1细胞的最佳收毒时间为接毒后的48 h。本研究结果可为下一步用DF1细胞研制ARV灭活苗及致病机理研究奠定基础[15]。
[1] Saif Y M.禽病学[M].12版.高 福,索 勋,译.北京:中国农业出版社,2012:355-379.
[2] 张昆丽,谢芝勋,黄 莉,等.禽呼肠病毒感染对SPF鸡外周血T细胞亚型变化和细胞因子转录的影响[J].动物医学进展,2015,36(1):1-4.
[3] Javier B,Jose M C.Avian reovirus:structure and biology[J].Virus Res,2006,123(2):105-119.
[4] Jones R C.Avian reovirus infection[J].Revue Scientifique Et Technique,2000,19(2):614-625.
[5] 谢丽基.禽呼肠孤病毒σ3基因的真核表达及σNS基因的原核表达与序列分析[D].广西南宁:广西大学,2006.
[6] Hedayati M,Shojadoost B,Peighambari S M.Detection of avian reoviruses causing tenosynovitis in breeder flocks in Iran by reverse transcription-polymerase chain reaction (RT-PCR) and restriction enzyme fragment length polymorphism (RFLP)[J].Acta Paediatrica,2013,50(4):385-386.
[7] 丁明洋,毕庄莉,朱英奇,等.新型鸭呼肠孤病毒TH11株在BHK21细胞系中的增殖特性[J].中国兽医科学,2015,45(9):918-922.
[8] 黄 莉,谢芝勋,谢丽基,等.禽呼肠孤病毒强毒株和弱毒疫苗株感染Vero细胞的蛋白质组学研究[J].畜牧兽医学报,2016,47(2):331-339.
[9] 余爱花.3株鸭源呼肠孤病毒对鸭胚成纤维细胞致病性研究[D].湖北武汉:华中农业大学,2012.
[10] 陈少莺,陈仕龙,林锋强,等.新型鸭呼肠孤病毒的分离与鉴定[J].病毒学报,2012,28(3):224-230.
[11] Heffels-Redmann U,Muller H,Kaleta E F.Structural and biological characteristics of reoviruses isolated from Muscovy ducks (Cairinamoschata)[J].Avian Pathol,1992,21(3):481-491.
[12] 韩 燕,朱 玲,周远成,等.犬细小病毒四川株的分离鉴定与一步生长曲线的测定[J].中国兽医学报,2014,34(3):379-383.
[13] 代洪波,陈 蕾,朱 玲,等.猪传染性胃肠炎病毒四川株的分离鉴定与一步生长曲线的测定[J].中国兽医科学,2012,42(12):1224-1229.
[14] 付玉志,张洪辉,李传峰,等.Ⅰ型鸭肝炎病毒在鸭胚成纤维细胞系中的增殖特性研究[J].中国预防兽医学报,2013,35(2):110-113.
[15] 孙永珍.传染性喉气管炎病毒LJS09株在LMH细胞上的生长特性研究及糖蛋白gG、gC的抗体制备[D].北京:中国农业科学院,2014.
猜你喜欢
广西中医药大学学报(2021年3期)2021-12-01
养殖与饲料(2019年11期)2019-02-26
生物技术通讯(2019年1期)2019-02-18
天然产物研究与开发(2018年9期)2018-10-08
中国畜禽种业(2018年9期)2018-01-20
广东海洋大学学报(2015年4期)2016-01-13
山东医药(2015年14期)2016-01-12
色谱(2015年6期)2015-12-26
医学研究杂志(2015年4期)2015-06-10
中国医药导报(2015年26期)2015-02-28
