
OP16联合雷帕霉素对人食管鳞癌裸鼠移植瘤生长和细胞凋亡的影响
2018-02-05 06:07李浩男彭柯峥师莹莹刘小林曹燕君郝雨桐覃佳宁侯桂琴
郑州大学学报(医学版) 2018年1期
李浩男,彭柯峥,师莹莹,刘小林,曹燕君,郝雨桐,覃佳宁,侯桂琴
郑州大学药学院临床药学系 郑州 450001
食管鳞癌是我国常见的恶性肿瘤,早期诊断困难、侵袭性强、预后差、易耐药的特点是其病死率居高不下的主要原因[1]。尽管近年来治疗手段不断进步,但是患者的5 a生存率仍然很低,因此寻找新的治疗食管鳞癌的方法及开发新的抗食管鳞癌药物就显得非常急迫。冬凌草作为传统中药,对多种肿瘤尤其是消化道肿瘤具有良好的治疗效果,而且较传统化疗药物具有不良反应少等优点,其化学成分主要包括冬凌草甲素、乙素、丙素等[2]。其中冬凌草甲素是冬凌草主要的抗肿瘤有效成分,通过阻滞细胞周期、诱导细胞凋亡和自噬等作用,对多种肿瘤均有抑制作用[3]。然而,冬凌草甲素水溶性较差,作用机制尚不完全明确,且抗肿瘤活性相对传统化疗药物较差,这些缺点限制了冬凌草甲素的临床应用[4]。OP16是本课题组所在的郑州大学国家药物关键制备技术教育部重点实验室对冬凌草甲素进行结构改造得到的一系列小分子化合物之一,其理化性质更稳定、抗肿瘤活性更强[5]。作者的前期研究[5]已证实OP16在体外能明显抑制食管鳞癌细胞增殖,并能显著抑制食管鳞癌细胞中Akt的磷酸化激活,因而与雷帕霉素在体外联用时具有较强的协同作用。因此本研究拟通过建立食管鳞癌裸鼠移植瘤模型,对OP16与雷帕霉素单独或联合应用的抗肿瘤效果进行体内评价,并对可能的分子机制进行探讨,为OP16作为一种抗肿瘤新药的开发提供实验和理论依据。
1 材料与方法
1.1材料食管鳞癌EC9706细胞购自中科院上海细胞库;RPMI 1640培养基和胎牛血清购自以色列BI生物科技公司;雷帕霉素购自美国Sigma-Aldrich公司;SPF级BALB/c裸鼠(6周龄)购自湖南斯莱克景达实验动物有限公司;PI3K p85α、PI3K p110α、p-Akt(Thr308)、p-Akt(Ser473)、p-mTOR(Ser2448)、mTOR和GAPDH兔抗人单克隆抗体均购自美国CST公司;TUNEL试剂盒购自美国Roche公司。
1.2细胞培养及动物饲养食管鳞癌EC9706细胞在含有体积分数10%胎牛血清的RPMI 1640培养液中,于37 ℃、含体积分数5%CO2的细胞培养箱中进行培养。裸鼠在郑州大学药学院动物中心无菌空气层流室饲养,且保持恒温(26~28 ℃)及恒湿(相对湿度40%~60%)。所有鼠笼、垫料、饲料及饮水全部经过高压蒸汽灭菌处理。动物饲养过程完全符合SPF标准。
1.3成瘤实验40只裸鼠适应性饲养7 d后,取对数生长期的EC9706细胞进行消化、离心、重悬、计数,调整细胞密度至2×107mL-1。裸鼠在无菌室内常规消毒后,于右前肢皮下接种0.2 mL的细胞悬液。接种成功后定期观察小鼠的日常情况包括精神、饮食及排便等,称重并用游标卡尺测定肿瘤的长径(a)和短径(b),按以下公式计算肿瘤体积:V=0.5ab2。该组成瘤率为100%。
1.4裸鼠分组及治疗当移植瘤体积达到60~80 mm3时,采用随机数字表法将裸鼠分为4组,每组10只,分别为:对照组(尾静脉注射生理盐水,隔 d 1次);OP16组(尾静脉注射,20 mg/kg,隔d 1次);雷帕霉素组(腹腔注射,50 μg/kg,隔d 1次);联合治疗组,给药方式及剂量分别同单独治疗组,2种药物每d注射其中1种,轮流注射。给药期间,每d观察裸鼠饮食饮水、精神状态和排便情况等,并用游标卡尺测量肿瘤大小。以生长时间为横坐标,肿瘤体积为纵坐标,绘制肿瘤生长曲线。治疗2周后麻醉处死小鼠,剥取瘤组织,称重并拍照。
1.5各组肿瘤组织中细胞凋亡的TUNEL法检测
将取出的肿瘤组织在40 g/L多聚甲醛中固定后进行石蜡包埋,然后按5 μm的厚度进行切片,使用TUNEL试剂盒对肿瘤组织中细胞凋亡情况进行检测,实验步骤按照试剂盒说明书进行操作。将切片在显微镜下进行观察,正常细胞为蓝色,细胞核内着棕黄色颗粒为TUNEL阳性细胞。每张切片选取5个视野(×400),计数TUNEL阳性细胞数,计算TUNEL阳性率。
1.6各组肿瘤组织中PI3K/Akt/mTOR通路相关蛋白表达量的Westernblot检测用含蛋白酶抑制剂和磷酸酶抑制剂的裂解液裂解肿瘤组织,提取总蛋白。BCA法测定蛋白浓度,上样量均为30 μg。SDS-PAGE法分离蛋白,然后将蛋白转移至硝酸纤维素膜。用含50 g/L脱脂奶粉的T-BST封闭2 h后,分别加入PI3K p85α、PI3K p110α、p-Akt(Thr308)、p-Akt(Ser473)、p-mTOR(Ser2448)、mTOR和GAPDH兔抗人单克隆抗体(按11 000稀释),4 ℃过夜。次日用PBST洗3次,10 min/次,随后加入含二抗的T-BST(按110 000稀释),室温封闭2 h。然后PBST洗3次,10 min/次。ECL超敏发光液孵育1 min后,暗室曝光。实验重复3次,结果用Image J软件进行统计。
1.7统计学处理采用SPSS 19.0进行分析,应用析因设计的方差分析对实验数据进行处理,检验水准α=0.05。
2 结果
2.1 4组荷瘤裸鼠的肿瘤生长情况见图1。在给药期间,4组裸鼠饮食饮水及精神状态良好,体重差异无统计学意义,无死亡出现。虽然治疗期间OP16组、雷帕霉素组肿瘤体积仍在增大,但与对照组相比生长速度相对缓慢,而联合治疗组肿瘤组织则几乎停止生长(P<0.01)。治疗结束后OP16组、雷帕霉素组和联合治疗组的肿瘤质量分别为(0.510±0.100) g、(0.490±0.170) g和(0.170±0.050) g,与对照组(0.900±0.150) g比较均减小;且联合治疗组与OP16组和雷帕霉素组相比,肿瘤质量进一步减小(FOP16=17.562,F雷帕霉素=18.302,F交互=20.716,P<0.001)。

A:各组裸鼠肿瘤生长曲线;B:各组裸鼠处死后剥离的瘤组织图1 OP16联合雷帕霉素对肿瘤生长的影响
2.2 4组肿瘤组织中细胞凋亡情况比较见图2。对照组TUNEL阳性率(5.800±0.520)%明显小于OP16组(56.867±6.300)%、雷帕霉素组(21.733±1.880)%和联合治疗组(71.800±5.320)%,差异有统计学意义(FOP16=209.515,F雷帕霉素=230.235,F交互=445.186,P<0.001);且联合治疗组与OP16组或雷帕霉素组相比,TUNEL阳性率更大(P<0.01)。
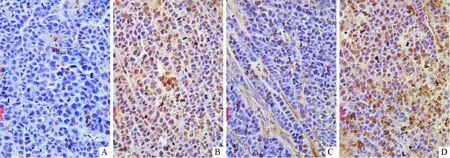
A:对照组;B:OP16组;C:雷帕霉素组;D:联合治疗组图2 OP16联合雷帕霉素对肿瘤组织中细胞凋亡的影响(DAB染色,×400)
2.3治疗后各组肿瘤组织中相关蛋白表达情况见表1、图3。

表1 治疗后各组肿瘤组织中PI3K/Akt/mTOR通路相关蛋白表达量的比较(n=3)
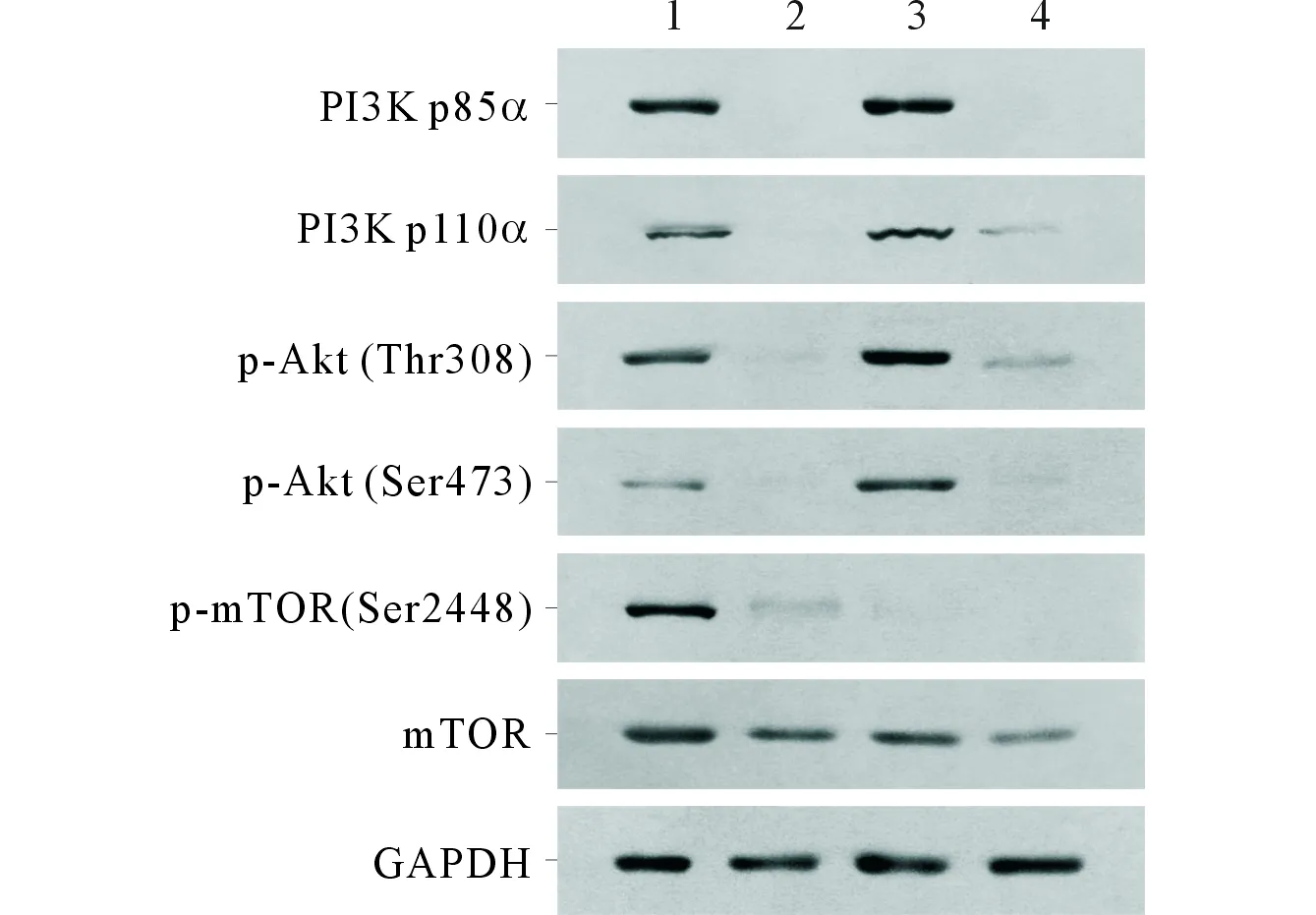
1:对照组;2:OP16组;3:雷帕霉素组;4:联合治疗组图3 OP16联合雷帕霉素对肿瘤 组织中PI3K/Akt/mTOR通路相关蛋白表达的影响
3 讨论
PI3K/Akt/mTOR是一条细胞内重要的信号转导通路,且在多种肿瘤中都处于异常激活状态[6]。其中,PI3K是一种胞内磷脂酰肌醇激酶,可使蛋白激酶Akt第308位上的苏氨酸位点磷酸化而激活Akt,而活化的Akt可以通过磷酸化哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)的第2 448位的丝氨酸位点激活mTOR,从而在细胞周期、细胞凋亡、细胞增殖等方面发挥重要作用[7]。mTOR是一种非典型丝氨酸/苏氨酸蛋白激酶,在多种肿瘤包括食管鳞癌中均为异常激活状态[8]。mTOR由mTORC1和mTORC2两个复合物组成,其中mTORC1主要调控细胞生长和蛋白合成,mTORC2主要调控细胞凋亡[9]。雷帕霉素通过与FKBP-12结合形成复合物从而抑制mTOR通路,因此具有较好的抗肿瘤作用,其衍生物依维莫司等已被用于晚期肾细胞癌、乳腺癌等多种癌症的治疗[10]。作者的前期研究发现,食管鳞癌EC9706细胞中mTOR信号通路处于激活状态[11],并且雷帕霉素可以通过抑制mTOR信号通路从而抑制EC9706细胞的生长[12]。但后续研究发现,这些抑制剂在抑制mTOR从而产生抗肿瘤作用的同时,也能通过一些负反馈通路促进Akt的磷酸化激活[13],Akt的激活对细胞增殖和凋亡有促进作用[14],从而削弱了雷帕霉素及其衍生物的抗肿瘤活性。因此本研究拟建立人食管鳞癌裸鼠移植瘤模型,对OP16和雷帕霉素的联用效果进行体内评价,并对二者联用的分子机制进行进一步探讨。
本研究结果显示,雷帕霉素在单独应用时,确实能使mTOR的表达及磷酸化受到抑制,因此可以抑制裸鼠移植瘤的生长;但与此同时,雷帕霉素也使PI3K的p85α和p110α亚基的表达升高,并促进Akt在Thr308位点及Ser473位点的磷酸化,这也是雷帕霉素单独应用对细胞凋亡的影响较小的原因。然而OP16在单独应用时,能够有效地抑制PI3K的p85α和p110α亚基的表达,同时抑制Akt在Thr308位点及Ser473位点的磷酸化,因此可以抑制肿瘤生长并促进肿瘤组织中细胞凋亡的发生。当二者联用时,OP16可以有效地抑制雷帕霉素引起的PI3K/Akt负反馈激活,且二者联用可以协同抑制Akt下游的mTOR的表达及磷酸化,因此二者联用可以有效地抑制裸鼠移植瘤的生长,并且促进裸鼠移植瘤中细胞凋亡的发生。
综上所述,该研究通过体内实验初步证实了OP16与雷帕霉素联用具有较强协同抗肿瘤活性,并初步探明了二者协同作用的分子机制,为食管鳞癌治疗提供了新的思路,并为OP16作为抗肿瘤注射剂的开发提供实验基础和理论依据。然而OP16的抗肿瘤机制并不完全明确,有待于进一步的临床前和临床试验进行研究。
[1] CHEN W,ZHENG R,BAADE PD,et al.Cancer statistics in China, 2015[J].CA Cancer J Clin,2016,66(2):115
[2] SHI M, LU XJ, ZHANG J, et al.Oridonin, a novel lysine acetyltransferases inhibitor, inhibits proliferation and induces apoptosis in gastric cancer cells through p53- and caspase-3-mediated mechanisms[J].Oncotarget,2016,7(16):22623
[3] XU S,LUO S,YAO H,et al.Probing the anticancer action of oridonin with fluorescent analogues: visualizing subcellular localization to mitochondria[J].J Med Chem,2016,59(10):5022
[4] DING Y,DING C,YE N,et al.Discovery and development of natural product oridonin-inspired anticancer agents[J].Eur J Med Chem,2016,122:102
[5] 杨帅.雷帕霉素和OP16对食管鳞癌细胞增殖及Akt/PTEN表达的影响[D].郑州:郑州大学,2014.
[6] DIENSTMANN R,RODON J,SERRA V,et al.Picking the point of inhibition: a comparative review of PI3K/AKT/mTOR pathway inhibitors[J].Mol Cancer Ther,2014,13(5):1021
[7] PORTA C,PAGLINO C,MOSCA A.Targeting PI3K/Akt/mTOR signaling in cancer[J].Front Oncol,2014,4:64
[8] HOU G,XUE L,LU Z,et al.An activated mTOR/p70S6K signaling pathway in esophageal squamous cell carcinoma cell lines and inhibition of the pathway by rapamycin and siRNA against mTOR[J].Cancer Lett,2007,253(2):236
[9] ALBERT V,HALL MN.mTOR signaling in cellular and organismal energetics[J].Curr Opin Cell Biol,2015,33:55
[10]CHIARINI F,EVANGELISTI C,MCCUBREY JA,et al.Current treatment strategies for inhibiting mTOR in cancer[J].Trends Pharmacol Sci,2015,36(2):124
[11]侯桂琴,范天黎,鲁照明,等.EC9706和Eca109细胞中雷帕霉素靶蛋白信号通路激活状态观察[J].郑州大学学报(医学版),2007,42(2):223
[12]王琼叶,侯桂琴,王莉莉,等.雷帕霉素抑制mTOR信号通路对EC9706细胞生长及凋亡的影响[J].郑州大学学报(医学版),2010,45(3):356
[13]YORI JL,LOZADA KL,SEACHRIST DD,et al.Combined SFK/mTOR inhibition prevents rapamycin-induced feedback activation of AKT and elicits efficient tumor regression[J].Cancer Res,2014,74(17):4762
[14]MUNDI PS,SACHDEV J,MCCOURT C,et al.AKT in cancer: new molecular insights and advances in drug development[J].Br J Clin Pharmacol,2016,82(4):943
猜你喜欢
——雷帕霉素
食品与健康(2022年8期)2022-10-22
波谱学杂志(2022年1期)2022-03-15
国际眼科杂志(2022年3期)2022-03-11
昆明医科大学学报(2022年1期)2022-02-28
云南医药(2021年3期)2021-07-21
实用药物与临床(2021年2期)2021-03-18
昆明医科大学学报(2020年11期)2020-12-28
中老年保健(2020年2期)2020-12-04
分析化学(2017年12期)2017-12-25
安徽医科大学学报(2015年9期)2015-12-16
