
天山樱桃种质资源表型多样性研究
2018-01-31 03:09李春侨董凯向刘春燕
西北农业学报 2018年1期
李春侨,周 龙,陆 彪,董凯向,刘春燕
(1.新疆农业大学 林学与园艺学院,乌鲁木齐 830052;2.特克斯县林业局,新疆特克斯 835500)
天山樱桃(CerasustianschanicaPojark)又称野樱桃,属于蔷薇科(Rosaceae)樱桃属(Cerasus)矮生樱亚属植物,是中国濒临灭绝的树种之一,也是野生珍奇果品资源,在国内仅分布于新疆伊犁和塔城地区[1]。天山樱桃集中分布在海拔1 100~1 500 m的阳坡地,冬季最低温达-35 ℃,平均年降水量300~600 mm,土壤类型多为栗钙土,与栽培樱桃相比有较强的抗寒、抗旱和耐瘠薄能力,且矮化效果明显,因此了解天山樱桃遗传多样性对栽培樱桃的育种和优良砧木的筛选具有重要意义。目前,对于天山樱桃的研究主要集中在繁殖生物学特性[2]、花粉萌发特性[3]和花芽形态分化[3]等方面,尚未涉及到天山樱桃表型多样性方面的研究。刘胤等[4]对80份中国樱桃种质资源的26个表型性状进行多样性分析,认为参试材料的数量性状在群体间和群体内均表现出极显著差异。曾斌等[5]研究新疆野巴旦杏天然居群的表型多样性时发现叶片形态在群系间和群系内存在广泛差异,且形态变异主要来源于群系内。周龙等[6]利用表型性状变异研究新疆野生樱桃李遗传多样性,认为枝条长度、枝条粗度、叶片长度和叶柄长度是造成新疆野生樱桃李表型差异的主要因素。表型多样性是遗传多样性与生境多样性的综合体现,主要研究各居群在不同生境条件下的表型变异。有关表型多样性方面的研究文献较多,并且随着遗传多样性研究的加强而逐步成为热点。开展天山樱桃野生群体表型多样性研究有利于该物种种质资源的收集、保存、评价与利用研究。为此,本研究通过对天山樱桃4个野生居群(特克斯、喀拉峻、大西沟、裕民县)的16个表型性状进行观测,分析不同居群间形态性状的多样性和变异特点,以期为天山樱桃种质资源保护与利用提供参考。
1 材料与方法
1.1 天山樱桃分布区域概况
裕民县野生居群位于巴尔鲁克山以北的山地草原,海拔1 115 m,为温带大陆性气候,年平均降水量280 mm,受岩画风景区与野扁桃风景区的影响,人为干扰频繁;特克斯县野生居群位于蒙古乡北山较为封闭狭长的山谷中,海拔1 375 m,高山气候,年平均降水量383 mm,由于实行禁牧措施,人为干扰较少;喀拉峻野生居群位于天山北麓,海拔1 454 m,是典型的山地草甸类型草原,年平均降水量600 mm,放牧严重;大西沟野生居群位于天山西部,海拔1 165 m,为山地逆温带气候,年平均降水量671 mm,受中华福寿山景区影响,人为干扰频繁。4个天山樱桃野生居群的地理位置如表1所示。

表1 4个天山樱桃野生居群的地理位置Table 1 Geographical position of 4 natural populations of Cerasus tianschanica Pojark
1.2 材 料
从2012年起,连续5 a对天山樱桃分布区(特克斯、喀拉峻、大西沟、裕民县)进行野外实地调查,按照不同海拔高度,不同坡向,选取不同的植株作为定点观测样株。为尽可能降低树龄对表型特征的影响,选择基径相近的单株,共筛选45棵具有代表性的样株,为避免株间亲缘关系较近,每样株相隔不少于500 m。采用全球卫星定位系统(Global Positioning System,GPS)定位,并挂牌标记以便跟踪调查。
1.3 方 法
1.3.1 表型性状观测 枝条停止生长后,使用电子游标卡尺(测量精度为0.01 mm)测量1 a生枝长度和粗度;取新梢中部30枚成龄叶片,测定叶片长度和宽度;盛花期,随机选取30朵花,测量花冠直径、花瓣长、花瓣宽、花筒长度、雄蕊数目和雌蕊长度;果实成熟期,采集成熟度一致的果实30个,测量果实纵径和横径,并用1/1 000电子天平称量果实,剥离果皮及果肉,使用手持折光仪检测果肉可溶性固形物质量分数,并称量种核质量,剥离种壳,称量种仁质量。
1.3.2 数据处理 变异系数是衡量材料中各观测指标变异程度的统计量,当观测指标单位不一致时,可使用变异系数消除单位对多个材料变异程度的影响。Simpson多样性指数与Shannon-weaver多样性指数是用来估算遗传多样性水平高低的指标。欧氏距离是居群间遗传关系远近的指标。类平均法是一种常见的聚类分析方法,目的是寻找数据中潜在的自然分组结构,此方法较适用于物种树的建立。采用DPS 7.05数据处理软件计算各野生居群表型性状的变异系数、Simpson多样性指数、Shannon-weaver多样性指数和欧氏距离(Euclidean distance),采用类平均法(UPGMA)对居群进行聚类分析。利用IBM SPSS Statistics 19.0软件对天山樱桃16个表型性状进行主成分分析,确定形成表型差异的主要表型性状。
2 结果与分析
2.1 天山樱桃野生居群表型多样性
变异系数、Shannon-weaver指数和Simpson指数是衡量种质资源遗传多样性的重要指标,值越高,遗传多样性越丰富。如表2所示,16个表型性状的变异系数为3.08%~50.32%,Shannon-weaver指数为2.22~4.00,Simpson指数为0.78~1.47,4个野生居群的平均变异系数、Shannon-weaver指数和Simpson指数由大到小排列一致,均为裕民县>大西沟>特克斯>喀拉峻,表明裕民县野生居群的表型多样性最为丰富,喀拉峻野生居群表型多样性水平最低。
2.2 天山樱桃表型性状的变异及分化
变异系数表示群体的离散程度,表型变异系数越大,表型性状的变异程度越大。由表3可以看出,天山樱桃16个表型的变异系数为8.67%~48.78%,其中种核质量(40.61%)、单果质量(48.78%)和果实纵径(40.25%)的变异系数较大,可溶性固形物质量分数(8.67%)、雄蕊数目(11.24%)和花瓣宽(12.81%)的变异系数较小,稳定性好。16个表型性状的分化系数变幅为3.74%~51.93%,种核质量(51.93%)的表型分化系数最大,说明天山樱桃种核质量受环境影响较大,各表型性状平均分化系数为25.77%,即居群内表型分化系数为74.23%,表明天山樱桃野生居群内多样性程度高于野生居群间。

表2 天山樱桃野生居群变异系数、Shannon-weaver指数和Simpson指数Table 2 Variable coefficient,Shannon-weaver index and Simpson index of different populations of Cerasus tianschanica Pojark

表3 天山樱桃表型性状变异系数及分化系数Table 3 Variable coefficient and differentiation coefficient from morphological data of Cerasus tianschanica Pojark
2.3 天山樱桃野生居群的聚类分析
4个天山樱桃野生居群间的欧式距离为3.75~7.16,欧氏距离平均值为5.56,表明4个天山樱桃野生居群的亲缘相对较近。由欧式距离矩阵可知特克斯野生居群与喀拉峻野生居群的欧式距离最小(3.75),遗传分化程度小;裕民县野生居群与大西沟野生居群的欧式距离最大(7.16),遗传分化程度大,亲缘关系相对较远(表4)。
根据4个野生居群间的欧氏距离,采用UPGMA聚类法对居群进行聚类分析(图1)。4个天山樱桃野生居群的距离系数为0~6.34,在距离系数为5.27时,4个野生居群被分为3大类;裕民县野生居群单独为第1类,特克斯野生居群与喀拉峻野生居群聚为第2类,大西沟野生居群为第3类。
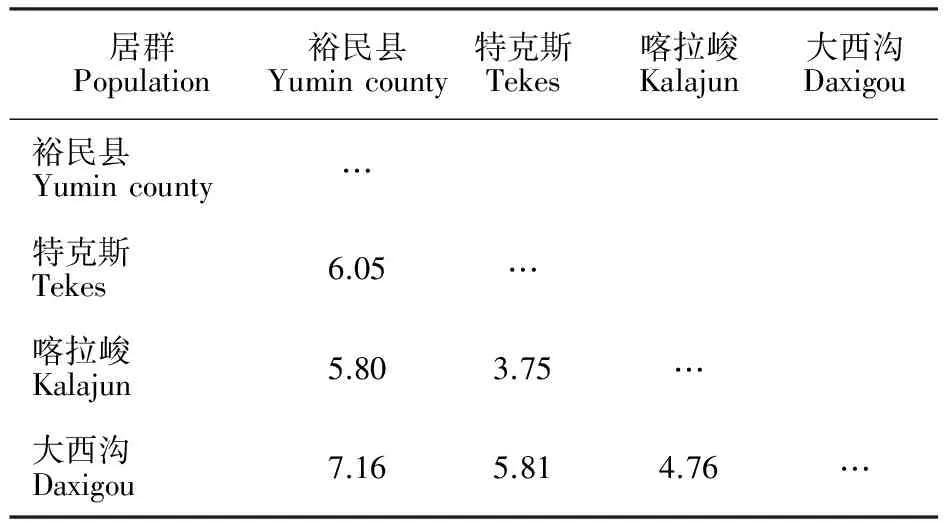
表4 不同天山樱桃野生居群间表型性状的欧式距离Table 4 Euclidean distances from morphological data of different populations of Cerasus tianschanica Pojark

图1 基于16个表型性状的UPGMA聚类图Fig.1 UPGMA cluster of 16 phenotypic dates
2.4 天山樱桃表型性状主成分分析
主成分分析是将原来众多具有一定相关性的变量,在不损失原有数据信息的基础上,重新组合成彼此相互独立的变量[7]。通过对天山樱桃16个表型性状的主成分分析(表5、表6),前4个主成分的特征根大于1,累计贡献率达76.02%。其中第1主成分的贡献率为35.07%,起决定性作用的表型性状有果实纵径(0.40)、果实横径(0.39)、单果质量(0.39)、种核质量(0.39)和种仁质量(0.38)。第2主成分贡献率为20.15%,起决定性作用的表型性状有叶片长度(0.38)、叶片宽度(0.43)、1 a生枝长度(0.31)和雌蕊长度(0.45)。第3主成分贡献率为12.43%,起决定性作用的表型性状有花冠直径(0.53)、花瓣长(0.54)、花筒长度(0.45)和雄蕊数目(0.39)。第4主成分的贡献率为8.37%,叶片长度(0.34)、1 a 生枝粗度(0.51)、花瓣宽(-0.32)和可溶性固形物质量分数(-0.49)起决定性作用。使用4个主成分的得分进行居群聚类(图2),与使用16个表型性状所得聚类结果基本一致,表明筛选出的表型性状可以代表天山樱桃16个表型性状的数据信息。
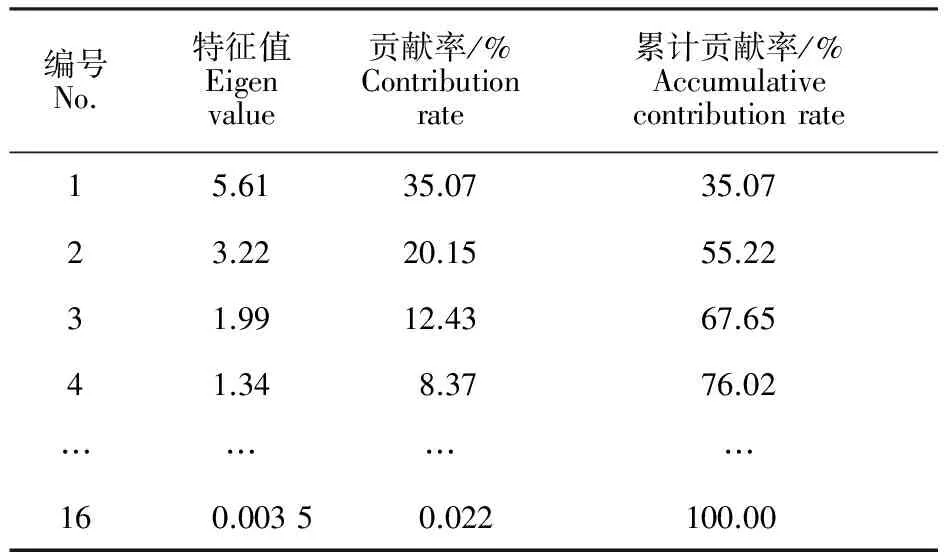
表5 主成分特征值、贡献率及累计贡献率分析Table 5 Analysis on matrix eigen value,contribution rateand accumulative contribution rate of principal component

表6 表型性状主成分的负载量分析Table 6 Analysis of load capacity of principal component of phenotypic traits
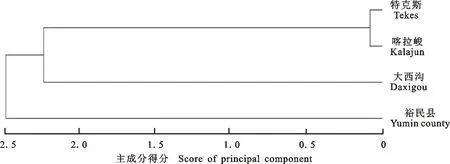
图2 基于主成分得分的系统聚类Fig.2 System cluster based on score of principal component
3 讨 论
3.1 天山樱桃表型性状变异特征
植物表型性状既具有变异性,又有其稳定性,同时受植物自身遗传物质和生态环境两方面的影响[8]。本研究中16个表型性状的变异幅度为8.67%~48.78%,其中1 a生枝长、果实纵径、单果质量和种核质量的变异系数较大。各器官表型性状变异系数表现为果实>种子>枝叶>花,这可能与天山樱桃作为野生果树受坡向、土壤特征和水分等微气候的影响有关,果实性状表现出较为丰富的变异。梁玉琴等[9]分析河南省柿的表型多样性,刘娟等[10]研究新疆杏表型多样性时也发现同样的结果,均认为表型性状的各个指标中果实性状变异最为丰富,易受生境影响。
3.2 天山樱桃表型分化
群体表型分化系数是群体间表型方差占群体间和群体内表型总方差的比例[11]。本研究的16个表型性状表型分化系数均值为25.77%,居群内变异显著高于居群间变异,天山樱桃的变异主要来源于居群内。在中国板栗[12]、海南岛青梅[13]和浙江柿[14]的表型多样性研究中也发现变异主要来源于居群内,与本研究结果一致。天山樱桃属于异花授粉植物,由于长期的授粉杂交,在同一居群内后代表现出丰富的多样性,其次,野生居群所处地形、地貌与气候类型多样,也会促使天山樱桃形成丰富的表型变异。
3.3 天山樱桃表型性状主成分分析
本研究对天山樱桃的16个表型性状进行主成分分析,结果显示:前4个主成分因子的累计贡献率达到76.02%,认为果实纵径、果实横径、雌蕊长度、花瓣长和1 a生枝长度等是评价天山樱桃表型多样性的重要指标。刁松锋等[15]对无患子表型性状进行主成分分析,认为种子质量、种子纵径、种子横径、果实纵径和果实横径可以代表原始因素的大部分信息。张强英等[16]将榆叶梅野生群体的16表型性状进行主成分分析,认为花朵大小、花朵颜色、花瓣萼片反卷程度和花密度可以作为今后榆叶梅多样性评估的指标。与前人研究有一定的相似之处,但也存在差异,这可能与材料选择和测定指标不同有关。综合分析认为,本研究所筛选的果实纵径、果实横径、雌蕊长度、花瓣长和1 a生枝长度可以代表原始数据大部分信息进行天山樱桃表型多样性的评价。
4 结 论
综上所述,4个天山樱桃野生居群中,裕民县野生居群的表型多样性最为丰富,大西沟野生居群次之,喀拉峻野生居群的表型多样性水平最低。观测的枝、叶、花和果表型性状中以果实性状变异最为丰富,花性状变异最小,遗传稳定性相对较好。天山樱桃表型分化系数均值为25.77%,表明天山樱桃的变异主要来源于居群内。聚类分析表明特克斯野生居群与喀拉峻野生居群亲缘关系最近,裕民县野生居群与大西沟野生居群亲缘关系最远。16个表型性状中果实纵径、果实横径、雌蕊长、1 a生枝长度和花冠直径是天山樱桃进行表型观测时的重要指标。
Reference:
[1] 林培钧,崔乃然.天山野果林资源-伊犁野果林综合研究[M].北京:中国林业出版社,2000.
LIN P J,CUI N R.The Wild Fruit Forest Resources[M].Beijing: Chinese Forestry Publishing House,2000.
[2] 周 龙,彭 妮,王 超,等.不同激素处理对天山樱桃绿枝扦插生根效果研究[J].新疆农业科学,2013,51(12):2236-2240.
ZHOU L,PENG N,WANG CH,etal.Study on different hormone treatments on soft wood cutting rooting efficiency ofCerasustianschanicaPojark[J].XinjiangAgriculturalSciences,2013,51(12):2236-2240.
[3] 王 超,周 龙,彭 妮,等.天山樱桃花粉特性研究[J].新疆农业大学学报,2013,36(6):449-453.
WANG CH,ZHOU L,PENG N,etal.A study on the pollen characterisics ofCerasustianschanicaPojark[J].JournalofXinjiangAgriculturalUniversity,2013,36(6):449-453.
[4] 刘 胤,陈 涛,张 静,等.中国樱桃地方种质资源表型性状遗传多样性分析[J].园艺学报,2016,43(11):2119-2132.
LIU Y,CHEN T,ZHANG J,etal.Genetic diversity analysis of chinese cherry Landraces (Prunuspseudocerasus) based on phenotypic traits[J].ActaHorticulturaeSinica,2016,43(11):2119-2132.
[5] 曾 斌,罗淑萍,李 疆,等.新疆野巴旦杏天然居群叶片性状表型多样性研究[J].新疆农业科学,2008,45(2):221-224.
ZENG B,LUO SH P,LI J,etal.Study on phenotypic diversity of morphologic characteristics ofAmygdalusledebourianashlecht leafs in Xinjiang[J].XinjiangAgriculturalSciences,2008,45(2):221-224.
[6] 周 龙,胡建芳,许 正,等.野生樱桃李天然群体表型多样性研究[J].新疆农业大学学报,2011,34(3):222-225.
ZHOU L,HU J F,XU ZH,etal.A study on phenotypic diversity of natural populations ofPrunusdivaricataLdb[J].JournalofXinjiangAgriculturalUniversity,2011,34(3):222-225.
[7] 唐启义.DPS数据处理系统—实验设计、统计分析及数据挖掘[M].北京:科学出版社,2010.
TANG Q Y.DPS Data Processing System:Experimental Design,Statistical Analysis and Data Mining[M].Beijing:Science Press,2010.
[8] 肖 亮,蒋建雄,易自力,等.广西省芒野生居群表型多样性研究[J].草业学报,2013,22(4):43-50.
XIAO L,JIANG J X,YI Z L,etal.A study on phenotypic diversity ofMiscanthussinensisnatural population in Guangxi province[J].ActaPrataculturaeSinica,2013,22(4):43-50.
[9] 梁玉琴,韩卫娟,张嘉嘉,等.河南省柿种质资源表型多样性研究[J].中国农业大学学报,2015,20(1):74-85.
LIANG Y Q,HAN W J,ZHANG J J,etal.Phenotypic diversity of persimmon germplasms in Henan province[J].JournalofChinaAgriculturalUniversity,2015,20(1):74-85.
[10] 刘 娟,廖 康,曼苏尔·那斯尔,等.新疆杏种质资源表型多样性研究[J].果树学报,2014,31(6):1047-1056.
LIU J,LIAO K,Mansuer Nasier,etal.Research on phenotypic diversity of apricot germplasm resources in Xinjiang[J].JournalofFruitScience,2014,31(6):1047-1056.
[11] 郭 宁,杨树华,葛维亚,等.新疆天山山脉地区疏花蔷薇天然居群表型多样性分析[J].园艺学报,2011,38(3):495-502.
GUO N,YANG SH H,GE W Y,etal.Phenotypic diversity of natural populations ofRosalaxaRetz in Tianshan Mountains of Xinjiang[J].ActaHorticulturaeSinica,2011,38(3):495-502.
[12] 江锡兵,龚榜初,刘庆忠,等.中国板栗地方品种重要农艺性状的表型多样性[J].园艺学报,2014,41(4):641-652.
JIANG X B,GONG B CH,LIU Q ZH,etal.Phenotypic diversity of important agronomic traits of local cultivars of Chinese chestnut[J].JournalofFruitScience,2014,41(4):641-652.
[13] 尚帅斌,郭俊杰,王春胜,等.海南岛青梅天然居群表型变异[J].林业科学,2015,51(2):154-162.
SHANG SH B,GUO J J,WANG CH SH,etal.Phenotypic variations in natural populations ofVaticamangachapoiinHainan[J].ScientiaSilvaeSinicae,2015,51(2):154-162.
[14] 井振华,李 皓,邵文豪,等.浙江柿天然群体表型多样性研究[J].植物研究,2010,30(3):325-331.
JING ZH H,LI H,SHAO W H,etal.Phenotypic diversity of natural populations inDiospyrosglaucifolia[J].BulletinofBotanicalResearch,2010,30(3):325-331.
[15] 刁松锋,邵文豪,姜景民,等.基于种实性状的无患子天然群体表型多样性研究[J].生态学报,2014,34(6):1451-1460.
DIAO S F,SHAO W H,JIANG J M,etal.Phenotypic diversity in natural populations ofSapindusmukorossibased on fruit and seed traits[J].ActaEcologicaSinica,2014,34(6):1451-1460.
[16] 张强英,张启翔,程堂仁.榆叶梅野生群体表型多样性的研究[J].中南林业科技大学学报,2012,32(5):155-160.
ZHANG Q Y,ZHANG Q X,CHENG T R.Study on phenotypic diversity ofPrunustrilobawild populations[J].JournalofCentralSouthUniversityofForestry&Technology,2012,32(5):155-160.
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
纺织学报(2022年8期)2022-08-26
黄河之声(2021年12期)2021-10-25
浙江中医药大学学报(2021年6期)2021-07-12
天津医科大学学报(2021年1期)2021-01-26
中国粮油学报(2020年12期)2021-01-09
意林·全彩Color(2019年11期)2019-12-30
中国男科学杂志(2016年5期)2016-12-01
中国康复理论与实践(2015年10期)2015-12-24
作物研究(2014年6期)2014-03-01
