
电针神庭、百会对脑缺血再灌注大鼠认知功能及Beclin-1表达的影响①
2018-01-31 02:34冯晓东史景湾明月刘承梅
中国康复理论与实践 2018年1期
冯晓东,史景,湾明月,刘承梅
河南中医药大学第一附属医院康复中心,河南郑州市450000
认知障碍在脑卒中患者中发病率达56.6%[1],严重影响脑卒中患者的整体功能恢复和身心健康[2]。针刺百会、神庭穴可以改善脑卒中患者的学习记忆能力[3],其具体作用机制并未完全阐明。
自噬(autophagy)是真核细胞通过代谢和分解其自身蛋白质或衰老细胞器,并进一步消化和再利用的生物过程[4]。在缺血缺氧等应激状态下,神经细胞死亡率高,自噬激活明显提高细胞存活率,认为应激状态下自噬对神经细胞产生保护作用[5-7]。Beclin-1作为自噬标记物,是衡量自噬发生程度的方法之一[8]。本研究制备大脑中动脉闭塞(middle cerebral artery occlusion,MCAO)脑缺血再灌注大鼠模型,观察电针神庭、百会穴对MCAO大鼠学习记忆能力、脑梗死体积以及Beclin-1表达的影响。
1 材料与方法
1.1 实验动物
健康雄性清洁级Sprague-Dawley大鼠45只,体质量(260±20)g,由郑州大学实验动物中心提供,合格证号41003100004006。饲养在河南省中医院中心实验室的独立通风笼具中,每笼5只,予自由饮食、饮水,适应性饲养1周。实验过程严格按照国际动物保护和使用指南的规定进行。
1.2 实验试剂和仪器
水合氯醛(30037516):镇江德为化学品有限公司。2%2,3,5-氯化三苯基四氮唑(2,3,5-triphenyltetrazolium chloride,TTC)染色液(G3005-100)、BCA蛋白浓度测定试剂盒(PC0020-500)、10×丽春红染液(P0012-10):北京索莱宝科技有限公司。FITC山羊抗兔IgG(h+1,SA00003-2):武汉三鹰生物技术有限公司。GAPDH抗体(GTX100118-S)、Beclin-1抗体(GTX55535):美国GENETEX公司。
Morris水迷宫(XR-XM101):上海欣软信息科技有限公司。G6805电针仪:上海华谊医用仪器有限公司。华佗牌30号0.5寸毫针:苏州医疗用品厂。
1.3 分组和造模
大鼠以随机数字表法分成假手术组、模型组、电针组,各15只。术前动物禁食24 h。称重后,腹腔注射10%水合氯醛3 ml/kg麻醉,参造Longa法[9]建立左侧MCAO模型。分离并暴露左颈总动脉、颈外动脉和颈内动脉,依次结扎颈总动脉近心端、颈外动脉远心端,并于颈总动脉分叉处近心端预留结扎线。微动脉夹夹闭远端颈内动脉,距颈总动脉分叉处1 mm处用显微剪刀剪一切口,将已备好的线栓插入颈内动脉,插入长度约18~22 mm,至大脑前动脉近端,完全阻断大脑中动脉血供。缚紧预先放置于颈总动脉的结扎线。常规缝合消毒,4-0缝合线缝合伤口。缝合时皮肤外留线栓1 cm。2 h后,轻柔回抽线栓至颈总动脉分叉处。
术中保持室温25℃左右,白炽灯照射动物以保持肛温[9]。实验过程中密切观察大鼠变化,注意动物保温。
假手术组只分离血管,不结扎和插入栓线。
动物完全苏醒后,Longa法行神经行为学评分。0分,无神经缺失的症状;1分,不能完全伸展对侧前爪;2分,向外(右)侧转圈;3分,向对侧倾倒;4分,神经缺失严重,意识丧失,不能自发行走[9]。0分、4分大鼠予以排除。合格大鼠放回笼中正常饲养。
1.4 方法
术后2 h,电针组参考《实验针灸学》取大鼠神庭和百会穴,用华佗牌30号0.5寸毫针,神庭穴向上斜刺,深2 mm;百会穴向前斜刺,深2 mm。接G6805-2A电针仪,电压峰值6 V,以鼠耳轻轻抖动为度,疏密波,频率1~20 Hz。每次30 min,每天1次,共7 d[10]。假手术组和模型组造模后回笼饲养,予同等条件抓取。
1.5 观察指标
1.5.1 神经行为学评分
术后每天同一时间采用Longa评分进行评定,记录术后1 d、3 d、5 d、7 d的评分进行比较。
1.5.2 Morris水迷宫检测
术后4 d起连续训练4 d。水迷宫直径160 cm,深55 cm,水深30 cm,水温22~26℃;池壁上4个等距离点分水池为4个象限,任选一象限中央放置平台,平台直径12 cm,高28 cm,没于水面下2 cm。池壁外标4个入水点,水池周围参照物保持不变。
1.5.2.1 定位航行实验
连续训练4 d,每天训练4次,分别从4个象限入水点将大鼠放入水中。如果大鼠在90 s内爬上平台,并停留3 s以上,则认为大鼠找到平台,此为逃避潜伏期。如果90 s内大鼠未能找到平台,则拖拽其尾部,将其引导到平台,熟悉10 s,潜伏期计为90 s。每次训练间隔5 min。取4个方向潜伏期的平均数作为该日成绩。
1.5.2.2 空间探索实验
测试第5天上午,撤去平台,取任意入水点将大鼠放入水中,观察并记录90 s内大鼠穿过原平台区域的次数[10]。
1.5.3 TTC染色
治疗7 d结束后,各组取5只动物断头处死,生理盐水灌注,取脑,脑组织迅速置于-20℃冰箱冷冻20 min,至脑组织变硬。从大脑半球额极到枕极做连续冠状位切片,厚约2 mm,共6片。脑片放入2%TTC溶液中,37℃恒温箱15~30 min,并不时翻动脑片,使脑片均匀接触染色液。整个过程注意避光。Image-Pro Plus 6.0软件测量脑梗死面积和大脑面积,计算脑梗死体积百分比

1.5.4 Western blotting
治疗7 d结束后,各组取5只动物断头处死取脑。取大鼠大脑海马组织,置液氮罐中速冻,再置-80℃冰箱保存。取大鼠脑组织200 mg,加裂解缓冲液2 ml,15,000 r/min 4℃离心15 min后取上清。BCA法测蛋白浓度,用凝胶加样缓冲液将各管蛋白浓度调为一致。取样品20 ml煮沸5 min,120 V、50 mA电泳1.5 h;将凝胶取出,80 V、100 mA转印2 h至PVDF膜。取出,丽春红预染,证实蛋白质确实转移到膜上。5%脱脂奶粉封闭2 h,TBS洗2次,每次5 min。分别加入兔Beclin-1单克隆抗体和内参抗体GAPDH孵育(均1∶4000),4℃过夜。TBST洗2次,每次5 min。加入碱性磷酸酶标记山羊抗兔IgG(1∶3000),室温2 min。TBST洗2次,每次5 min。将滤膜放入配好的显色液,ECL法发光,于化学发光成像系统中成像,Image Lab软件检测条带灰度值,取Beclin-1与GAPDH相对灰度进行半定量分析。
1.6 统计学分析
应用SPSS 18.0统计软件进行数据处理。计量资料以(xˉ±s)表示。Longa评分不符合正态分布,在模型组与电针组间进行秩和检验。其他多组计量资料符合正态分布,进行单因素方差分析(One-way ANOVA),两两比较,若方差齐者用LSD法,方差不齐则用Games-Howell法。显著性水平α=0.05。
2 结果
2.1 神经行为学评分
造模大鼠30只全部成功。造模后1 d模型组死亡1只。与模型组比较,电针组Longa评分术后1 d无显著性差异,3 d、5 d、7 d低于模型组(P<0.05)。见表1。

表1 各组Longa评分比较
2.2 Morris水迷宫检测
定向航行实验结果表明,随着时间推移,各组逃避潜伏期逐渐缩短,电针组少于模型组(P<0.05)。空间探索实验,电针组穿越平台次数多于模型组(P<0.05)。见表2、表3。
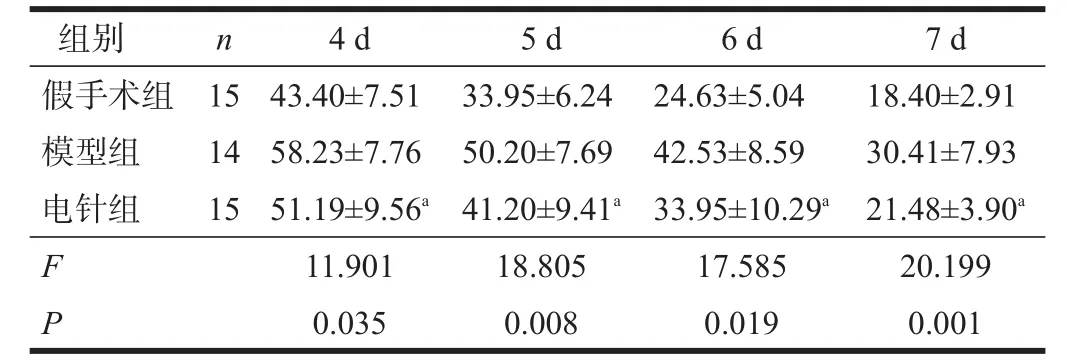
表2 各组逃避潜伏期比较(s)
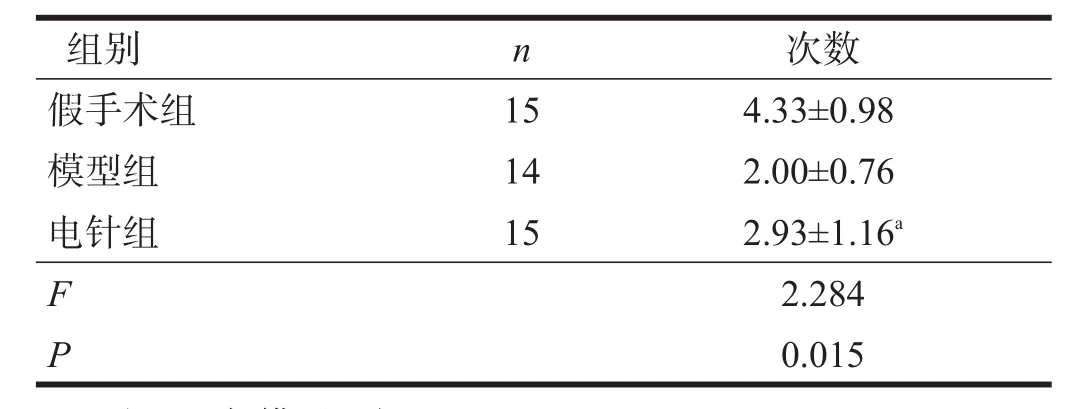
表3 各组穿越平台次数比较
2.3 脑梗死体积
与模型组比较,电针组脑梗死体积占全脑百分比(6.59±0.94)%,显著小于模型组(28.04±4.89)%(F=7.651,P<0.001)。
2.4 Western blotting
与假手术组比较,模型组Beclin-1表达增加(P<0.05),电针组Beclin-1表达比模型组增加(P<0.05)。见表4。

表4 各组Beclin-1表达(%)
3 讨论
本研究显示,电针可改善MCAO大鼠学习记忆能力,改善脑缺血再灌注后病理损伤,同时伴有Beclin-1表达升高。
Beclin-1位于17q21,有12个外显子,编码蛋白相对分子质量60,000,由450个氨基酸序列构成[11]。Beclin-1也称BECN1,是酵母ATG6的同系物,是自噬体形成过程中必需分子,在自噬过程中Beclin-1的表达上升[12-13]。作为自噬的直接执行者,Beclin-1蛋白除了接受自噬信号,还可接受其他信号,对自噬进行调节。Beclin-1已经成为衡量自噬发生程度的检测方法之一[14]。
自噬是细胞受刺激后,通过溶酶体途径降解细胞内物质的统称,是细胞自我消化的过程。自噬也是近年来逐渐被认识的细胞除坏死和凋亡外第3种死亡方式[15-16]。作为一种程序化的细胞内降解机制,自噬的生理和病理作用非常广泛,涉及癌症、神经退行性病变、代谢性疾病、衰老以及免疫系统疾病[17-19]。Beclin-1能介导自噬蛋白定位于吞噬泡,调控哺乳动物自噬体的形成与成熟[20]。在生理条件下,自噬处于低水平,主要清除细胞内被降解的蛋白;在缺血再灌注等因素刺激下,细胞启动自噬机制清除损伤线粒体,同时提高细胞对低氧的耐受力,对细胞起一定保护作用[21]。脑缺血后,大量活性氧、自由基释放引起氧化应激,上调微管相关蛋白1轻链3、Beclin-1表达,从而诱导细胞自噬[22]。较早的研究显示,脑缺血缺氧后,自噬可降解损伤的细胞器以避免凋亡,继而产生能量以避免离子失衡和细胞坏死[8]。但之后的一些研究表明,自噬在缺血性脑损伤过程中会产生复杂的影响,甚至产生负面作用,即自噬在缺血性脑损伤中有可能发挥双刃剑作用[23]。在轻度饥饿、缺氧等情况下,自噬不仅通过降解蛋白提供能量,且通过降解损伤蛋白合成新的蛋白,从而保护机体细胞[24]。若重度饥饿、缺氧,或延长细胞损伤时间,激活凋亡相关调控蛋白产生凋亡后,自噬也过度激活;过度激活的自噬又进一步促进凋亡发生,从而对机体产生损伤作用[25]。
中医认为,督脉是人体诸阳之汇,有“总督诸阳”和“阳脉之海”之说。“病变在脑,首取督脉”,为治疗脑部疾病首选。神庭、百会是督脉穴位,与脑密切联系,是调节大脑功能的要穴。电针神庭、百会改善脑卒中后认知功能的作用在前期临床及基础实验中均已证实[3,10,26]。在研究显示,电针上调Beclin-1表达,从而诱发自噬,伴随认知功能和病理损伤的改善,提示Beclin-1得到适度表达,起到神经保护作用。
综上所述,电针神庭、百会穴可治疗MACO大鼠学习记忆障碍,这种治疗作用可能与上调自噬相关基因Beclin-1从而启动自噬系统网络有关。要精确调控自噬,为临床治疗提供新的治疗靶点,需要更清楚地了解自噬调控通路的每一个环节。这仍有待进一步深入。本研究只检测了与自噬相关的一种基因,存在一定局限性,有待今后研究继续改善。
[1]曲艳吉,卓琳,詹思延.中国脑卒中后认知障碍流行病学特征的系统评价[J].中华老年心脑血管病杂志,2013,14(12):1294-1301.
[2]Park JH,Kim BJ,Bae H,et al.Impact of post-stroke cognitive impairment with no dementia on health-related quality of life[J].JStroke,2013,15(1):49-56.
[3]冯晓东,刘娇,陈立典.电针对局灶性脑缺血大鼠学习记忆能力的影响及其机制[J].中国康复理论与实践,2013,19(3):227-230.
[4]Marcucci F,Bellone M,Caserta CA,et al.Pushing tumor cells towards a malignant phenotype:Stimulifrom the microenvironment,intercellular communications and alternative roads[J].Int JCancer,2014,135(6):1265-1276.
[5]Wang P,Xue Y,Wei K,et al.ARRB1/β-arrestin-1 mediates neuroprotection through coordination of BECN1-dependent autophagy in cerebral ischemia[J].Autophagy,2014,10(9):1535-1548.
[6]王燕梅.自噬参与低氧后处理对成年大鼠短暂全脑缺血的保护作用[D].广州:广州医学院,2012.
[7]丁培炎,王冬,赫曼,等.短期禁食诱导自噬对大鼠脑缺血损伤的保护作用[J].中华神经外科疾病研究杂志,2013,12(3):201-204.
[8]Gabryel B,Kost A,Kasprowska D.Neuronal autophagy in cerebral ischemia-a potential target for neuroprotective strategies?[J].Pharmacol Rep,2012,64(1):1-15.
[9]Longa EZ,Weinstein PR,Carlson S,et al.Reversible middle cerebral artery occlusion without craniectomy in rats[J].Stroke,1989,20(1):84-91.
[10]Feng XD,Yang S,Liu J,et al.Electroacupuncture ameliorates cognitive impairment through inhibition of NF-κB-mediated neuronal cell apoptosis in cerebral ischemia-reperfusion injured rats[J].Mol Med Rep,2013,7(5):1516-1522.
[11]Liang XH,Jackson S,Seaman M,et al.Induction of autophagy and inhibition of tumorigenesis by beclin1[J].Nature,1999,402(6762):672-676.
[12]Vanderlaag K,Su Y,Frankel AE,et al.1,1-Bis(3'-indolyl)-1-(p-substituted phenyl)methanes induce autophagic cell death in estrogen receptor negative breast cancer[J].BMC Cancer,2010,10:669.
[13]黄凤媛,李菲,黄贵华,等.自噬基因Beclin1启动子细胞自噬模型的建立[J].广州中医药大学学报,2015,32(2):325-329.
[14]梁巧青,冯震博.自噬基因Beclin1的研究现状[J].当代医学,2010,16(9):21-22.
[15]Xu F,Gu JH,Qin ZH.Neuronal autophagy in cerebral ischemia[J].Neurosci Bull,2012,28(5):658-666.
[16]付俊,尚海旭,贾弘禔,等.Beclin1与自噬及肿瘤的关系[J].生理科学进展,2012,43(2):155-158.
[17]杨扬,赵美,赵铭,等.细胞自噬与心血管疾病中炎症反应的相关性[J].心脏杂志,2015,27(3):340-343.
[18]陈晓龙,张明亮,余祖江,等.细胞自噬在肝脏疾病中作用机制的研究进展[J].河南医学研究,2015,24(4):70-72.
[19]Ding ZB,Shi YH,Zhou J,et al.Association of autophagy defect with a malignant phenotype and poor prognosis of hepatocellular carcinoma[J].Cancer Res,2008,68(22):9167-9175.
[20]王远航,夏盛源,王芳,等.自噬通路中mTOR、Beclin1与肿瘤关系研究的最新进展[J].基因组学与应用生物学,2015,34(8):1656-1662.
[21]冯晓东,王慧灵,陈卓,等.自噬在脑卒中后认知障碍中的作用研究[J].时珍国医国药,2017,28(7):1730-1732.
[22]Qin L,Wang Z,Tao L,et al.ER stress negatively regulates AKT/TSC/mTOR pathway to enhance autophagy[J].Autophagy,2010,6(2):239-247.
[23]胡伟,彭亚文,蒋帅,等.自噬在缺血性脑损伤中的作用研究进展[J].中华神经医学杂志,2015,5(14):529-531.
[24]Nishida K,Kyoi S,Yamaguchi O,et al.The role of autophagy in theheart[J].Cell Death Differ,2009,16(1):31-38.
[25]Lipton P.Ischemic cell death in brain neurons[J].Physiol Rev,1999,79(4):1431-1568.
[26]刘娇,冯晓东.电针百会、神庭穴配合康复训练治疗脑卒中后认知障碍临床研究[J].中医学报,2013,28(4):608-610.
猜你喜欢
上海针灸杂志(2022年10期)2022-10-28
世界科学技术-中医药现代化(2021年8期)2021-12-21
昆明医科大学学报(2021年8期)2021-08-13
天津医科大学学报(2021年3期)2021-07-21
昆明医科大学学报(2020年11期)2020-12-28
中华养生保健(2020年4期)2020-11-16
养生保健指南(2019年3期)2019-12-16
农村百事通(2019年2期)2019-03-26
婚育与健康(2019年1期)2019-03-18
浙江医学(2018年16期)2018-09-08
