
4个东南亚国家水稻种质的表型多样性分析
2018-01-08 03:46张晓丽吕荣华唐茂艳梁天锋高国庆
西南农业学报 2017年12期
张晓丽,吕荣华,王 强,唐茂艳,陈 雷,3,郭 辉,梁天锋,高国庆*
(1.广西农业科学院水稻研究所/广西水稻遗传育种重点实验室,广西 南宁 530007;2. 广西农业科学院国际合作处,广西 南宁 530007;3. 广西大学农学院,广西 南宁 530004)
4个东南亚国家水稻种质的表型多样性分析
张晓丽1,吕荣华2,王 强1,唐茂艳1,陈 雷1,3,郭 辉1,梁天锋1,高国庆1*
(1.广西农业科学院水稻研究所/广西水稻遗传育种重点实验室,广西 南宁 530007;2. 广西农业科学院国际合作处,广西 南宁 530007;3. 广西大学农学院,广西 南宁 530004)
【目的】对水稻种质资源的多样性进行评价,为指导育种中优势亲本的选择和利用、提高选育效率和基因开发提供参考。【方法】采用统计分析和主成分分析,在广西南宁对298份东南亚国家水稻种质资源的表型多样性进行评估。【结果】不同数量性状变异系数最小的为结实率(6.51 %),最大的为有效穗数(34.86 %),株高、茎秆长和穗粒数等均具有较大的变异系数;Shannon-wiener多样性指数平均为0.7715;描述性性状多样性指数平均为0.2502,变化幅度为0.0769~0.4549;不同国家群体数量性状与描述性性状的多样性表现趋于一致,依次为菲律宾、柬埔寨、越南、缅甸。前8个主成分(主要性状包括株高、茎秆茎节包露、千粒重、穗粒数、茎秆节间色、颖尖色、芒长和叶片卷曲度)的累计贡献率达到67.99 %,代表了23个表型性状的大部分变异,可作为品种选育的参考依据。【结论】参试材料的表型多样性丰富,尤其是菲律宾和柬埔寨两个国家,应切实做好保护和利用工作。
种质资源;表型;多样性;东南亚;水稻
【研究意义】作物种质资源是作物品种选育和生产推广的源头,目前在水稻育种中,骨干亲本的单一化导致遗传基础狭窄,产量水平难以大幅度突破以及抗性遗传的脆弱性。对种质资源的多样性进行评价,可以深入指导育种中优势亲本的选择和利用,进而提高基因开发利用和选育效率。东南亚国家的气候多样化,地形较复杂,造就和孕育了丰富的水稻种质资源[1-2],研究东南亚国家的水稻种质资源,对有效管理和充分挖掘其遗传多样性对实现其品种改良均具有重要意义。【前人研究进展】目前,相关研究所采用的方法主要有多样性指数、变异系数及主成分分析等,对作物种质资源的表型进行评价[3-6]。有研究表明,不同地区间的种质资源遗传多样性高于同一地区内的不同品种间的多样性[7]。胡标林等[8]研究认为亚洲、非洲和大洋洲的水稻资源具有较丰富的表型遗传多样性,而不同性状的遗传多样性在洲际间表现不同,粒长宽比、碱消值、株高、粒宽、千粒重和淀粉含量 6 个性状具有很高的表型遗传多样性;贺治洲等[9]研究表明不同来源的热带水稻种质各性状变异范围较大,遗传多样性指数较高研究。【本研究切入点】明确东南亚部分国家水稻种质资源的遗传多样性,可为东南亚水稻种质资源的利用和保护提供依据,但目前针对东南亚国家水稻种质资源多样性的研究相对较少。【拟解决的关键问题】引进部分东南亚国家的298份水稻种质资源,进行表型多样性分析,明确四个东南亚国家水稻种质的表型变异情况,加强对其遗传基础的了解,为水稻育种的亲本选择提供依据。
1 材料与方法
1.1 试验材料
供试的298份水稻种质资源,为广西农业科学院水稻研究所保存的东南亚水稻种质资源,其中,来自菲律宾236份,柬埔寨28份,越南27份,缅甸7份。
1.2 试验方法
试验在广西农业科学院水稻研究所试验田(东经108°14′51″,北纬22°50′54″)进行,对298份东南亚水稻种质资源的23个表型性状进行调查。每份材料种植5行,每行20株,单株种植,种植规格16 cm×20 cm,常规的水肥管理和病虫害防治。性状鉴定和各项指标记录按照《水稻种质资源描述规范和数据标准》[10]进行,全生育期随机抽取典型性状的植株5株,调查、测量和记录23项农艺性状,性状的选择首先考虑的是育种利用以及性状测量的准确可靠性,从而保障了整体数据的精准可靠性[11]。主要包括数量性状10个:株高、穗长、茎秆长、有效穗数、穗粒数、结实率、千粒重、谷粒长度、谷粒宽度、长宽比;描述性性状13个:种皮颜色、叶片卷曲度、茎秆角度、剑叶长度、剑叶宽度、茎秆节间色、茎秆茎节包露、芒长、芒色、颖尖色、颖色、倒伏性、粘糯性。
1.3 数据标准化

1.4 数据分析

2 结果与分析
2.1 描述性性状的遗传多样性分析
对298份东南亚国家的水稻种质资源的描述性性状进行赋值标准化(表1),并对其表型进行统计分析。结果表明,各个性状在各描述级别上基本上均有分布,但不均匀(表2)。由表2分析可知东南亚国家水稻种质资源13个描述性性状均表现出一定的相对集中性,例如种皮色有4种表型,以白色所占比例最高;叶片卷曲度有4种表型,以不卷或卷度很小所占比例最高;茎秆角度有3种表型,以直立所占比例较高等同时,分析可得,不同性状的遗传多样性程度不同,其表型遗传多样性指数的平均值为0.2505,变化幅度为0.0769(芒长)~0.4549(剑叶长度),其他指数的遗传多样性大小排序为:芒色(0.4472)>茎秆节间色(0.4088)>叶片卷曲度(0.3306)>颖尖色(0.3254)>茎秆茎节包露(0.2916)>剑叶宽度(0.2214)>茎秆角度(0.1868)>倒伏性(0.1686)>粘糯性(0.1181)>颖色(0.1138)>种皮(0.109)。
2.2 数量性状的表型多样性分析
从表3可知,东南亚水稻种质资源数量性状的表型变异明显,多样性丰富,变异范围较大,10个数量性状的变异系数为6.51 %~34.86 %,平均值为19.90 %,以有效穗数最高,结实率最低,除结实率之外其余性状的变异系数均在10 %以上,株高、茎秆长、穗粒数的变异系数超过20 %,说明这些性状的变异幅度较大。10个数量性状的平均表型遗传
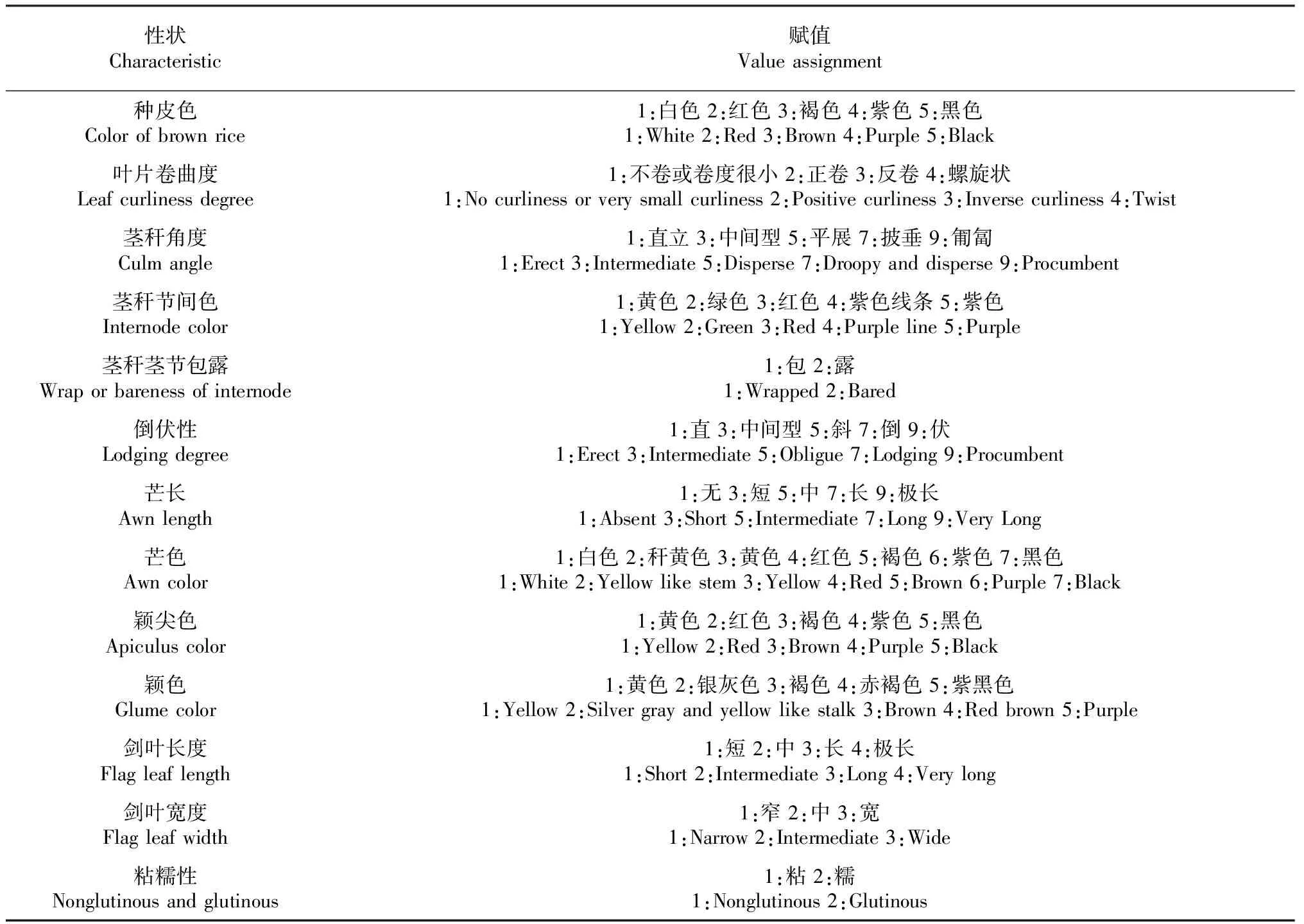
表1 水稻描述性性状的赋值

表2 东南亚水稻种质资源13个描述型性状的频率分布及多样性指数
多样性指数为0.7715,谷粒宽度多样性指数最高(0.9073),有效穗数多样性指数最低(0.0103),其余性状中,株高、千粒重、谷粒长度、茎秆长、结实率、长宽比、穗粒数的多样性指数均值0.84以上,说明

表3 东南亚水稻种质资源表型多样性统计分析
参试材料的如上数量性状的遗传多样性较为丰富。
2.3 不同国家水稻种质资源群体间的表型差异
2.3.1 描述性性状的表型差异 从13个描述性性状多样性指数分析(表4)可知,同一性状在不同国家水稻种质资源之间的遗传多样性指数存在差异。越南国家参试材料中,其种皮色遗传多样性指数较大(0.2184),柬埔寨参试材料中,其倒伏性、芒长、颖尖色、颖色的遗传多样性指数相对较大,分别为0.3378、0.1775、0.4499、0.2560。菲律宾参试材料中,其叶片卷曲度、茎秆节间色、茎秆茎节包露、芒色、剑叶长度的遗传多样性指数相对较大,分别为0.3524、0.3363、0.2904、0.4151、0.4545。缅甸参试材料中,其剑叶宽度的遗传多样性指数相对较大(0.2598)。采用各性状多样性指数的平均值来评估不同国家水稻种质资源的遗传多样性,4个国家的水稻种质资源多样性指数为0.0993~0.2562,柬埔寨(0.2562)>菲律宾(0.2326)>越南(0.1717)>缅甸(0.0993),说明柬埔寨水稻种质资源的多样性较为丰富,其次是菲律宾、越南、缅甸。
2.3.2 数量性状的遗传差异 由表5可知,各数量性状平均值在4个不同东南亚国家水稻种质资源间存在显著差异,越南参试材料中,株高、茎秆长和谷粒长度等3个性状指标均为最高,分别为137.67、114.16、4.46 mm;柬埔寨参试材料中,有效穗数、结实率、千粒重、谷粒宽度等4个性状指标均为最高,分别为9.57、91.03 %、28.37 g、1.74 mm;缅甸参试材料中,穗长、穗粒数、长宽比等3个性状指标均为最高,分别为23.98 cm、184.91个、3.19。

表4 不同地理来源水稻种质的13个描述性性状的遗传多样性指数

表5 东南亚不同地理来源水稻种质资源表型多样性分析
对不同国家水稻种质资源各性状的变异系数平均值进行分析(表5),4个国家的水稻种质资源群体的平均变异系数大小依次为菲律宾(19.75 %)>柬埔寨(18.56 %)>越南(13.61 %)>缅甸(11.25 %)。根据4个不同国家水稻种质资源群体单个性状变异系数来分析(表5),越南群体的千粒重的变异系数高(18.75),可选择性强;柬埔寨群体的株高、茎秆长、穗粒数的变异系数较高,分别为24.83 %、27.52 %、31.4 %。菲律宾群体的穗长、有效穗数、谷粒宽度、长宽比的变异系数高,分别为19.8 %、35.55 %、12.79 %、18.88 %,性状变异较大。
对4个不同来源群体的水稻种质资源多样性指数进行比较,菲律宾国家最高(236份、0.83),其次是柬埔寨群体(28份、0.78),这两个群体具有丰富的遗传多样性,越南群体(27份、0.72)排在第3位,缅甸群体(7份、0.57)排在第4位。
2.4 表型性状主成分分析
分析结果表明前8个主成分的累计贡献率达到67.99 %,说明这8个主成分包含了所有表型的67.99 %的内容,为水稻表型特征的重要主成分。由表6可知,第1主成分的特征根植为3.578,贡献率为16.26 %。最高载荷为株高,种皮色为最大副载荷,特征向量关系表明株高越高,种皮颜色越浅,千粒重越小;第2主成分的特征根值为3.109,贡献率为14.13 %,茎秆茎节包露具有最高载荷,谷粒宽度为最大负载荷,说明茎秆包裹越严重,谷粒越细长;第3主成分的特征根值为1.892,贡献率为8.60 %,千粒重和谷粒长度载荷值较高,由向量关系可知千粒重大、谷粒长的参试品种有效穗数较少;第4主成分的特征根值为1.584,贡献率为7.20 %,其中穗粒数贡献率最大,谷粒长度为最大负值,说明穗粒数越多,谷粒越短;第5主成分的特征根值为1.386,贡献率为6.30 %,贡献率最大的是茎秆节间色,向量关系表明,茎秆节间色越深的品种,颖色越浅;第6主成分的特征根值为1.229,贡献率为5.59 %,茎秆节间色、颖尖色及颖色贡献率相对较大,芒长为最大负向载荷,说明芒长越短,茎秆节间色、颖尖色和颖色越深;第7主成分的特征根值为1.14,贡献率为5.18 %,芒长载荷值最大,根据特征向量关系,芒长较长的参试品种,其结实率较低;第8主成分的特征根值为1.038,贡献率为4.72 %,其中叶片卷曲度贡献率最大,对比各特征向量可知,叶片卷曲度越大,剑叶和穗子越短,有效穗数越少。
在水稻实际生产过程中,涉及产量的有效穗数、穗粒数、结实率、千粒重,以及涉及外观品质的长宽比、颖色、剑叶长度、谷粒长宽比等性状是选育优良品种时需要考虑的因素。综上所述,第3、4、8主成分的值越大,育种选择潜力越大。
3 讨 论
目前常用的研究种质资源遗传多样性的检测方式有表型性状分析、DNA分子标记、染色体分析等,其中表型性状同时具有稳定性和变异性,同时最为直观、简单和经济,因此被首选并广泛运用[8,14-15]。表型变异是环境和DNA变异互作的结果,当对大量材料进行遗传多样性分析时可以先采用表型性状进行初步分析,可同步结合分子标记检测进行深层次分析[16]。
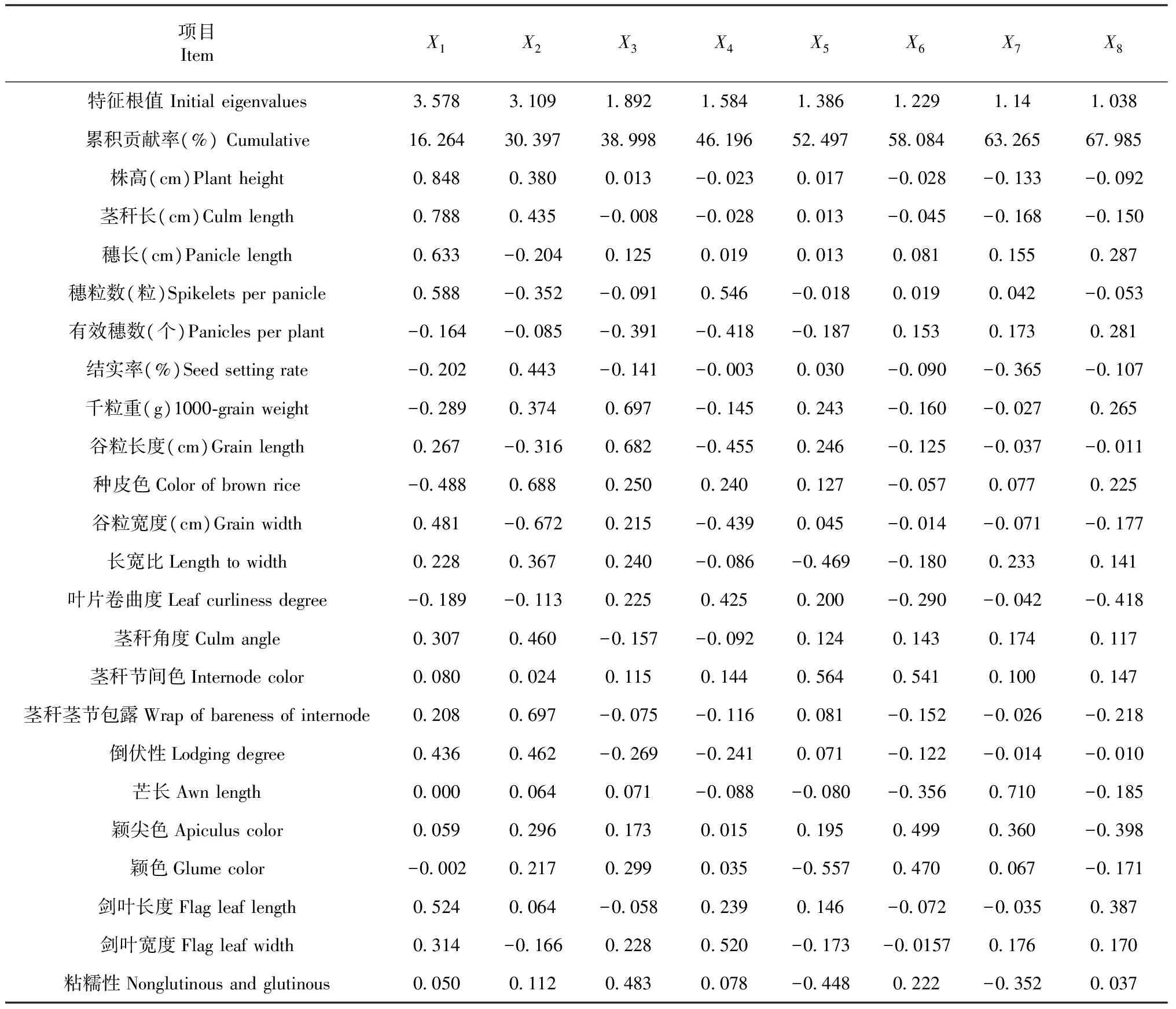
表6 东南亚水稻种质资源的主成分分析
本研究选用23个表型性状对298份东南亚水稻种质资源遗传多样性分析表明,10个数量性状均表现出不同程度的遗传变异,变异系数变幅在6.51 %~34.86 %,各个数量性状的变异程度上来看,有效穗数的变异程度最大,结实率的变异程度最小,同时参试材料在株高、茎秆长、穗粒数、有效穗数具有较大的变异幅度。其平均Shannon-wiener多样性指数为0.7715,7个性状的多样性指数均值0.84以上。同时13个描述性性状的多样性指数变化幅度在0.0769~0.4549,平均值为0.2502。所收集的东南亚水稻种质资源在表型上差异明显,具有较丰富的遗传多样性,可进一步从中挖掘优异基因,在育种上加以利用。
对比东南亚4个国家的水稻种质资源群体间的13个描述性性状多样性指数,同一性状在不同国家之间的遗传多样性指数存在差异,多样性指数在0.0993~0.2562之间,柬埔寨>菲律宾>越南>缅甸。说明柬埔寨水稻种质资源的多样性较为丰富,其次是菲律宾、越南、缅甸。同时10个数量性状在4个不同东南亚国家水稻种质资源间存在显著差异,表现与描述性性状趋于一致,其平均变异系数大小依次为菲律宾>柬埔寨>越南>缅甸;其多样性指数上来讲,菲律宾国家最高,其次是柬埔寨,这两个群体具有丰富的遗传多样性,应该做更规范的保护和利用。贺治洲等[9]对9个不同来源地的热带水稻资源多样性分析结果表明,不同来源的热带水稻种质各性状变异范围较大,各性状间存在显著差异,多样性指数在0.780~1.547,以泰国、柬埔寨和海南较高。这与本研究得出的结论相似。
主成分分析方法在将若干个形态标记经数据分析软件转化为少数几个毫无关联的主成分的基础上,又能较为全面地反映原来若干个形态标记的主要内容[17]。本研究通过SPSS统计软件,对298份东南亚水稻种质资源的22个农艺表型性状主成分分析,前8个主成分的累计贡献率达到67.99 %,可以很好地反映22个表型性状的大部分变异,为水稻品种选育中的性状相关性提供一定的参考依据。
东南亚国家高温高湿环境下,会产生大量抗性好,适应性强的种质资源,加之遗传多样性丰富,在目前遗传基础狭窄,水稻育种进展缓慢[18]的情况下,毋庸置疑,其水稻种质资源将会起到打破此瓶颈的作用。
4 结 论
本研究结果明确了东南亚部分国家水稻种质资源的遗传多样性程度,初步推断柬埔寨和菲律宾两个国家拥有较为丰富的遗传变异,应切实做好保护和利用工作。
[1]Glaszmann J C, Kilian B, Upadhyaya H D, et al. Accessing genetic diversity for crop improvement[J]. Current Opinion in Plant Biology, 2010, 13(2): 167-173.
[2]Zhao K, Tung C W, Eizenga G C, et al. Genome-wide association mapping reveals a rich genetic architecture of complex traits inOryzasativa[J]. Nat Commun, 2011, 467(2): 1-10.
[3]李振姣,马斯霜,部丽群,等. 宁夏外引水稻种质资源表型性状遗传多样性分析[J]. 江苏农业科学, 2016, 44(8): 117-121.
[4]蒋会兵,宋维希,矣 兵,等. 云南茶树种质资源的表型遗传多样性[J]. 作物学报, 2013, 39(11): 2000-2008.
[5]王海平,李锡香,沈 镝,等. 基于表型性状的中国大蒜资源遗传多样性分析[J].植物遗传资源学报, 2014, 15(1): 26-33.
[6]代攀虹,孙君灵,何守朴,等. 陆地棉核心种质表型性状遗传多样性分析及综合评价[J].中国农业科学, 2016, 49(19): 3694-3708.
[7]Xing H W, Xiao P Y, Han Y Y, et al. Temporal changes in SSR allelic diversity of major rice cultivars in China[J]. Journal of Genetics & Genomics, 2009, 36(6): 363-370.
[8]胡标林,万 勇,李 霞,等. 水稻核心种质表型性状遗传多样性分析及综合评价[J]. 作物学报, 2012, 38(5): 829-839.
[9]贺治洲,尹 明,谢振宇,等. 热带水稻优异种质资源的表型遗传多样性分析[J].热带农业科学,2014,34(9):37-42.
[10]韩龙植,魏兴华. 水稻种质资源描述规范和数据标准[M]. 北京:中国农业出版社, 2006: 3-120.
[11]王海岗,贾冠清,智 慧,等. 谷子核心种质表型遗传多样性分析及综合评价[J]. 作物学报, 2016, 42(1): 19-30.
[12]杨秀丽,宁东贤,武银玉,等. 花生种质主要数量性状的遗传多样性和相关性分析[J].中国农学通报, 2015, 1(9): 82-87.
[13]王 兵,郑秋红,郭 浩. 基于Shannon-wiener指数的中国森林物种多样性保育价值评价方法[J]. 林业科学研究, 2008, 21(2): 268-274.
[14]刘新龙,蔡 青,吴才文,等. 甘蔗品种资源的表型遗传多样性[J]. 生物多样性, 2010, 18(1): 37-43.
[15]蔡一林,刘志斋,王天宇,等. 国内部分玉米地方品种的品质与农艺性状的表型多样性分析[J]. 植物遗传资源学报, 2011, 12(1): 31-36.
[16]马斯霜,李振姣,赵 璐,等.宁夏水稻地方品种与自育品种表型性状遗传多样性分析[J].西北农业学报, 2017, 26(2): 216-226.
[17]贺治洲,尹 明,谢振宇,等. 热带水稻优异种质资源的表型遗传多样性分析[J]. 热带农业科学, 2014, 34(9): 37-42.
[18]李慧峰,陈天渊,黄咏梅,等. 甘薯种质资源形态标记遗传多样性分析[J]. 西南农业学报, 2015, 28(6): 2401-2407.
[19]孙希平,杨庆文.中国与东南亚三国(越、老、柬)普通野生稻遗传多样性的比较研究[J].作物学报, 2009, 35(4): 679-684.
PhenotypicDiversityEvaluationsofRiceGermplasmfromFourSoutheastAsiaCountries
ZHANG Xiao-li1, LV Rong-hua2, WANG Qiang1, TANG Mao-yan1, CHEN Lei1,3, GUO Hui1, LIANG Tian-feng1, GAO Guo-qing1*
(1.Rice Research Institute, Guangxi Academy of Agricultural Sciences, Guangxi Academy of Agricultural Sciences/Guangxi Key Laboratory of Rice Genetics and Breeding, Guangxi Nanning 530007,China;2.Guangxi Academy of Agricultural Sciences, Guangxi Nanning 530007,China;3.Agriculture College of Guangxi University, Guangxi Nanning 530004,China)
【Objective】Evaluation of genetic diversity could be beneficial to guide the selection and utilization of dominant parents in breeding, and further improve choose efficiency and gene exploit.【Method】298 rice germplasm genetic diversity were evaluated in Guangxi Zhuang Autonomous Region Nanning city based on the phenotypic character through usual statistical analysis and principal component analysis methods.【Result】The coefficient of variation of quantitative traits ranged from 6.51 % (seed rate) to 34.86 % (panicle number), while plant height, stalk length and grain number have also greater variation range.The average Shannon-wiener diversity index was 0.7715. The average diversity index of descriptive characters was 0.2502 ranged from 0.0769 to 0.4549. We compared genetic diversity from different countries, the result indicated the performance characteristics of quantitative traits and descriptive characters were consistent,the order was Philippines, Cambodia,Vietnam and Myanmar.Besides, the first eight principal components (plant height, wrap of bareness of internode, 1000-grain weight, spikelets per panicle, internode colorm, apiculus color and awn length)of the contribution rate was 67.99 %, it indicated that the first eight principal component contained all phenotypic character to 67.99 % of the content. At the same time, it could offer some references to rice variety breeding.【Conclusion】These resources have abundant genetic diversity especially, Philippines and Cambodia’s rice germplasm resources and should be better protected and utilized.
Germplasm resources; Phenotypic; Genetic diversity; Southeast Asia; Rice
1001-4829(2017)12-2617-07
10.16213/j.cnki.scjas.2017.12.001
2017-07-18
国家自然科学基金项目(31560363);广西科学研究与技术开发计划项目(桂科合15104001-27);广西自然科学基金项目(2016GXNSFBA380171);广西农业重点科技计划项目(201530);广西农业科学院科技成果转化项目(2017NZ03);广西农业科学院基本科研业务专项(桂农科2017YM17)
张晓丽(1982-),女,山西临汾人,农艺师,主要从事东南亚水稻种质资源研究工作,E-mail:xiaozhu8209@126.com,*为通讯作者:高国庆,E-mail:gqgao@gxaas.net。
S511.033
A
(责任编辑温国泉)
猜你喜欢
江西农业大学学报(2022年3期)2022-07-07
作文小学中年级(2020年9期)2020-12-29
作文·小学低年级(2020年9期)2020-11-30
农业机械学报(2020年11期)2020-11-25
农业机械学报(2019年4期)2019-04-29
作文小学中年级(2019年5期)2019-01-10
文苑(2018年18期)2018-11-08
现代园艺(2017年21期)2018-01-03
中国男科学杂志(2016年5期)2016-12-01
中国康复理论与实践(2015年10期)2015-12-24
