
烟草反义Mlo基因表达载体的构建
2018-01-05 00:35邬晓勇时羽杰王跃华孙雁霞
成都大学学报(自然科学版) 2017年4期
邬晓勇,时羽杰,李 杰,罗 倩,王跃华,唐 媛, 孙雁霞
(成都大学 药学与生物工程学院,四川 成都 610106)
烟草反义Mlo基因表达载体的构建
邬晓勇,时羽杰,李 杰,罗 倩,王跃华,唐 媛, 孙雁霞
(成都大学 药学与生物工程学院,四川 成都 610106)
获得烟草反义Mlo基因的植物表达载体pBI 121-Mlo,为进行烟草遗传转化,获得该基因表达的缺陷型植株打下基础.从烟草叶片中提取总RNA,利用RT-PCR技术扩增得到Mlo基因的cDNA,以此为模板设计反义引物,通过PCR扩增出反义Mlo基因,将此反义Mlo基因与T载体连接,测序正确后再将此反义片段与植物表达载体pBI 121连接,构建烟草反义Mlo基因的植物表达载体pBI 121-Mlo.经Kan选择筛选出反义重组菌落,碱裂解法小量提取质粒后,用XbaI和BamHI双酶切后再进行电泳鉴定.结果表明,目的基因已与植物表达载体pBI 121连接成功.成功构建了烟草反义Mlo基因表达载体pBI 121-Mlo.
Mlo基因;pBI 121Mlo;RT-PCR;反义片段;双酶切
0 引 言
植物在长期的进化过程中形成了一系列复杂而严密的防御机制,而使自身免受病原物的侵害,这涉及到一系列抗病相关基因的表达和调控,其中,Mlo基因就是植物抗病基因中的重要成员之一[1-2].研究发现,白粉病是由白粉菌引起的真菌性病害,能侵染多种单子叶植物和双子叶植物[3-4],野生型Mlo基因是大麦抗白粉病的负调控因子,可赋予大麦对白粉菌的广谱抗性[5].
目前,科研人员已经对拟南芥、水稻、玉米和杨树中的Mlo基因家族进行了深入研究[6],而对烟草的相关研究未见报道.对此,本研究以构建烟草Mlo基因反义表达载体为基础,鉴定Mlo基因是否是抗白粉病的负调控因子,拟对其在烟草抗病方面的负调控作用提供实验依据.
1 实验与方法
1.1 材 料
实验所用材料包括:红花大金元品种烟草的无菌苗叶片;DL2000 DNA Marker、GREENspspin多糖多酚类植物RNA快速提取试剂盒、高纯度质粒提取试剂盒、pBLUE-T载体、小量琼脂糖凝胶DNA回收试剂盒,购于北京庄盟生物有限公司,TIANgen midi purification kit试剂盒,购自于大连TaKaRa公司;JM109菌株由北京林业大学生物科学与技术学院陆海教授惠赠.
1.2 仪 器
实验所用仪器包括:5020型基因扩增仪(Thermo公司),PAC300型电泳仪(BIORAD公司),MDF-382E型超低温冰箱(SANYO公司),Eppendorf微量移液器、凝胶成像系统(BIO-RAD公司),ZF-90型暗箱式紫外透射仪(上海顾村电光仪器厂),FA2004型电子天平(上海超平科学仪器有限公司),AIRTECH超净工作台(安泰公司).
1.3 方 法
1.3.1 烟草Mlo基因的克隆.
1)烟草总RNA提取.通过GREENspspin多糖多酚类植物RNA快速提取试剂盒提取烟草的总RNA,并用琼脂糖凝胶电泳检测RNA提取情况.
2)NA反转录获得cDNA.根据提取到的RNA,用TIANgen midi purification kit试剂盒转录合成cDNA,-20 ℃保存备用.
反转录具体步骤:①在Ep管中依次加入下列溶液,Oligo(dT)18 Primer,1 μ、烟草RNA样品,10 μL.此操作可在PCR仪中完成,70 ℃后立即置于冰上冰浴2~5 min.②依次加入下列试剂,AMV Reverse Transcriptase 2 μL、5*Reverse Transcriptase Buffer 5 μL、dNTP Mixture(10 mM each) 2 μL、ddH2O 5 μL.
3)cDNA扩增成烟草Mlo基因.以反转录获得的cDNA为模板,利用设计的引物扩增烟草Mlo基因.
①上游引物(P1):
CACGGATTAGCTAAAGGTTAGGGC
②下游引物(P2):
GCTGAATTACTCCCAATCGGCATC
PCR反应体系(25 μL)如表1所示.

表1 PCR反应体系
PCR反应条件为:首先,94 ℃预变性5 min;其次,94 ℃变性30 s,56 ℃退火30 s,72 ℃延伸60 s,30个循环;最后,72 ℃延伸10 min,获得烟草Mlo基因的DNA序列.
4)烟草Mlo基因与T载体连接.扩增产物经1%琼脂糖电泳检测后,用小量琼脂糖凝胶DNA回收试剂盒回收目的片段,回收的Mlo基因与pBLUE-T载体连接后由北京华大基因公司进行序列测定.
1.3.2 烟草Mlo基因反义表达载体构建.
1)PCR获得Mlo基因的反义片段.在成功克隆到与预期大小一致的Mlo基因后,通过反义引物克隆Mlo基因的反义片段,进一步构建反义表达载体.设计如下2条反义引物,同时为了将其与植物表达载体pBI 121连接,设计引物时还引入了带有保护碱基的酶切位点.
GCTCTAGAGTTAGCAGCAGCTTGTGGACAAC
画线部分为Xba I酶切位点.
②下游引物:
CGGGATCCTGAGATCTGTGGAATCCATAG
画线部分为BamH I酶切位点.
PCR反应体系(25 μL)如表2所示.

表2 PCR反应体系
扩增程序为:首先,94 ℃预变性5 min;其次,94 ℃变性45 s,58 ℃退火45,72 ℃延伸60 s,30个循环;最后,72 ℃延伸10 min,获得烟草Mlo基因的反义片段后,采用1%琼脂糖凝胶电泳检测反义片段是否扩增成功.
小窍门:瘦猪肉用蚝油腌制,因为蚝油比酱油含有更多的锌。各种新鲜、天然的蔬菜瓜果、不加糖的鲜榨果蔬汁、红薯、煮玉米、坚果类。当然,那些过度加工含有太多食品添加剂的零食就属于限制级的零食,尽量少给孩子吃或者不吃,这些食物包括:膨化食品、巧克力派、糖果、炸鸡块、炸鸡翅、奶油夹心饼干、方便面、奶油蛋糕、水果罐头、果脯、蜜枣脯、全脂或低脂炼乳、炸薯片和炸薯条、高糖分汽水或可乐等碳酸饮料、较甜的雪糕、冰激凌等。
2)反义片段测序.扩增出的反义片段经1%琼脂糖电泳检测后,用小量琼脂糖凝胶DNA回收试剂盒将此片段回收后与pBLUE-T载体连接,命名为pBLUE-Mlo,由北京华大基因公司进行序列测定.
3)反义Mlo基因与pBI 121连接.利用限制性内切酶Xba I、BamH I分别对反义克隆载体pBLUE-Mlo及植物表达载体pBI l21进行37 ℃过夜双酶切.酶切反应体系(20 μL)如表3、表4所示.

表3 反义pBLUE-Mlo双酶切体系
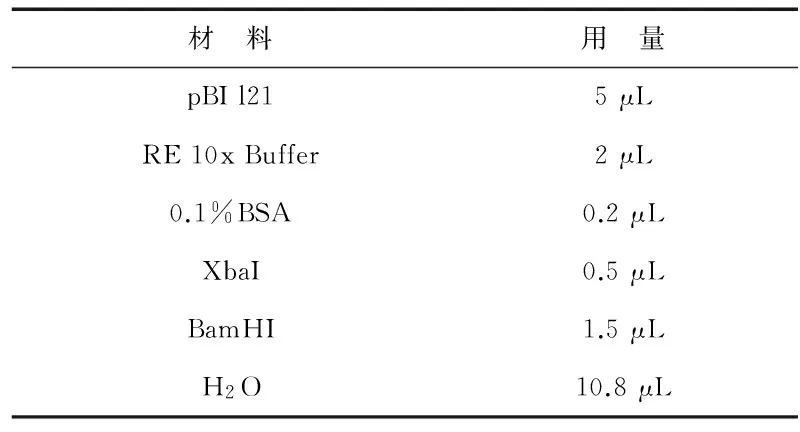
表4 pBI 121双酶切体系
将反义Mlo基因片段定向连接到植物表达载体pBI 121中,命名为pBI 121Mlo(14 ℃连接至少48 h),连接体系(10 μL)如表5所示.

表5 反义Mlo与pBI 121连接体系
反义Mlo基因片段与表达载体pBI 121连接重组载体pBI 121Mlo的构建过程如图1所示.
4)大肠杆菌转化与反义表达载体的鉴定.将pBI 121-Mlo热激法转化大肠杆菌JM109,菌液涂布于含Kan的LB平板上,37 ℃倒置培养14~16 h.挑取培养基上的单菌落接种于LB液体培养基中,在37 ℃的振荡培养箱中培养12 h,用质粒提取试剂盒提取质粒.

图1烟草反义表达载体构建过程
5)经Kan选择筛选出反义重组菌落,利用碱裂解法小量提取质粒进行电泳后,用BamH I和XBa I对质粒进行双酶切,酶切后用1%的琼脂糖凝胶电泳鉴定是否反义表达载体pBI 121-Mlo构建成功.
2 结果与分析
2.1 烟草总RNA提取结果
通过GREENspspin多糖多酚类植物RNA快速提取试剂盒提取烟草的总RNA,并用琼脂糖凝胶电泳检测RNA提取情况,电泳图如图2所示.

图2 RNA电泳图
由图2可以看出,2个烟草样品分别有RNA的28 s与18 s 2个条带,说明其RNA提取是成功的.
2.2 烟草Mlo基因克隆结果
2.2.1 RT-PCR结果检测.
将提取到的烟草RNA通过反转录得到烟草cDNA.利用设计的P1、P2引物经PCR反应扩增目的片段,扩增产物经1%琼脂糖电泳,结果如图3所示.
由图3可知,扩增产物的目的片段均在1 000 bp左右,与预期结果951 bp较为符合.
2.2.2 PCR产物与T载体连接结果.
PCR产物与T载体连接结果如图4所示.

图3烟草Mlo基因RT-PCR扩增产物电泳图

图4基因片段与T载体连接图
由图4可知,样品1、6、8、10、11比其他剩余的样品分子量小,说明这5个样品中载体没与基因片段连接上,剩余的样品已连接成功.
2.2.3Mlo基因扩增产物测序结果.
Mlo基因扩增产物测序结果如下,

由产物测序结果可知,其31~945处的碱基为烟草Mlo基因编码的开放阅读框(ORF),大小为915 bp.下划线部分别为上游和下游引物的位置,加粗部分分别为起始和终止密码子.结果表明,从烟草中成功克隆到了与预期大小一致的Mlo基因.
利用上述获得的Mlo基因,由Bioedit软件得到Mlo基因的反义序列如下,

其中,引物A,带有Xba I酶切位点,为,
GCTCTAGAGTTAGCAGCAGCTTGTGGACAAC
引物B,带有BamH I酶切位点,为,
CGGGATCCTGAGATCTGTGGAATCCATAG
2.3 反义片段扩增结果
反义片段扩增结果如图5所示.

1,2为反义扩增样品
图5反义片段扩增电泳图
由图5可知,样品电泳片段大小在500~1 000 bp之间,由Bioedit软件得到Mlo基因的反义序列大小是577 bp,表明反义片段扩增成功.切下扩增的条带用小量琼脂糖凝胶DNA回收试剂盒回收,并与T载体进行连接,再测序.
2.4 反义片段载体连接结果
反仪片段载体连接结果如图6所示.
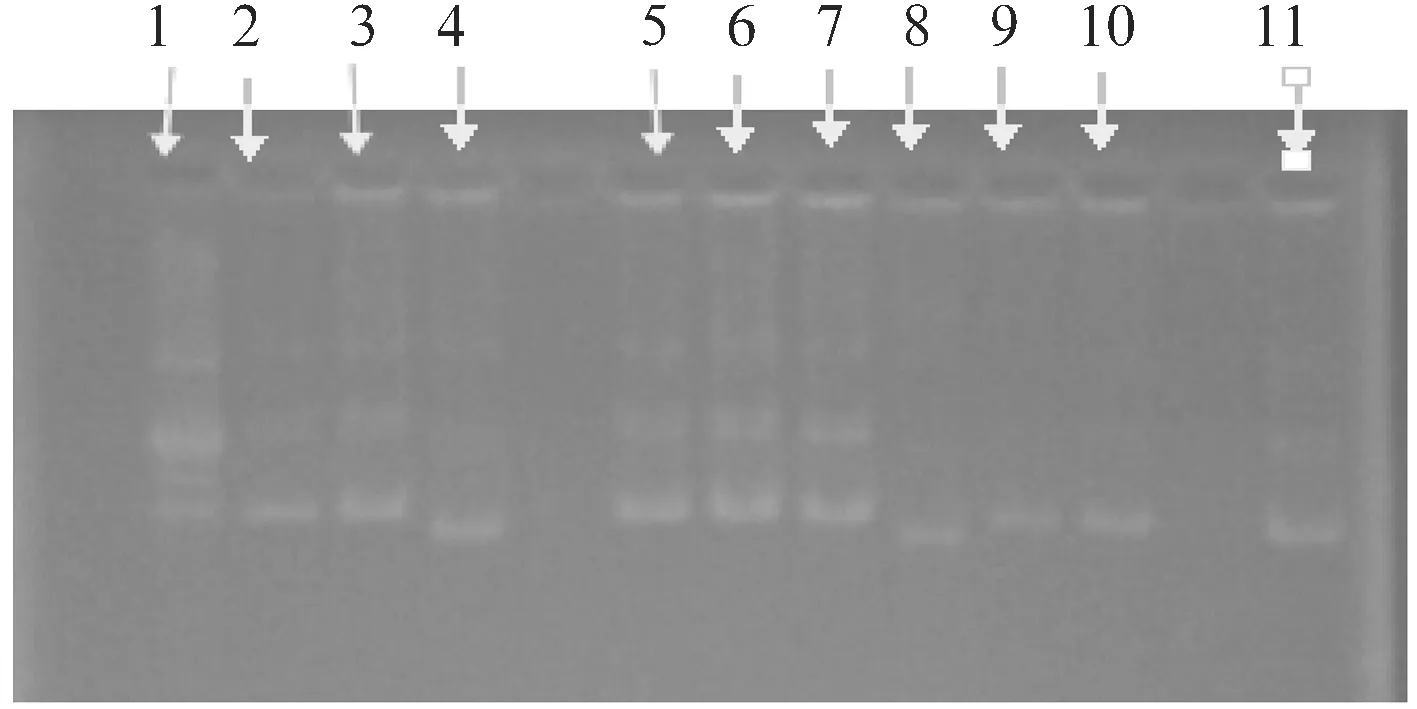
图6反义片段载体电泳图
由图6可知,除了第4、8、11以外,其他反义片段扩增成功.
2.5 反义片段测序结果
反义片段测序结果如下,下划线部分为设计的反义引物.

2.6 构建反义表达载体双酶切电泳结果
用已通过连接的pBI 121-Mlo热激法转化大肠杆菌JM109.经Kan筛选出反义重组菌落,利用碱裂解法小量提取质粒后进行电泳鉴定,结果如图7所示.

图7反义表达载体双酶切电泳图
因为载体PBI 121和反义片段具有相同的酶切位点.由图7可以看出,样品1和样品2分别产生了0.6 kb和2.4 kb的基因片段,说明目的片段成功插入PBI 121.此表明,烟草Mlo基因反义表达载体构建成功,图7中PBI 121-Mlo为已构建成功的反义表达载体.
3 结 论
本研究以烟草的无菌苗叶片为材料,利用RT-PCR方法成功克隆了一个Mlo基因,同时,通过反义引物克隆Mlo基因的反义片段,测序后成功找到设计的反义片断,并利用反义引物成功扩增了Mlo基因的反义片段.
反义片段与载体pBLUE-T连接后转入大肠杆菌,Kan实验提质粒后,用反义片段构建的pBI 121载体经Bam HI和XBaI双酶切,其电泳图可以看见样品分为0.6 kb和2.4 kb两类大小的目的片段,说明插入成功,即成功构建了烟草Mlo基因反义表达载体.
利用反义RNA技术[10]获得烟草Mlo基因的缺陷性植株的关键是获取烟草反义Mlo基因.本研究从烟草叶片中提取总RNA,利用RT-PCR技术扩增得到Mlo基因的cDNA,并以此为模板,设计反义引物通过PCR扩增出反义Mlo基因.其中反义Mlo基因的长短选择极为重要,因为反义Mlo基因过短可能无法起到抑制Mlo基因表达的作用,而过长则会给连接载体造成困难.对此,本研究选择了长为577 bp的片段进行引物设计和反义序列扩增,以确保反义Mlo基因既能很好地与载体连接,又能抑制Mlo基因的表达.
由反义Mlo基因与植物表达载体pBI 121进行连接后的产物的双酶切电泳鉴定结果可知,本研究已成功构建了烟草反义Mlo基因表达载体pBI 121-Mlo,并有望将携带反义Mlo基因的植物表达载体质粒导入根癌农杆菌,对烟草进行遗传转化研究,获得该基因表达的缺陷型植株,进而研究其在烟草抗病方面的负调控作用.
[1]邬晓勇,孙雁霞,何钢,等.烟草Mlo基因的克隆及其序列特性分析[J].烟草科技,2010,54(6):63-67.
[2]邬晓勇,孙雁霞,何钢,等.一个玉米Mlo基因的电子克隆与生物信息学分析[J].玉米科学,2011,19(1):148-152.
[3]李凡,胡东维,陈锋,等.小麦TaMlo基因的原核表达、抗体制备及细胞化学分析(简报)[J].分子细胞生物学报,2005,38(6):550-553.
[4]罗臻,张敬泽,胡东维.大麦Mlo近等基因系与叶枯病菌互作的细胞学研究[J].植物病理学报,2009,39(1):362.
[5]Devoto A,Hartmann H A,Piffanelli P,et al.Molecularphylogenyandevolutionoftheplantspecificseven-transmembraneMolfamily[J].J Mol Evol,2003,56(1):77-88.
[6]丁时超,杜文,任建新,等.LC-MS/MS定量分析卷烟中的烟草特有亚硝胺(TSNAs)[J].中国烟草学报,2005,11(6):17-22.
[7]曹阳,袁澍,徐飞,等.美洲黑杨木质素合成关键基因的克隆及反义表达载体的构建[J].中南林业科技大学学报,2010,30(10):77-82.
[8]潘昱名,刘风珍,万勇善,等.花生磷酸烯醇式丙酮酸羧化酶基因的克隆及反义表达载体的构建[J].山东农业大学学报(自然科学版),2010,41(1):1-5.
[9]Gookin T E,Kim J,Assmann S M.Wholeproteomeidentificationofplantcandidateg-proteincoupledreceptorsinarabidopsis,riceandpoplar:computationalpredictionandin-vivoproteincoupling[J].Genome Biol,2008,9(1):120-131.
[10]王伏林,王远山,胡张华.反义RNA在植物基因工程中的应用[J].生物技术,2003,13(1):34-35.
ConstructionofTobaccoAnti-senseMloGeneExpressionVector
WUXiaoyong,SHIYujie,LIJie,LUOQian,WANGYuehua,TANGYuan,SUNYanxia
(School of Pharmacy and Bioengineering, Chengdu University, Chengdu 610106, China)
Tobacco anti-senseMlogene plant expression vector PBI 121-Mlowas constructed not only for tobacco genetic transformation,but also to lay the foundation for the defective plant gene expression.Total RNA was extracted from tobacco leaves and the cDNA ofMlowas obtained by PT-PCR amplification.The cDNA was served as a template to amplify the anti-senseMlogene by the anti-sense primers.Then the anti-senseMlogene was connected with T vector and sequenced and then the anti-sense fragment was connected to the plant expression vector named pBI 121.The plant expression vector of the tobacco anti-senseMlonamed pBI121-Mlowas constructed.The recombinant bacteria were selected through Kanamycin and a few plasmids were extracted by alkaline lysis method.The plasmid was identified by electrophoresis after double digestion by Xba I and BamH I.The results illustrated that the target gene was successfully connected to the plant expression vector pBI 121.The genetic expression vector named pBI 121-Mloof tobacco anti-senseMlowas constructed.
Mlogene;pBI 121-Mlo;RT-PCR;anti-sense fragment;double digestion
S572.032
A
1004-5422(2017)04-0333-05
2017-09-09.
成都大学药食同源植物资源开发四川省高校重点实验室开放基金(10Y201409)资助项目.
邬晓勇(1975 — ),男,博士,副教授,从事植物天然产物研究.
猜你喜欢
环球时报(2022-09-20)2022-09-20
太原理工大学学报(2021年6期)2021-11-25
小学生学习指导·低年级(2021年6期)2021-09-10
考试与评价·七年级版(2021年4期)2021-08-14
今日农业(2020年24期)2020-12-15
中国海洋大学学报(自然科学版)(2019年7期)2019-05-21
中国海洋大学学报(自然科学版)(2019年7期)2019-01-04
小学阅读指南·低年级版(2018年5期)2018-11-02
广州化工(2016年11期)2016-09-02
小资CHIC!ELEGANCE(2015年15期)2015-09-01
