
紫茎泽兰入侵对不同植物群落类型中昆虫多样性的影响
2017-12-29 07:00蒋智林邓丹丹刘万学万方浩李正跃
生态环境学报 2017年12期
蒋智林,邓丹丹,刘万学,万方浩,李正跃
1. 普洱学院,云南 普洱 665000;2. 云南农业大学农业生物多样性与病虫害控制教育部重点实验室,云南 昆明 650201;3. 植物病虫害生物学国家重点实验室//中国农业科学院植物保护研究所,北京 100094
紫茎泽兰入侵对不同植物群落类型中昆虫多样性的影响
蒋智林1,2*,邓丹丹1,2*,刘万学3,万方浩3,李正跃2*
1. 普洱学院,云南 普洱 665000;2. 云南农业大学农业生物多样性与病虫害控制教育部重点实验室,云南 昆明 650201;3. 植物病虫害生物学国家重点实验室//中国农业科学院植物保护研究所,北京 100094
紫茎泽兰Ageratina adenophora入侵对不同植物群落中昆虫群落的影响,以及不同植物群落抵御紫茎泽兰入侵的生态机制尚不清楚。本研究选取紫茎泽兰不同入侵程度和未入侵不同功能型植物群落样地,采用巴氏罐诱法、扫网法结合黄板法对各样地地表昆虫进行调查分析。在各样地共采集到昆虫标本12 099个,隶属于11目64科168种。其中,优势类群为膜翅目、同翅目、双翅目、缨翅目昆虫,占总种数的94.86%。随紫茎泽兰的入侵,群落中直翅目、缨翅目、同翅目、双翅目、鞘翅目、鳞翅目昆虫在群落中所占的比例呈先降低再升高的趋势,而膜翅目、半翅目昆虫在群落中所占比例呈先升高再降低趋势。紫茎泽兰群落中的同翅目、半翅目昆虫所占比例较不同功能型本地植物群落低,而膜翅目昆虫所占比例较其他群落高。随紫茎泽兰入侵程度的加重,昆虫多样性指数D、H、E均呈先降低后升高的趋势;昆虫优势度指数C则呈先升高再降低的趋势。多年生禾本科植物群落的昆虫各多样性指数均大于紫茎泽兰群落,与紫茎泽兰群落间的差异最小,且有较高的相似性(0.602),可能是对紫茎泽兰的入侵具有较强抵御作用的指示因子之一。
紫茎泽兰;昆虫多样性;多样性指数;昆虫群落组成;不同植物群落
在长期的自然进化过程中,生物与生物之间相互作用,形成了稳定的互作关系。外来植物入侵往往对土著生态系统的结构和功能产生不良影响。入侵植物作为初级生产者,直接影响本地植食性昆虫及其天敌,或通过影响本地植物而改变其营养特性和挥发物的释放,从而间接影响本地昆虫(Wu et al.,2009)。对紫茎泽兰(Ageratina adenophora)不同入侵程度的群落或生态系统生物多样性调查发现,该种的入侵会对入侵地植物多样性和节肢动物多样性产生严重的影响,引起生物多样性的降低(王文琪等,2009);其入侵通常会导致土壤动物类群总数、个体总数显著减少,这与紫茎泽兰的淋漓分泌物或者入侵生境的类型存在一定交互关系(刘志磊等,2006);而对黄顶菊(Flaveria bidentis)的入侵影响研究发现,该物种的入侵会打破入侵地群落本地昆虫与天敌昆虫的关系,建立新的复杂竞争、捕食和共生互利昆虫群落关系(杜喜翠等,2011);此外,也有研究发现,入侵植物能够与本地同属植物争夺传粉昆虫,导致本地植物授粉受到影响而不断衰退(Brown et al.,2001)。可见,入侵植物可通过直接或间接的关系影响昆虫群落,这种影响往往受制于外界条件而呈现出复杂性。
紫茎泽兰是一种恶性入侵有毒杂草,为菊科多年生丛生型半灌木草本植物,原产于南美洲的墨西哥至哥斯达黎加一带(刘伦辉等,1985),现已广泛分布于世界热带、亚热带的30多个国家和地区。该入侵杂草约于20世纪40年代由缅甸传入中国云南边境地区,现已在云南、贵州、四川、重庆、广西、西藏等地区广泛分布(高贤明等,2003),已给入侵地区的生态系统和环境造成了严重的危害(于文清等,2012)。一些研究者研究了入侵植物对土壤养分、酶活性、微生物的影响,调查了入侵植物对节肢动物、土壤动物等群落多样性的影响(Wu et al.,2009;张修玉等,2010;亓东明,2014;邓丹丹等,2015),而关于未被入侵的不同功能型植物群落对入侵植物的抵御机制研究尚少。植物功能型群落是基于植物的形态、生理、生活史或其他相关的生态系统过程以及与物种相联系的某些生物学特性划分的,是对特定环境因素有相似反应的不同植物种类的组合(秦浩等,2015)。本研究选取紫茎泽兰不同入侵程度及未入侵的不同功能型植物群落作为研究样地。入侵程度分为重度入侵、轻度入侵、未入侵植物群落,未入侵植物群落按照不同功能型划分为:多年生禾本科植物群落、一年生禾本科植物群落、多年生双子叶植物群落及一年生双子叶植物群落。采用巴氏罐诱法、扫网法结合黄板法采集各研究样地地表昆虫,分析紫茎泽兰入侵对不同研究样地地表昆虫的类群、数量、多样性的影响,为解析不同植物群落抵御紫茎泽兰入侵的生态机制,剖析紫茎泽兰成功入侵的机理、紫茎泽兰的防治以及被入侵生态系统的修复提供参考。
1 材料和方法
1.1 研究地点
研究区位于云南省澄江县麒麟山及周边区域(24°42′N,102°52′E),海拔 1957~2015 m。该区属于亚热带高原季风气候带,年降雨量 900~1200 mm,各月份间降雨量分布不均,干湿季节分明,每年5月中旬—10月中旬降水量占全年的 85%左右,为雨季;10月下旬—次年5月上旬降雨量相对较少,为旱季;年平均气温约 16.8 ℃,极端最高气温为 33.7 ℃,极端最低气温为-3.9 ℃。土壤类型为典型的南方红壤,属于滇中地区主要的土壤类型。该地区植物物种有入侵草本植物紫茎泽兰(A.adenophora),土著草本植物有艾蒿(Artemisia argyi)、龙牙草(Agrimonia pilosa)、酢浆草(Oxalis corniculata)、菅草(Themeda triandra)、茅草(Imperata cylindrica)、荩草(Arthraxon hispidus)、画眉草(Eragrostis pilosa)、小藜(Chenopodium serotinum)、牛筋草(Eleusine indica)和野燕麦(Avena fatua)等。
1.2 调查方法
根据紫茎泽兰和当地植物群落竞争演替的不同阶段,以及紫茎泽兰盖度和入侵时间的不同,将入侵地划分为重度入侵群落、轻度入侵群落、未入侵群落等 3种类型。划分依据如下:(1)重度入侵群落:紫茎泽兰为优势种群,盖度大于60%,入侵时间10年以上;(2)轻度入侵群落:紫茎泽兰与当地植物处于竞争生长,紫茎泽兰盖度为10%~30%,入侵时间 3年左右;当地植物盖度为30%~50%;(3)未入侵群落:紫茎泽兰无成株或仅有零星幼苗生长,盖度小于0.2%(牛红榜,2007)。重度入侵群落和轻度入侵群落中外来入侵植物只有紫茎泽兰,而本地的主要优势植物为多年生禾本科植物,其次为多年生双子叶植物。
进一步将未入侵地区的植物群落类型划分为:多年生禾本科植物为主的群落、一年生禾本科植物为主的群落、多年生双子叶植物为主的群落、一年生双子叶植物为主的群落等4种类型。多年生禾本科植物主要包括菅草、茅草等;一年生禾本科植物主要有荩草、画眉草、牛筋草、野燕麦等;多年生双子叶植物主要包括艾蒿、龙牙草、酢酱草等;一年生双子叶植物主要为小藜等。
2016年6—10月,在相同地理区域分别选取各群落类型研究样地5个,每个样地的面积约为0.1 hm2,每月调查生境中的地表昆虫种类及其数量。昆虫采集方法采用巴氏罐诱法(陷阱法)、扫网法结合黄板法。巴氏罐诱法:以上口径为11 cm,高为7.5 cm的塑料杯作为陷阱容器,杯壁上方1/4处打有小孔,以避免由于雨水过多使标本流失。所用引诱剂为醋、糖、酒精和水的混合物,质量比为2∶1:6∶16,并加入少量洗衣粉,去除表面张力。每个样地设6个陷阱(2×3排列),塑料杯内放入引诱剂40~60 mL,杯口与地面相平,放置3 d后进行一次性采集。将昆虫样本从陷阱中取出后,放入盛有75%酒精的标本瓶中保存,带回实验室进行分类鉴定。扫网法:采用“Z”型路线,各样地扫网50网,将捕得的昆虫置于毒瓶中毒死后,装入75%酒精的瓶内保存,带回实验室进行分类鉴定。黄板法:各样地按五点法挂黄板5块,黄板高度刚好高于植物群落高度,3 d后收集黄板带回实验室鉴定。种类鉴定的参考依据为:《昆虫分类》(郑乐怡等,1999)、《中国土壤动物检索图鉴》(尹文英等,1998)、《中国动物志》(中国科学院中国动物志编辑委员会,
2003)等。
1.3 数据处理
昆虫多样性测度指标选用 Margalef丰富度指数、Simpson优势度指数、Shannon-Wiener多样性指数、Pielou均匀度指数、Jaccard相似性指数。采用地表昆虫各种类的个体数来度量各调查样地中昆虫丰富度、优势度、多样性、均匀度及相似性。各指标计算公式如下:
Margalef丰富度指数D(张海周等,2009):

Simpson优势度指数C(Simpson,1949):

Shannon-Wiener多样性指数 H(Pielou,1975):

Pielou均匀度指数E(Pielou,1975):

Jaccard相似性指数q(张海周等,2009):

以上各式中,S为物种数,N为个体总数,Pi为第 i个类群的个体数占群落总个体数的比率,c为两群落的共有类群数,a和b分别为群落A和群落B的类群数。相似性指数q值为0~0.25为极不相似,0.25~0.50不相似,0.50~0.75相似,0.75~1极相似。
采用Excel软件对昆虫种类、数量进行统计;采用 SPSS(SPSS 16.0,Inc.,Chicago,USA)软件对昆虫多样性进行分析,不同类型植物群落间、紫茎泽兰不同入侵程度植物群落间昆虫多样性的差异性均分别采用单因子方差分析(SPSS,one-way ANOVA)。
2 结果与分析
2.1 不同植物群落中的昆虫群落组成
2.1.1 紫茎泽兰群落与不同功能型本地植物群落昆虫群落组成
由表1、表2可知,在紫茎泽兰群落与不同功能型植物群落中,共获得昆虫标本9508个,隶属于11目64科168种。优势类群为膜翅目(44.81%)、同翅目(23.75%)、双翅目(15.34%)、缨翅目(10.54%)昆虫,这4个目累计占总数的94.21%。此外,还有少量的鞘翅目、直翅目、螳螂目、脉翅目、鳞翅目、蜚蠊目、半翅目昆虫(表2)。膜翅目中,主要优势种有东方行军蚁(Dorylus orientalis)、长足切叶蚁(Aphaenogaster longipes)、广大腿小蜂(Brachymeria obscurata)、食蚜蝇姬蜂(Diplozon laetatorius)等;同翅目中,主要优势种有小绿叶蝉(Empoasca flavescens)、大青叶蝉(Tettigoniella viridis)、棉蚜(Aphis gossypii)等;双翅目中,主要优势种有食蚜蝇(Didea fasciata)、大灰食蚜蝇(Syrphus corollae)等;缨翅目中,主要优势种有食蚜蝇(Didea fasciata)、大灰食蚜蝇(Syrphus corollae)等;缨翅目中,主要优势种有花蓟马(Frankliniella intonsa)和八节黄蓟马(Thrips flavidalus)。
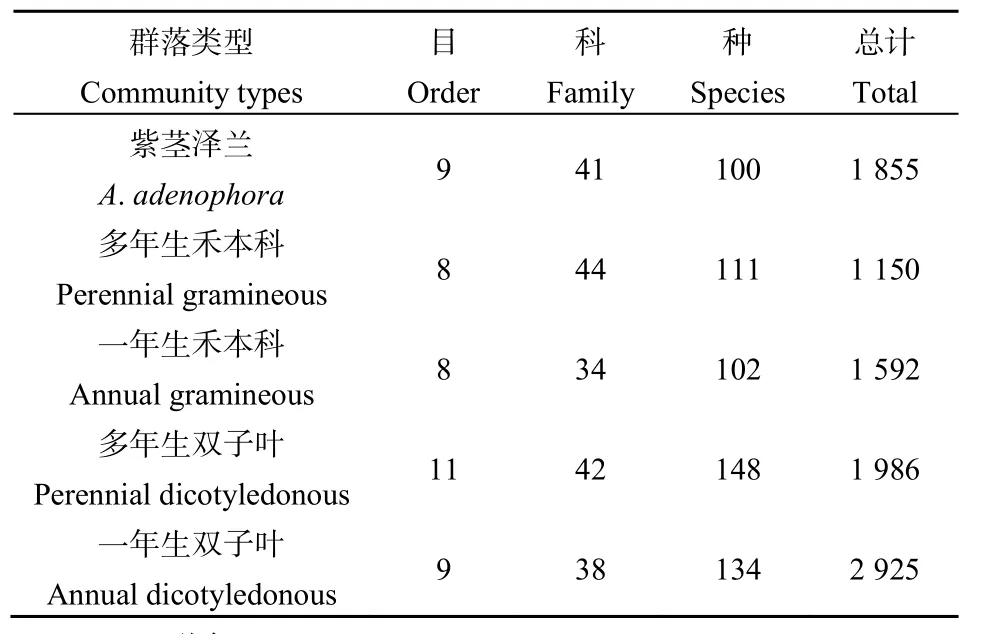
表1 紫茎泽兰群落与不同功能型植物群落间昆虫组成Table1 Compositions of insect in A. adenophora community and different function plant communities
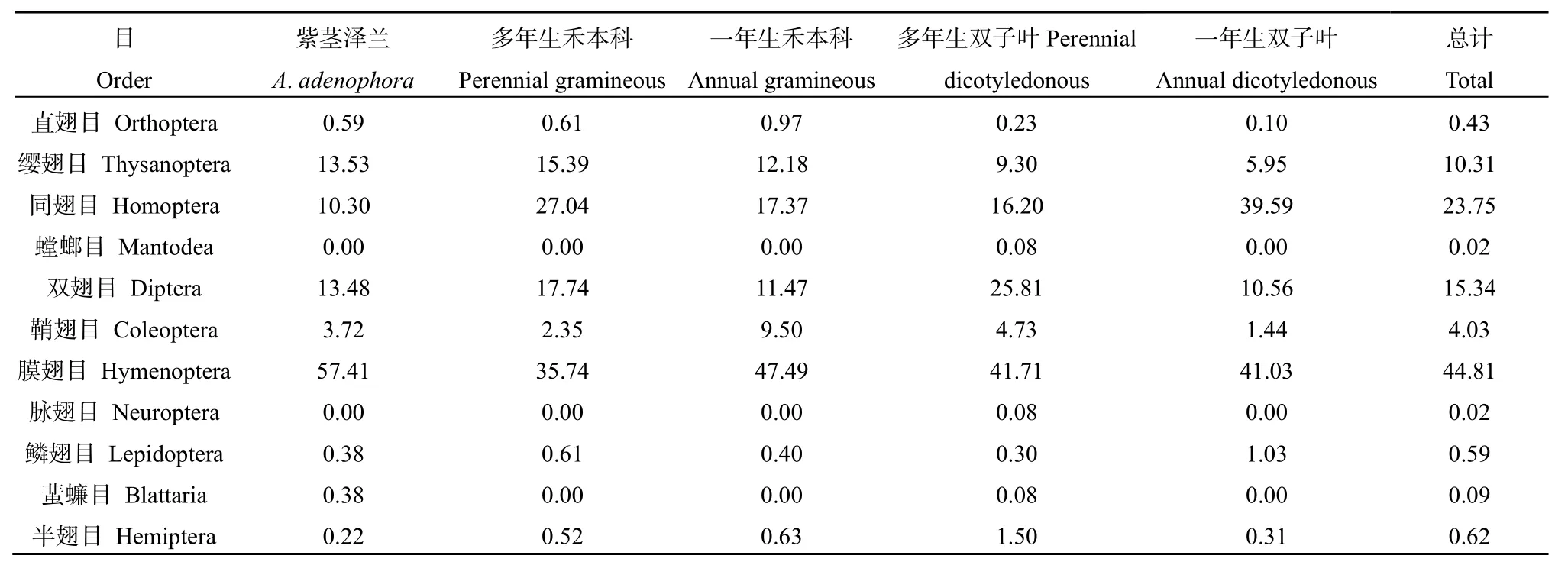
表2 紫茎泽兰群落与不同功能型植物群落昆虫群落组成Table2 Compositions of insect communities in A. adenophora community and different function plant communities
由表1可知,紫茎泽兰群落中采集到的昆虫物种数最少,但紫茎泽兰群落中采集到的昆虫个体数量比禾本科群落中的多,比双子叶群落中的少。由表2可知,紫茎泽兰群落中的同翅目、半翅目昆虫所占比例较其他不同功能型植物群落中的低;膜翅目昆虫所占比例较其他不同功能型植物群落中的高;直翅目、鳞翅目昆虫所占比例比禾本科植物群落中的低,但比双子叶植物群落中的高。螳螂目和脉翅目昆虫仅在多年生双子叶群落中有采集到。
2.1.2 紫茎泽兰入侵对昆虫群落组成的影响
在紫茎泽兰不同入侵程度的植物群落中,共获得昆虫标本5595个,隶属于10目52科138种。优势类群分别为膜翅目(61.11%)、同翅目(12.67%)、双翅目(11.76%)、缨翅目(11.17%)昆虫,这4个目累计占总数的96.71%。此外,还有少量的鞘翅目、直翅目、脉翅目、鳞翅目、蜚蠊目、半翅目昆虫(表4)。随紫茎泽兰入侵程度的增加,群落中昆虫的种类先减少后逐渐回升,而采集到的昆虫数量则先增加后减少(表3)。随紫茎泽兰的入侵程度的增加,不同群落中直翅目、缨翅目、同翅目、双翅目、鞘翅目、鳞翅目昆虫在群落中所占的比例呈先降低再升高的趋势;膜翅目、半翅目昆虫在群落中所占比例呈先升高再降低的趋势;蜚蠊目昆虫在未入侵群落中未采集到,随入侵程度的增加,其在群落中的比例也逐渐升高;脉翅目昆虫仅在轻度入侵群落中采集到(表4)。
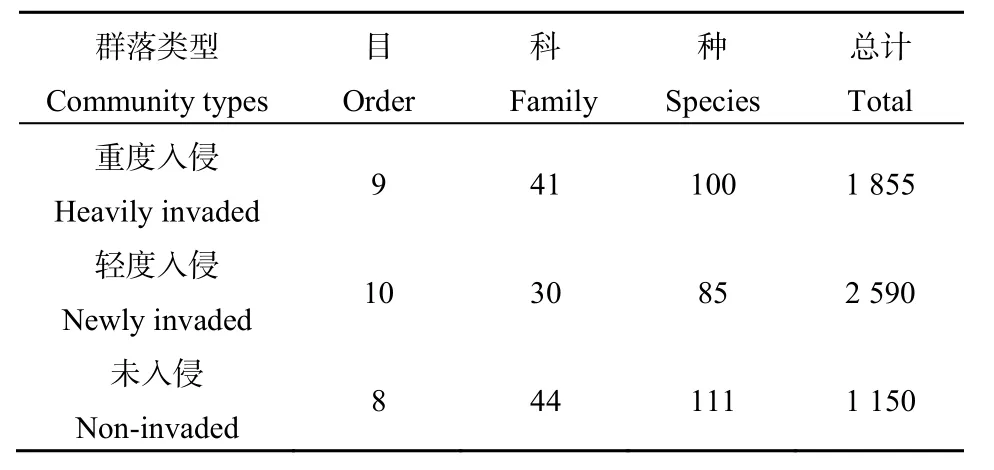
表3 紫茎泽兰入侵对昆虫组成的影响Table3 Effect of insect compositions with the invasion of A. adenophora
膜翅目中,主要优势种有东方行军蚁(D.orientalis)、长足切叶蚁(A. longipes)、广大腿小蜂(B. obscurata)、姬蜂科的食蚜蝇姬蜂(D.laetatorius)等;同翅目中,主要优势种有小绿叶蝉(E. flavescens)、大青叶蝉(T. viridis)、棉蚜(A.gossypii)等;双翅目中,主要优势种有食蚜蝇(D.fasciata)、大灰食蚜蝇(S. corollae)等;缨翅目中,主要优势种有食蚜蝇(D. fasciata)、大灰食蚜蝇(S.corollae)等;缨翅目优势种有花蓟马(F. intonsa)和八节黄蓟马(T. flavidalus)。
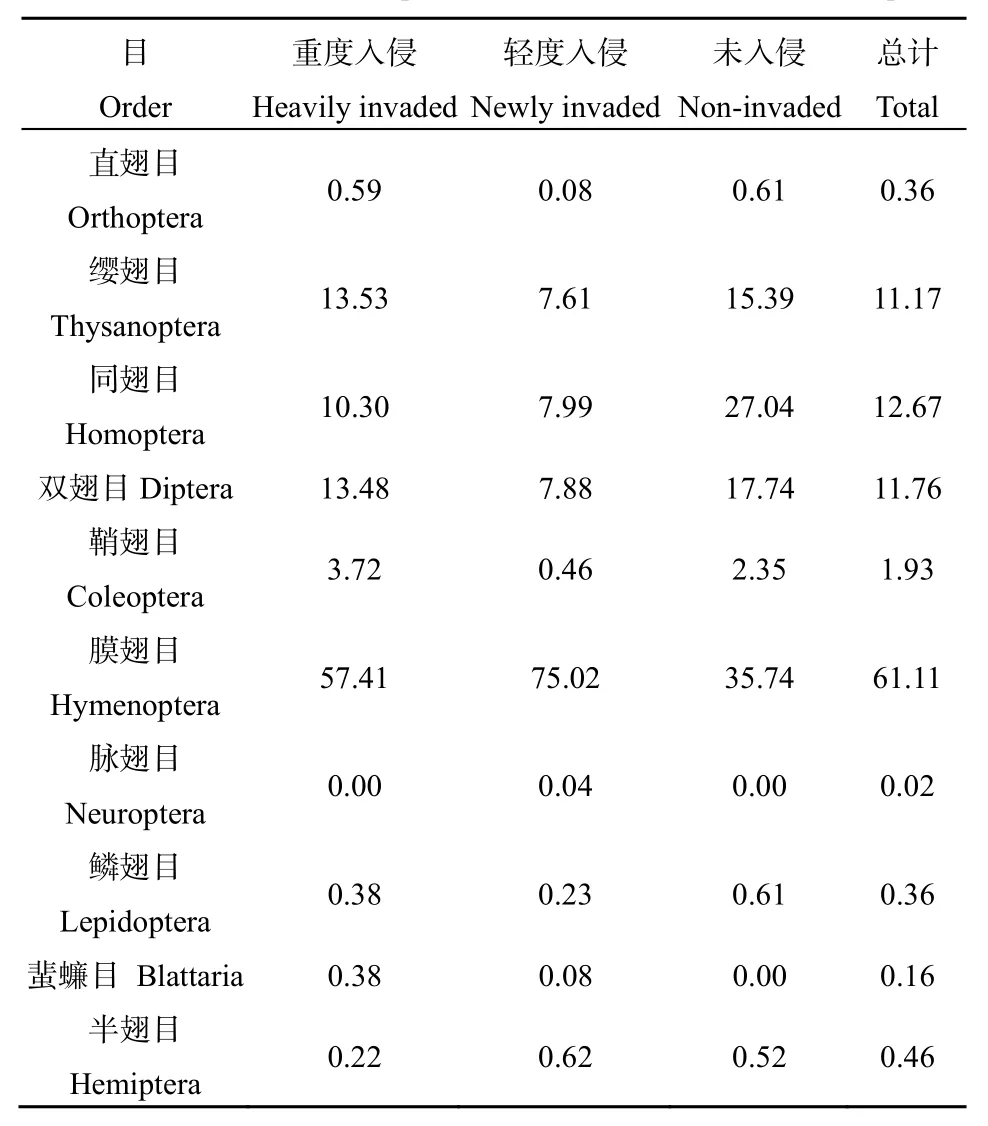
表4 紫茎泽兰入侵对昆虫群落组成的影响Table4 Effect of insect compositions with the invasion of A. adenophora
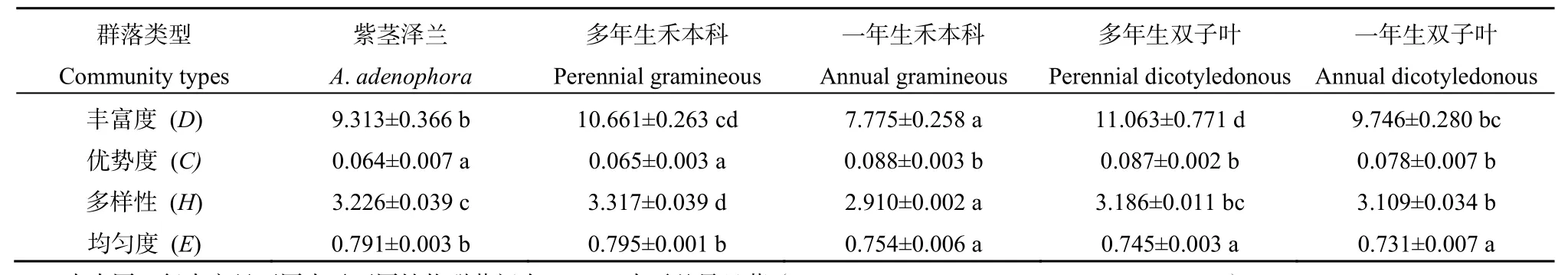
表5 紫茎泽兰群落与不同功能型植物群落昆虫多样性分析Table5 The diversity analysis of insect in A. adenophora community and different function plant communities
2.2 不同植物群落中的昆虫多样性分析
2.2.1 紫茎泽兰群落与不同功能型植物群落中的昆虫多样性分析
如表5所示,紫茎泽兰群落与不同功能型植物群落间的昆虫多样性指数具有一定的差异。紫茎泽兰与不同功能型植物群落间的昆虫丰富度指数(D)差异显著(F=13.636,P=0.000),紫茎泽兰群落中D显著高于一年生禾本科群落(P=0.003),但均显著低于多年生禾本科群落(P=0.019)和多年生双子叶群落(P=0.004),而与一年生双子叶无显著差异(P=0.423)。紫茎泽兰与不同功能型植物群落间的昆虫优势度指数(C)差异显著(F=8.563,P=0.000),紫茎泽兰群落中C与多年生禾本科群落无显著差异(P=0.806),但显著低于其他群落(P<0.05),而一年生禾本科、多年生双子叶和一年生双子叶群落中之间均无显著差异(P>0.05)。紫茎泽兰与不同功能型植物群落间的昆虫多样性指数(H)差异显著(F=26.101,P=0.000),紫茎泽兰群落中 H与多年生双子叶群落无显著差异(P=0.356),但均显著高于一年生禾本科群落和一年生双子叶群落(P<0.05),而显著低于多年生禾本科群落(P=0.046)。紫茎泽兰与不同功能型植物群落间的昆虫均匀度指数(E)差异显著(F=12.286,P=0.000),紫茎泽兰群落中E与多年生禾本科群落无显著差异(P=0.237),但均显著高于其他群落(P<0.05),而一年生禾本科、多年生双子叶和一年生双子叶群落之间均无显著差异(P>0.05)。
2.2.2 紫茎泽兰入侵对昆虫群落多样性的影响
如表6所示,紫茎泽兰入侵对昆虫多样性的影响如表6所示。紫茎泽兰不同入侵程度植物群落中昆虫丰富度指数(D)差异极为显著(F=121.966,P=0.000),随紫茎泽兰入侵程度的加重呈现先降低后升高的趋势;紫茎泽兰不同入侵程度植物群落中昆虫优势度指数(C)差异极为显著(F=125.300,P=0.000),随紫茎泽兰入侵程度的加重呈现先升高后降低的趋势,其轻度入侵群落中C显著高于未入侵群落和重度入侵群落(P<0.05),而重度入侵群落和未入侵群落间无显著差异(P=0.901);紫茎泽兰不同入侵程度植物群落中昆虫多样性指数(H)差异极为显著(F=344.987,P=0.000),随紫茎泽兰入侵程度的加重呈现先降低后升高的趋势,其轻度入侵群落中的 H显著低于未入侵群落和重度入侵群落(P<0.05),重度入侵群落显著低于未入侵群落(P=0.047);紫茎泽兰不同入侵程度植物群落中昆虫均匀度指数(E)差异极为显著(F=126.114,P=0.000),随紫茎泽兰入侵程度的加重呈现先降低后升高的趋势,其轻度入侵群落E显著低于未入侵群落和重度入侵群落(P<0.05),而重度入侵群落和未入侵群落之间无显著差异(P=0.828)。
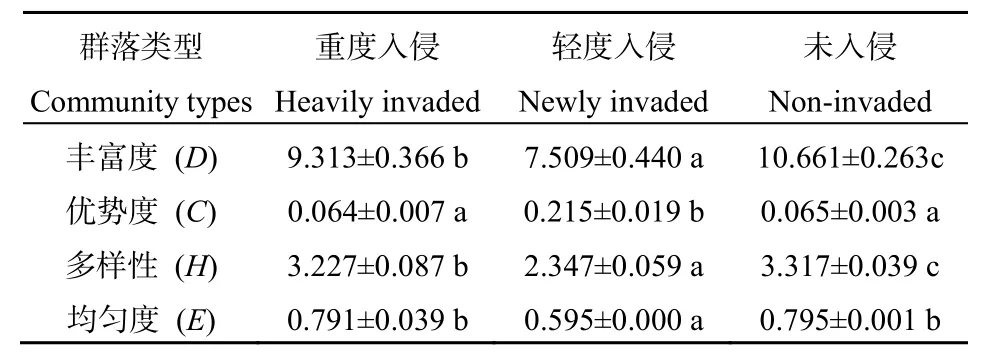
表6 紫茎泽兰入侵对群落昆虫多样性的影响Table6 Effect of insect diversity with the invasion of A. adenophora
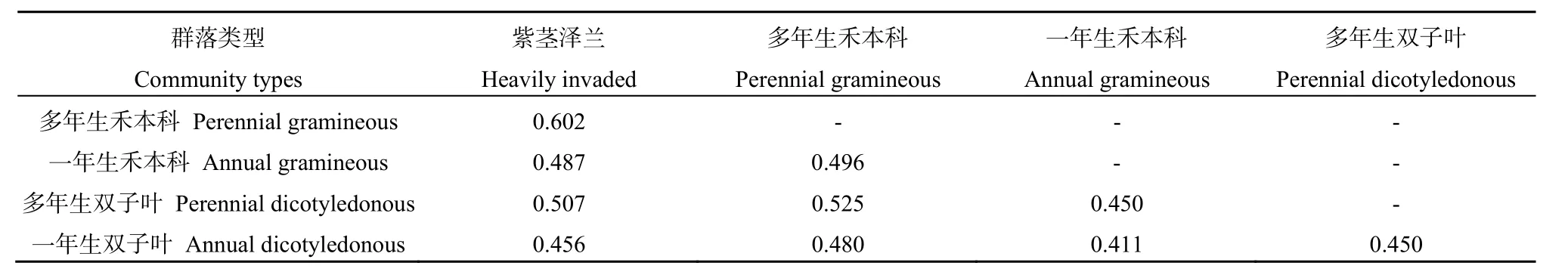
表7 紫茎泽兰群落与不同功能型植物群落中的昆虫群落相似性分析Table7 Similarity analysis of insect communities in A. adenophora community and different function plant communities

表8 紫茎泽兰入侵对群落中的昆虫群落相似性的影响Table8 Effect of similarity analysis of insect communities with the invasion of A. adenophora
2.3 不同植物群落中的昆虫群落相似性分析
2.3.1 紫茎泽兰群落与不同功能型植物群落中的昆虫群落相似性分析
对不同植物群落中昆虫群落组成的相似性进行分析(表7),结果表明,不同植物群落间的昆虫群落既有一定的相关性,有保持一定的独立性,群落相似性指数在 0.4~0.7间变化。紫茎泽兰群落中的昆虫群落与多年生植物群落相似,与多年生禾本科群落的相似性最高(0.602);与一年生植物群落不相似。多年生植物群落间的昆虫群落相似,一年生植物群落间的昆虫群落不相似,且与其他群落均不相似。
2.3.2 紫茎泽兰不同入侵程度群落中昆虫群落的相似性分析
对紫茎泽兰不同入侵程度的各群落中昆虫群落组成的相似性进行分析(表8),结果表明,不同入侵程度群落中的昆虫群落相似。随入侵程度的增加,群落中昆虫的相似性也增加。未入侵群落与轻度入侵群落相似,但相似性指数较低;随紫茎泽兰入侵程度的加重,轻度入侵群落与重度入侵群落的相似性指数升高。随紫茎泽兰入侵时间的增长,紫茎泽兰重度入侵群落中的昆虫群落逐渐恢复,与未入侵群落有较高的相似性,相似性指数达0.602。
3 讨论
地表昆虫群落与地上植物群落间存在动态的相互影响,植物群落的变化能对其昆虫群落结构产生重要影响,而昆虫群落间的相互作用又能对植物群落产生正负反馈作用(刘宁等,2014)。研究表明,植物及其凋落物可为地表昆虫提供丰富的食物来源和栖息场所,从而影响植物群落的昆虫群落结构(许湘琴等,2011)。有研究发现,入侵植物可通过改变原有食物网结构来影响生态系统的功能,引起营养链结构网络和周围环境变化,或通过非营养作用改变各级消费者的行为活动和种群密度,从而影响其昆虫群落多样性(王思凯等,2013)。入侵植物可通过与本地植物竞争营养、光照从而影响本地植物生长发育,竞争替代本地寄主植物或通过影响昆虫行为,而使这些植物群落的昆虫多样性发生变化(Lau et al.,2005;Bais et al.,2006;Russell et al.,2007)。有研究发现,一些入侵植物在新的栖息地建立种群,逃脱了本地共同进化的天敌;而入侵植物可作为本地植食性昆虫的资源,驱动本地昆虫的选择和形态变化(Strauss et al.,2006;Carroll et al.,2007)。牛燕芬等(2015)研究发现,随着入侵植物的入侵时间延长和入侵程度加重,入侵群落结构逐步趋于稳定,本地的昆虫亦与之协同进化,本地天敌开始从行为、生理生化等方面适应入侵植物,入侵植物天敌有逐渐增加的趋势。有研究认为,入侵植物可能是一种潜在高度合适的寄主(Harvey et al.,2010;Herrera et al.,2011),然而一些研究认为入侵植物可毒害本地植食性昆虫(Ding et al.,2009;Tallamy et al.,2010);一些植食性昆虫更喜欢本地植物(White et al.,2008;Brown et al.,2009),或更喜欢入侵植物(Fielding et al.,2011)。且邻近植物的存在会影响昆虫对寄主植物的选择,入侵植物可吸引植食性昆虫,增加植食性昆虫对本地植物的取食,或降低植食性昆虫对本地植物的取食(Bezemer et al.,2014)。有研究者对黄顶菊入侵的各生境进行调查,发现各生境均以膜翅目的蚁科昆虫为优势类群,各类昆虫在不同生境中的分布情况存在着较大的差异(刘宁,2013)。本研究对不同植物群落中昆虫群落的比较发现,各群落以膜翅目、同翅目、双翅目、缨翅目的昆虫数量居多;紫茎泽兰入侵群落中缨翅目昆虫所占比例均高于同翅目昆虫,而在未入侵的群落中缨翅目昆虫所占比例均低于同翅目,这可能是由于紫茎泽兰某化学成分对同翅目昆虫有毒害或趋避作用,亦或是对缨翅目昆虫有引诱作用导致的;此外,本研究调查发现双子叶植物群落中昆虫种类和数量多于禾本科植物群落,这与相关研究结果类似,双子叶植物群落的地上生物量较大、郁闭度较高、均匀度较小,可提供更多的异质性栖息地,容纳更多的昆虫种类和数量(Denno et al.,2004)。
入侵植物可通过与本地植物竞争或替代本地寄主植物而对本地昆虫情况产生重要影响(Wagner et al.,2010)。入侵植物可降低本地昆虫的多样性和丰富度,如在智利,被外来植物入侵的森林中节肢动物的丰富度和物种丰富度均比未入侵森林中的低(Hagen et al.,2010);而一些研究表明,当去除入侵植物后本地昆虫的丰富度和多样性均会增加(Hanula et al.,2011),甚至全部恢复(Gratton et al.,2006)。本研究结果发现,未入侵的不同植物群落经过长期的竞争演替,其中的生物群落已趋于相对稳定,而紫茎泽兰的入侵打破了群落的结构,使昆虫的各多样性发生了较大变化;紫茎泽兰轻度入侵群落的昆虫多样性指数 D、H、E显著低于重度入侵群落,且显著低于未入侵的其他植物群落;而多年生植物植物群落中昆虫群落相对稳定,紫茎泽兰作为一种多年生双子叶植物,其入侵群落的昆虫多样性指数与多年生植物群落间无显著差异,且均大于一年生植物群落。
本研究对不同植物群落的昆虫群落相似性分析表明,紫茎泽兰入侵群落间的昆虫群落多样性相似性较高,这可能是不同入侵程度紫茎泽兰所产生的相同挥发性物质、化感物质等对本地昆虫诱集或毒害作用相近有关,具体深入的影响关系有待进一步研究证实。相关专家对类似相关性研究发现,入侵群落与未入侵多年生禾本科植物群落中的昆虫群落有较高的相似性,而对未入侵多年生禾本科植物如百喜草(Paspalum natatu)(龙忠富等,2011)、黑麦草(Lolium perenne)(于亮等,2009)、宽叶雀稗(Paspalum wetsfeteini)(卢向阳,2014)、非洲狗尾草(Setaria sphacelata)(蒋智林等,2014;王五云等,2014)、皇竹草(Pennisetum purpureum)、臂形草(Brachiaria ramosa)等(彭恒等,2010;卢向阳等,2012;华劲松,2013)的研究发现,这些植物对紫茎泽兰均具有很强的入侵抵御作用,这说明与紫茎泽兰单优群落昆虫多样性相似性高的特征可能是对紫茎泽兰入侵具有较强抵御作用的指示因子之一。
4 结论
紫茎泽兰不同入侵程度和未入侵不同功能型植物群落样地共采集到昆虫标本12099个,隶属于11目64科168种,优势类群为膜翅目、同翅目、双翅目、缨翅目昆虫,占总数的94.86%。随紫茎泽兰的入侵,群落中直翅目、缨翅目、同翅目、双翅目、鞘翅目、鳞翅目昆虫在群落中所占的比例呈先降低再升高的趋势;膜翅目、半翅目昆虫在群落中所占比例呈先升高再降低的趋势。紫茎泽兰群落中的同翅目、半翅目昆虫所占比例较不同功能型本地植物群落低,而膜翅目昆虫所占比例较其他群落高。随紫茎泽兰入侵程度的加重,昆虫多样性指数D、H、E呈先降低后升高的趋势;昆虫优势度指数C呈先升高再降低的趋势。多年生禾本科群落的各昆虫多样性指数均大于紫茎泽兰群落,与紫茎泽兰群落间的差异最小,且有较高的相似性。紫茎泽兰的入侵对入侵生境植物群落的昆虫多样性会产生一定程度的影响,且在这个过程中随着入侵影响的加重,其植物群落趋于稳定,入侵生境植物群落昆虫多样性会发生变化,建立与新的植物群落相适应的昆虫群落,不断丰富而逐于稳定;另一方面,植物昆虫群落的特征及其稳定性也可能在一定程度上反应出对紫茎泽兰入侵的抵御性能。
BAIS H P, WEIR T L, PERRY L G, et al. 2006. The role of root exudates in rhizosphere interactions with plants and other organisms [J]. Annual Review of Plant Biology, 57: 233-266.
BEZEMER T M, HARVEY J A, CRONIN J T. 2014. Response of native insect communities to invasive plants [J]. Annual Review of Entomology, 59: 119-141.
BROWN B J, MITCHELL R J. 2001. Competition for pollination: effects of pollen of an invasive plant on seed set of a native congener [J].Oecologia, 129(1): 43-49.
BROWN W P, ZUEFLE M E. 2009. Does the periodical cicada, Magicicada septendecim, prefer to oviposit on native or exotic plant species? [J].Ecological Entomology, 34(3): 346-355.
CARROLL S P, FOX C W. 2007. Dissecting the evolutionary impacts of plant invasions: bugs and beetles as native guides [J]. Global Change Biology, 13: 1644-1657.
DENNO R F, MITTER M S, LANGELLOTTO G A et al. 2004. Interactions between a hunting spider and a web-builder: consequences of intraguild predation and cannibalism for prey suppression [J].Ecological Entomology, 29(5): 566-577.
DING J Q, BLOSSEY B. 2009. Differences in preference and performance of the water lily leaf beetle, Galerucella nymphaeae, populations on native and introduced aquatic plants [J]. Environmental Entomology,38(6): 1653-1660.
FIELDING D J, CONN J S. 2011. Feeding preference for and impact on an invasive weed (Crepis tectorum) by a native, generalist insect herbivore, Melanoplus borealis (Orthoptera: Acrididae) [J]. Annals of the Entomological Society of America, 104(6): 1303-1308.
GRATTON C, DENNO R F. 2006. Arthropod food web restoration following removal of an invasive wetland plant [J]. Ecological Applications, 16(2): 622-631.
HAGEN E N, BAKKER J D, GARA R I. 2010. Aerial arthropod communities of native and invaded forests, Robinson Crusoe Island,Chile [J]. Ecological Entomology, 39(4): 1159-1164.
HANULA J L, HORN S. 2011. Removing an invasive shrub (Chinese privet) increases native bee diversity and abundance in riparian forests of the southeastern United States [J]. Insect Conservation and Diversity, 4(4): 275-283.
HARVEY J A, BIERE A, FORTUNA T, et al. 2010. Ecological fits, mis-fits and lotteries involving insect herbivores on the invasive plant, Bunias orientalis [J]. Biological Invasions, 12(9): 3045-3059.
HERRERA A M, CARRUTHERS R I, MILLS N J. 2011. No evidence for increased performance of a specialist psyllid on invasive French broom[J]. Acta Oecologica, 37(2): 79-86.
LAU J A, STRAUSS S Y. 2005. Insect herbivores drive important indirect effects of exotic plants on native communities [J]. Ecology, 86:2990-2997.
PIELOU E C. 1975. Ecological diversity [M]. New York: John Wiley.
RUSSELL F L, LOUDA S M, RAND T A et .al. 2007. Variation in herbivore-mediated indirect effects of an invasive plant on a native plant [J]. Ecology, 88(2): 413-423.
SIMPSON E. H. 1949. Measurement of diversity [J]. Nature, 163: 688.
STRAUSS S Y, LAU J A, CARROLL S P. 2006. Evolutionary responses of natives to introduced species: What do introductions tell us about natural communities? [J]. Ecology Letters, 9(3): 354-371.
TALLAMY D W, BALLARD M, D’AMICO V. 2010. Can alien plants support generalist insect herbivores? [J]. Biological Invasions, 12(7):2285-2292.
WAGNER D L, VAN DRIESCHE R G. 2010. Threats posed to rare or endangered insects by invasions of nonnative species [J]. Annual Review of Entomology, 55: 547-568.
WHITE E M, SIMS N M, CLARKE A R. 2008. Test of the enemy release hypothesis: the native magpie moth prefers a native fireweed (Senecio pinnatifolius) to its introduced congener (S. madagascariensis) [J].Austral Ecology, 33(1): 110-116
WU Y T, WANG C H, ZHANG X D, et al. 2009. Effects of saltmarsh invasion by Spartina alterniflora on arthropod community structure and diets [J]. Biological Invasions, 11(3): 635-649.
邓丹丹, 刘棋, 蒋智林, 等. 2015. 紫茎泽兰与不同植物群落土壤养分及酶活性差异[J]. 生态环境学报, 24(9): 1466-1471.
杜喜翠, 谭万忠, 孙现超. 2011. 外来入侵植物黄顶菊上昆虫种类多样性研究[J]. 西南大学学报(自然科学版), 33(6): 1-6.
高贤明, 桑卫国. 2003. 紫茎泽兰入侵现状、特征、趋势及防治对策[A]//中国植物学会. 中国植物学会七十周年年会论文摘要汇编(1933—2003).
华劲松. 2013. 几种优质牧草替代种植对紫茎泽兰的防控效果分析[J].湖北农业科学, 52(10): 2368-2370, 2376.
蒋智林, 王五云, 雷桂生, 等. 2014. 紫茎泽兰与4种功能型草本植物根系生长特征和竞争效应[J]. 应用生态学报, 25(10): 2833-2839.
刘伦辉, 谢寿昌, 张建华. 1985. 紫茎泽兰在我国的分布危害、与防除途径的探讨[J]. 生态学报, 5(1): 1- 6.
刘宁, 付卫东, 张国良, 等. 2014. 黄顶菊入侵对不同生境地表土壤动物群落的影响[J]. 生态学杂志, 33(1): 176 - 183.
刘宁. 2013. 黄顶菊入侵对不同生境土壤动物群落的影响[D]. 泰安: 山东农业大学.
刘志磊, 徐海根, 丁晖. 2006. 外来入侵植物紫茎泽兰对昆明地区土壤动物群落的影响[J]. 生态与农村环境学报, 22(2): 31-35.
龙忠富, 刘华荣, 孟军江, 等. 2011. 百喜草对紫茎泽兰的生物替代控制作用[J]. 贵州农业科学, 39(8): 212-215.
卢向阳, 张锦华, 左相兵, 等. 2012. 几种替代植物对入侵杂草紫茎泽兰的防控效果[J]. 贵州农业科学, 40(6): 103-106, 109.
卢向阳. 2014. 替代植物的选育与紫茎泽兰的持续控制[J]. 杂草科学,32(1): 69-74.
牛红榜. 2007. 外来植物紫茎泽兰入侵的土壤微生物学机制[D]. 北京:中国农业科学院.
牛燕芬, 莫丽玲, 张永福, 等. 2015. 云南本地昆虫取食紫茎泽兰的原因研究[J]. 云南农业大学学报(自然科学), 30(4): 489-493.
彭恒, 曹志勇, 桂富荣, 等. 2013. 混种非洲狗尾草对紫茎泽兰表型特征的影响[J]. 云南农业大学学报(自然科学版), 25(3): 303-308.
亓东明. 2014. 紫茎泽兰入侵地不同林分类型地表节肢动物的群落多样性[J]. 江苏农业科学, 42(8): 375-377.
秦浩, 董刚, 张峰. 2015. 山西植物功能型划分及其空间格局. 生态学报, 35(2): 396-408.
王思凯, 盛强, 储忝江, 等. 2013. 植物入侵对食物网的影响及其途径[J]. 生物多样性, 21(3): 249-259.
王文琪, 王进军, 赵志模, 等. 2009. 不同生境紫茎泽兰对生态群落中节肢动物多样性的影响[J]. 西南大学学报, 31(12): 14-20.
许湘琴, 林植华, 陈慧丽. 2011. 凋落物分解对土壤生物的影响[J]. 生态学杂志, 30(6): 1258-1264.
尹文英, 胡圣豪, 王敏. 1998. 中国土壤动物检索图鉴[M]. 北京: 科学出版社.
于亮, 李世吉, 桂富荣, 等. 2009. 黑麦草和紫花苜蓿对紫茎泽兰的竞争作用研究[J]. 云南农业大学学报, 24(2): 164-168.
于文清, 刘万学, 桂富荣, 等. 2012. 外来植物紫茎泽兰入侵对土壤理化性质及丛枝菌根真菌 (AMF) 群落的影响[J]. 生态学报, 32(22):7027-7035.
张海周, 王正军, 张向欣. 2009. 野鸭湖湿地自然保护区地表昆虫多样性分析[J]. 首都师范大学学报(自然科学版), 30(6): 31-34, 44.
张修玉, 许振成, 宋巍巍, 等. 2010. 紫茎泽兰 (Eupatorium adenophorum)入侵地的生物多样性[J]. 生态环境学报, 19(7): 1525-1531.
郑乐怡, 归鸿. 1999. 昆虫分类[M]. 南京: 南京师范大学出版社.
中国科学院中国动物志编辑委员会. 2003. 中国动物志. 昆虫纲[M]. 北京: 科学出版社.
Effect of Ageratina adenophora Invasion on Insect Diversity in Subtropical Mountains in SW China
JIANG Zhilin1,2*, DENG Dandan1,2*, LIU Wanxue3, WAN Fanghao3, LI Zhengyue2*
1. Puer University, Puer 665000, China;2. Key Laboratory for Agricultural Biodiversity and Pest Management Ministry of Education//Plant Protection College, Yunnan Agricultural University,Kunming 650201, China;3. The State Key Laboratory for Biology of Plant Diseases and Insect Pests//Institute of Plant Protection, Chinese Academy of Agricultural Sciences,Beijing 100094, China
To assess the effects of Ageratina adenophora invasion on insect community and understand the ecological mechanisms of local plant communities resist invasive plants, we adopted trap method, sweeping method and yellow plate method to investigate insect diversity in A. adenophora invaded and non-invaded plant communities. Totally, there were 12 099 individuals of insect represent 168 species were collected from different plant communities, that belonging to 11 orders and 64 families. Our results indicated that the dominant insect (94.86% of the total) were Hymenoptera, Homoptera, Diptera, Thysanoptera. With the invasion of A. adenophora, the diversity of Orthoptera, Thysanoptera, Homoptera, Diptera, Coleoptera and Lepidoptera was decreased firstly and increased subsequently, whereas that of Hymenoptera and Hemiptera was increased firstly and decreased subsequently. The insect community diversity index of D, H and E were decreased firstly and increased subsequently. In contrast, the dominance index C was increased firstly and decreased subsequently. Our results suggest that the perennial Gramineae plants community have higher insect diversity than A. adenophora communitis. There may be some insects that eat or inhibit the growth of A. adenophora can be stronger to resist the invasion of A. adenophora in perennial gramineous plants community.
Ageratina adenophora; insect diversity; diversity index; insect community structure; different plant communities
10.16258/j.cnki.1674-5906.2017.12.002
X176
A
1674-5906(2017)12-2008-08
蒋智林, 邓丹丹, 刘万学, 万方浩, 李正跃. 2017. 紫茎泽兰入侵对不同植物群落类型中昆虫多样性的影响[J]. 生态环境学报, 26(12): 2008-2015.
JIANG Zhilin, DENG Dandan, LIU Wanxue, WAN Fanghao, LI Zhengyue. 2017. Effect of Ageratina adenophora invasion on insect diversity in subtropical mountains in SW China [J]. Ecology and Environmental Sciences, 26(12): 2008-2015.
国家自然科学基金项目(31360456;31060252);云南省中青年学术带头人后备人才项目(2014HB027);中国科学院“西部之光青年学者”项目(科发人函字[2016]84号);科技部重大专项“受威胁区域生物多样性恢复及示范”(2017YFC0505206);云南省自然基金项目(2011FB050);普洱学院生物安全与生物产业创新团队项目
蒋智林(1977年生),男,教授,研究方向为生物安全。E-mail: zhilin_jiang@126.com
*通信作者
2017-07-22
猜你喜欢
浙江农林大学学报(2022年6期)2022-12-07
热带农业工程(2022年3期)2022-08-16
云南化工(2021年7期)2021-12-21
湖南税务高等专科学校学报(2021年6期)2021-12-06
中成药(2019年12期)2020-01-04
中成药(2018年12期)2018-12-29
中成药(2018年6期)2018-07-11
湖南农业(2016年3期)2016-06-05
中国塑料(2015年12期)2015-10-16
