
围封对荒漠草原沙芦草种群构件生物量分配特性的影响
2017-12-29 07:00:38赵盼盼邵文山靳长青宋立肖龚诗佩李国旗
生态环境学报 2017年12期
赵盼盼,邵文山,靳长青,宋立肖,龚诗佩,李国旗*
1. 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川 750021;2. 宁夏大学西北退化生态系统恢复与重建教育部重点实验室,宁夏 银川 750021
围封对荒漠草原沙芦草种群构件生物量分配特性的影响
赵盼盼1,2,邵文山1,2,靳长青1,2,宋立肖1,2,龚诗佩1,2,李国旗1,2*
1. 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川 750021;2. 宁夏大学西北退化生态系统恢复与重建教育部重点实验室,宁夏 银川 750021
以宁夏盐池县四墩子荒漠草原沙芦草(Agropyron mongolicum)为研究对象,通过对比研究围栏内外沙芦草种群各构件生物量、构件分配格局及其相关关系,试图揭示围栏封育对植物资源分配格局和生态策略的影响。结果表明,(1)围栏封育后,沙芦草种群的高度、丛幅、盖度以及株丛生物量显著提高,提高比例依次为78.96%(P=0.040)、63.50%(P=0.013)、50.89%(P=0.032)、205.38%(P=0.022),而密度无显著变化(P=0.330)。(2)围封对沙芦草种群各构件生物量具有显著影响,与围栏外相比,围栏内沙芦草种群根、叶、穗生物量显著提高,提高比例依次为183.52%(P=0.020)、334.09%(P=0.011)、381.25%(P=0.005);且围栏内外均表现为根生物量最大(围栏内为38.53 g,围栏外为13.59 g),穗生物量最小(围栏内为6.16 g,围栏外为1.28 g)。(3)围封后,沙芦草种群的生物量分配比发生了不同程度的变化。其中,生殖枝和叶的生物量分配比显著升高,分别升高了31.45%(P=0.020)和4.20%(P=0.000),而营养枝生物量分配比和根冠比显著降低,降低比例分别为281.62%(P=0.002)和13.33%(P=0.023)。(4)除营养枝生物量与丛幅间表现为显著负相关关系外(P=0.039),沙芦草种群各构件生物量间以及各构件生物量与株高、丛幅之间均存在显著或极显著正相关性。围栏封育对沙芦草种群构件生物量及其分配格局有显著的影响,说明围封是植物维持其种群持续更新能力和恢复其在群落中地位的重要措施之一。
沙芦草;围封;种群构件;生物量分配
沙芦草(Agropyron mongolicum)又称蒙古冰草,为禾本科冰草属多年生草本植物,适应性较高,具有较强的抗旱、抗寒、耐风沙等特性(赵有璋,2004),常见于干草原和荒漠草原的典型沙质环境中,在荒漠化草原中多以伴生种出现(云锦凤等,1989)。有关沙芦草的研究表明(安渊等,1997;刘文清等,2003),沙芦草与其他种混播可以有效控制流沙移动,显著改善生态环境。围封作为一种促进退化草原尽快恢复的重要手段,是通过解除牲畜的践踏、采食及粪便等外界干扰,使退化生态系统在自身的更新能力下进行恢复(杨勇等,2016;闫玉春等,2009)。围封在国内外已广泛开展,有关其在草地恢复及其保护机制机理方面的研究最为深入(金艳强等,2017)。
构件生物量分配是指植物生长发育繁殖过程中所同化的资源用于根、茎、叶片、花、果实等各器官的比例(肖遥等,2014),它体现了植物在不同的生境中的生态策略(Weiner,2004),是对环境长期适应的结果(Poorter et al.,2012;王杨等,2017)。作为一个功能性指标,植物构件生物量不仅反映了生态系统中的物质循环和能量流动,而且在生态系统结构的组成方面具有非常重要的意义(Bloom et al.,1985;罗永开等,2017;严月等,2017;Hovenden et al.,2014)。因此,对植物各构件进行生物量测定在生态系统研究中具有重要意义(王蕙等,2015)。近年来,国内外已对植物生物量分配进行了大量研究,金艳强等(2017)对元江干热河谷围封条件下稀树灌草丛植被的生物量分配进行了研究,结果表明,围封显著降低了草本植物的地下、地上生物量比例。王蕙等(2015)研究了围封对沙质天然草地植物的构件生物量分配的影响。Barton(2016)对美国夏威夷当地物种做了相关研究:动物的采食显著减少了枝生物量的分配。本文通过研究围封条件下沙芦草种群各构件生物量的变化,旨在揭示围栏封育后沙芦草在构件尺度上生物量的分配规律,进一步为荒漠草原区生态系统可持续利用提供一定参考。
1 材料与方法
1.1 试验区概况
试验区位于宁夏东部盐池县四墩子村(E106°30′10″~107°48′,N37°04′~38°10′),毛乌素沙地西南缘,为黄土高原向鄂尔多斯台地的过度地带,自然条件较为恶劣,属于典型的中温带大陆性季风气候,年均气温为7.7 ℃,年降水量为250~350 mm,大部分集中在6—9月,年蒸发量在2500 mm左右,年无霜期 165 d。地貌为缓坡丘陵,土壤类型主要为灰钙土、风沙土、黄绵土,土壤质地为沙壤和粉沙壤(杨刚等,2003)。试验区位于中国温带草原的过渡地带,属于欧亚草原区,群落表现出旱生植物与典型草原建群种多年生禾草的镶嵌式分布格局。研究区域植被优势种主要有沙芦草(A.mongolicum)、柠条锦鸡儿(Caragana korshinskii)、猪毛蒿(Artemisia scoparia)、牛枝子(Lespedeza potaninii)、老瓜头(Cynanchum komarovii)、针茅(Stipaca pillata)等。
1.2 研究方法
1.2.1 取样
2017年8月初,沙芦草种子成熟时期,在具有相同气候条件和地理位置的围封16年(2001年开始围封,总盖度为 65%~75%)与未围封(总盖度为 40%~50%)样地内沿着对角线随机取 3个面积为100 m2(10 m×10 m)的大样方,在大样方的4个角各设1个4 m×4 m的中样方,再在中样方中按梅花形取5个1 m×1 m的小样方,每个样地60个小样方,总共120个小样方,调查记录每个小样方内沙芦草的高度、盖度、丛幅和密度。之后随机选取 30丛沙芦草,采取全挖法,尽量保证获取其整丛生物量。所采集的样品带回实验室后,将地上部分按营养枝和生殖枝分开;再将营养枝和生殖枝按茎、叶和茎、叶、穗等构件分开。地下部分去除杂草和死根,冲洗干净。将各构件分别装入纸袋,75 ℃烘干至恒重。
1.2.2 数据分析
运用 Microsoft Excel 2010进行数据整理及图表制作;运用SPSS 17.0进行数据处理:采用方差分析中的配对 T检验法进行差异显著性分析(α=0.05),采用Pearson相关系数进行相关性分析。
2 结果与分析
2.1 围封对沙芦草种群特征的影响
由表1可知,围栏封育后,沙芦草种群的高度、丛幅、盖度、总生物量均表现为显著增加趋势,增加比例依次为 78.96%(P=0.040)、63.50%(P=0.013)、50.89%(P=0.032)、205.38%(P=0.022);而其密度在数值上虽有微弱的增加,但没有达到显著水平(P=0.330)。
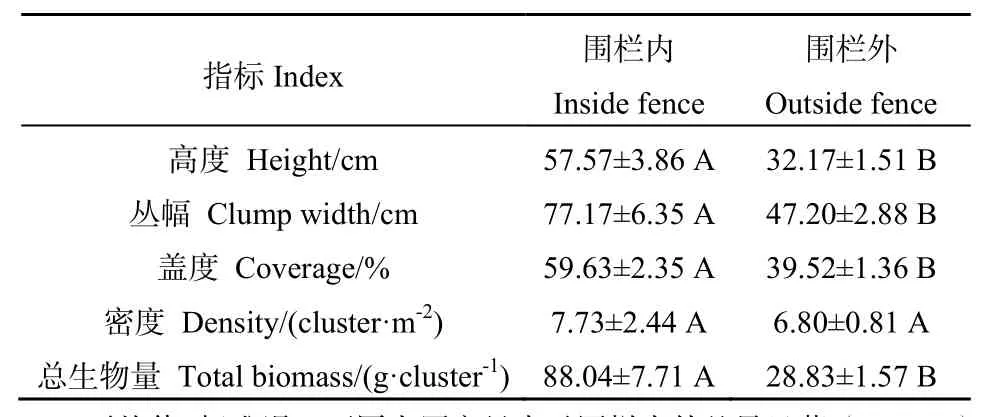
表1 围栏内外沙芦草的种群特征Table1 Population characteristics inside and outside the fence of A. mongolicum
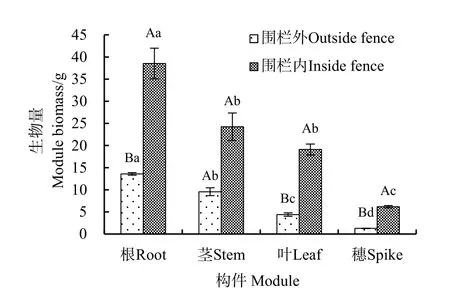
图1 围栏内外沙芦草种群各构件的生物量Fig.1 Module biomass of A. mongolicum population inside and outside the fence
2.2 围封对沙芦草种群各构件生物量的影响
图1所示为围栏内外沙芦草种群各构件的生物量变化情况。由图可知,围栏内外均表现为根生物量最大(围栏内为38.53 g,围栏外为13.59 g),穗生物量最小(围栏内为6.16 g,围栏外为1.28 g)。围栏内除茎、叶生物量间无显著差异外,其他构件生物量间均有显著差异(P=0.021);围栏外根、茎、叶、穗间均有显著差异(P=0.000)。围栏封育后,沙芦草的根、叶、穗生物量均显著提高,提高比例依次为183.52%(P=0.020)、334.09%(P=0.011)、381.25%(P=0.005);茎生物量虽有所提高,但未达到显著性水平(P=0.065)。
2.3 围封对沙芦草种群各构件生物量分配比的影响
由图2可知,围栏内外均表现为根生物量分配比最大(围栏内为43.74%,围栏外为47.44%),穗生物量分配比最小(围栏内为 7.09%,围栏外为4.44%),根、茎、叶、穗之间均存在显著性差异(P<0.05);另外,围栏内外生殖枝生物量分配比(围栏内为 53.17%,围栏外为40.42%)均大于营养枝的(围栏内为 3.21%,围栏外为 12.25%)。围栏封育后,沙芦草种群生物量分配发生不同程度的变化。其中,生殖枝生物量分配比和叶生物量分配比显著升高,分别升高 31.45%(P=0.020)和 4.20%(P=0.000),营养枝生物量分配比降低 281.62%(P=0.002),根、茎分配比有所降低,但差异不显著(P>0.05)。
2.4 围封对沙芦草种群根冠比的影响
围栏内外沙芦草种群的根冠比如表2所示。由表可知,沙芦草种群的根冠比在围栏内外分别为0.78和0.90,围栏封育后,其种群根冠比显著降低(P<0.05),降低比例为13.33%。
2.5 沙芦草种群株高、丛幅及其各构件生物量间的相关性
表3显示的是沙芦草种群株高、丛幅及其各构件生物量间的相关关系。由表可知,沙芦草种群营养枝生物量与其株高、丛幅、根、茎、叶、穗、生殖枝和全株生物量间均表现为不同程度负相关关系,且与丛幅间达到了显著负相关关系,相关系数r=-0.834(P<0.05)。株高、丛幅、根、茎、叶、穗、生殖枝及全株生物量之间均表现为显著或极显著的正相关关系,且相关系数均大于 0.875,其中以生殖枝生物量与全株生物量的相关系数最大(0.998)。

表2 围栏内外沙芦草种群的根冠比Table2 The root/shoot ratio of A. mongolicum population inside and outside the fence
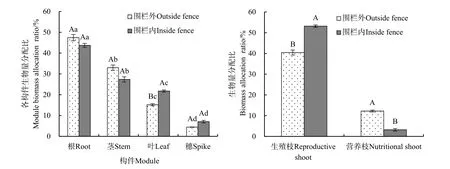
图2 围栏内外沙芦草种群各构件的生物量分配比Fig.2 Module biomass allocation of A. mongolicum population inside and outside the fence
3 讨论
围封解除了家畜的践踏、采食等干扰,使得草原生态系统在自然条件下向着一定方向演替(杨勇等,2016)。本研究中,围栏封育后,沙芦草种群的高度、丛幅、盖度及总生物量显著提高,而密度的变化不显著。这表明,围栏封育后,外界干扰的减小和生境的改善有利于植被的恢复和生产力的提高。一方面,作为优势种的沙芦草,其本身就是一种优良牧草,围栏封育使其避免了家畜的啃食和践踏,从而有效提高了沙芦草种群的高度、丛幅、盖度和生物量;另一方面,围栏封育后土壤养分积累、营养元素供给增加以及土壤物理性质的改善,都有助于植物的生长繁殖(刘忠宽等,2006;Hosseinzadeh et al.,2010)。这与前人(金艳强等,2017;郑伟等,2013;Lamas et al.,2013)的研究结果相似。沙芦草种群密度无显著变化,主要原因是,禾本科植物沙芦草以无性繁殖(分蘖)为主,种子繁殖为辅;另外围栏内沙芦草种群密度还可能与其种内的密度效应有关,而围栏外家畜的啃食、土壤水肥等都是影响其有性繁殖的因素。
生物量是反映植物与环境相互作用的重要指标,是植物对环境适应能力及生长发育规律的体现,其中个体及构件生物量又是生态系统获取能量的能力体现(宇万太等,2001)。在天然围封草地生态系统中,植被通过自身多样性和复杂性的调配对异质性资源做出响应,进而有效提高整个生态系统对有限资源的利用效率(Kroons et al.,1995;Pan et al.,2008;Wang et al.,2006)。本研究中,围栏内外均表现为根生物量显著大于其他构件生物量,围封显著提高了根、叶、穗的生物量。首先是因为在干旱的荒漠草原,植物为了获得生长繁殖所需要的水分和无机营养,加强根系的生长是最有效的策略之一(周兵等,2015)。其次,可能与其自身所在的生境有关,围封后,良好的生境大大提高了沙芦草的有效分蘖数,以致根、叶、穗的生物量显著增加。最后,围栏内根生物量显著增加,也可能是为了满足围封后沙芦草自身更多的营养和水分需求造成的。
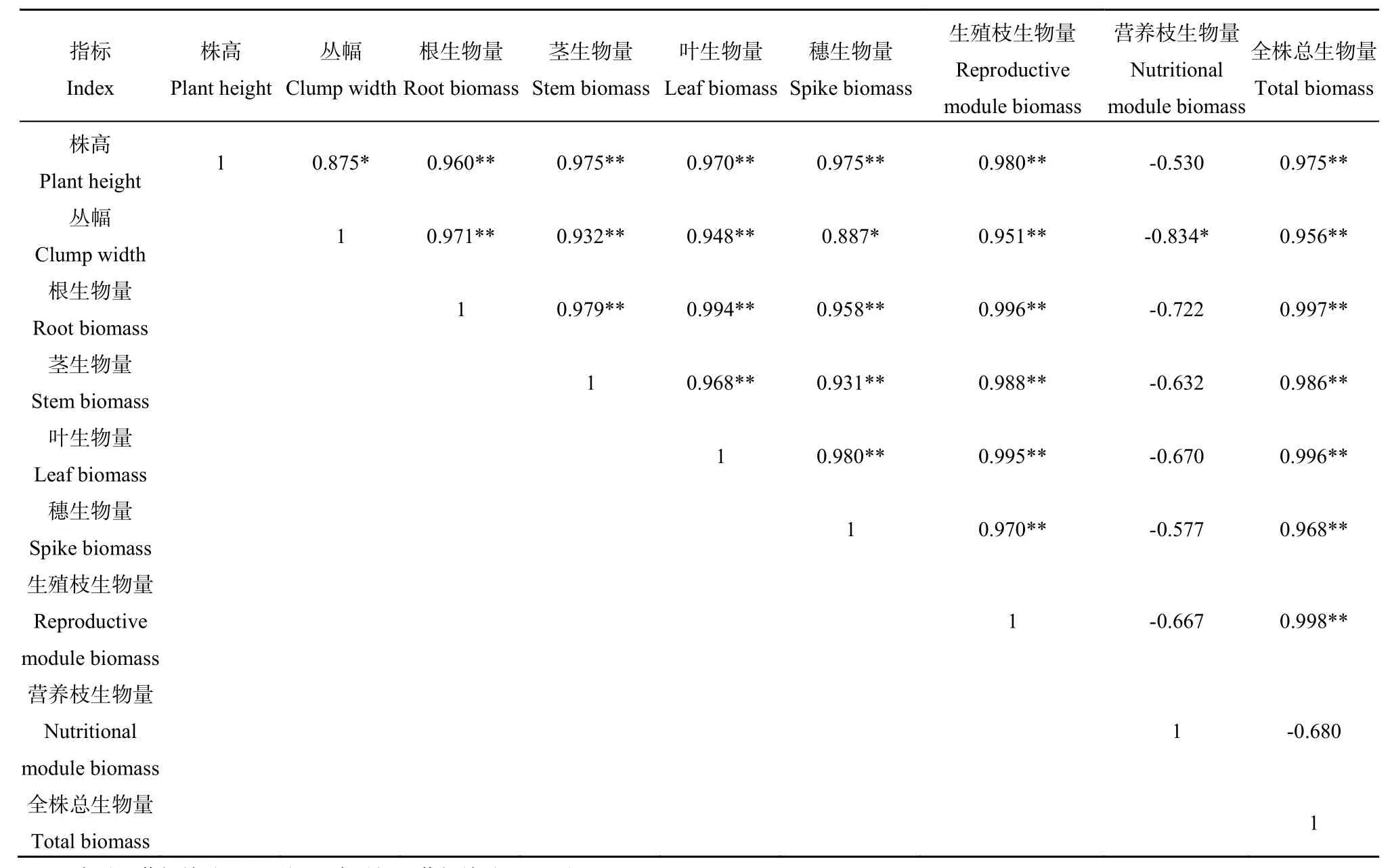
表3 沙芦草种群各构件生物量及其株高、丛幅间的相关性Table3 Correlation among module biomass, plant height and clump width of A. mongolicum population
本研究结果表明,围栏内外均表现为根生物量分配比最大,穗生物量分配比最小,且生殖枝生物量分配比大于营养枝。围封后,生殖枝、叶生物量分配比显著升高,营养枝生物量分配比显著降低。赵彬彬等(2009)在高寒草甸的研究表明,围栏禁牧后,放牧压力消失,植物对光照资源竞争增强,使其对光合器官的投资增加,而对生殖器官的投资减少。这与本研究结果不太一致,可能与高寒草甸系统的土壤养分、群落组成结构以及环境资源供给能力等与荒漠草原完全不同有关。除此之外,围封年限可能也是造成植物生物量分配发生变化的重要原因。生殖生长对不同环境的适应能力具有一定优越性(王静等,2005),围栏封育后环境中风险降低,种群更新一般优先于生殖生长。由此可知,生殖生长投资的增加是沙芦草种群在围栏封育后提高其适合度的重要因素(郑伟等,2013)。植物地上、地下生物量间的相关性影响着整个植物体的生长状况(Hui et al.,2006),而根冠比(R/S)是生物量在地下-地上之间分配的直接表现(Enquist et al.,2002)。围栏封育后,沙芦草种群的根冠比显著降低,说明实施围栏封育后,土壤风蚀得到一定缓解,土壤表面相对稳定,随着沙芦草植株的生长,冠幅和生物量不断增大,最终导致根冠比下降(王蕙等,2015)。
植物个体生长发育依赖于各构件生长发育间的协调,各构件生物量在个体生物量中所占的比率代表着同化产物向不同器官的分配比例和生长过程中各构件的协调关系(王伯荪等,1995)。本研究中,除营养枝生物量与各构件生物量、株高、丛幅间表现为负相关关系外,沙芦草种群各构件生物量间以及各构件生物量与株高、丛幅之间均存在显著或极显著正相关关系。前人(王蕙等,2015;周兵等,2015;张景慧等,2013)研究亦得出了相类似的结果,说明沙芦草种群株高、丛幅及其各构件生物量之间关系密切,同时说明围封是植物维持其种群持续更新能力和恢复其在群落中地位的重要措施之一(鲁为华等,2009)。
4 结论
(1)围封可以使沙芦草种群的高度、丛幅、盖度以及株丛生物量显著提高,而对其密度无显著影响。
(2)围栏封育后,沙芦草种群根、叶、穗生物量显著提高;此外,围封显著提高了沙芦草种群生殖枝和叶的生物量分配比,而显著降低了营养枝生物量分配比和根冠比。
(3)除营养枝生物量与丛幅间表现为显著负相关关系外,沙芦草种群各构件生物量之间均呈现极显著正相关性,各构件生物量与株高、丛幅之间亦呈现出显著或极显著正相关性。
BARTON K E. 2016. Low tolerance to simulated herbivory in Hawaiian seedlings despite induced changes in photosynthesis and biomass allocation [J]. Annals of Botany, 117(6): 1053-1062.
BLOOM A J, CHAPIN F S, MOONEY H A. 1985. Resource limitation in plants-An economic analogy [J]. Annual Review of Ecology &Systematics, 16(1): 363-392.
ENQUIST B J, NIKLAS K J. 2002. Global allocation rules for patterns of biomass partitioning in seed plants [J]. Science, 295(5559): 1517-1520.
HOSSEINZADEH G, JALILVAND H, TAMARTASH R. 2010. Short time impact of enclosure on vegetation cover, productivity and some physical and chemical soil properties [J]. Journal of Applied Sciences,10(18): 2001-2009.
HOVENDEN M J, NEWTON P C D, WILLS K E. 2014. Seasonal not annual rainfall determines grassland biomass response to carbon dioxide [J]. Nature, 511(7511): 583-586.
HUI D, JACKSON R B. 2006. Geographical and interannual variability in biomass partitioning in grassland ecosystems: a synthesis of field data[J]. New Phytologist, 169(1): 85-93.
KROONS H D, HUTCHINGS M J. 1995. Morphological Plasticity in Clonal Plants: The Foraging Concept Reconsidered [J]. Journal of Ecology, 83(1): 143-152.
LAMAS M I B, LARREGUY C, CARRERA A L, et al. 2013. Changes in plant cover and functional traits induced by grazing in the arid Patagonian Monte [J]. Acta Oecologica, 51(8): 66-73.
PAN Y X, WANG X P, JIA R L, et al. 2008. Spatial variability of surface soil moisture content in a re-vegetated desert area in Shapotou,Northern China [J]. Journal of Arid Environments, 72(9): 1675-1683.
POORTER H, NIKLAS K J, REICH P B, et al. 2012. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control [J]. New Phytologist, 193(1): 30-50.
WANG X P, LI X R, XIAO H L, et al. 2006. Evolutionary characteristics of the artificially revegetated shrub ecosystem in the Tengger Desert,northern China [J]. Ecological Research, 21(3): 415-424.
WEINER J. 2004. Allocation, plasticity and allometry in plants [J].Perspectives in Plant Ecology Evolution and Systematics, 6(4):207-215.
安渊, 王育青, 陈敏, 等. 1997. 沙地草场补播技术及其生态效益研究[J]. 草地学报, 5(1): 33-41.
金艳强, 李敬, 刘运通, 等. 2017. 围封对元江稀树灌草丛林下植被物种组成及生物量分配的影响[J]. 生态学杂志, 36(2): 343-348.
刘文清, 王国贤. 2003. 沙化草地旱作条件下混播人工草地的试验研究[J]. 中国草地学报, 25(2): 69-71.
刘忠宽, 汪诗平, 陈佐忠, 等. 2006. 不同放牧强度草原休牧后土壤养分和植物群落变化特征[J]. 生态学报, 26(6): 2048-2056.
鲁为华, 朱进忠, 王东江, 等. 2009. 天山北坡围栏封育条件下伊犁绢蒿幼苗分布格局及数量动态变化规律研究[J]. 草业学报, 18(4): 17-26.
罗永开, 方精云, 胡会峰. 2017. 山西芦芽山14种常见灌木生物量模型及生物量分配[J]. 植物生态学报, 41(1): 115-125.
王伯荪, 李鸣光, 彭少麟. 1995. 植物种群学[M]. 广州: 广东高等教育出版社.
王蕙, 王辉, 罗永忠, 等. 2015. 围封沙质天然草地植物的构件和个体生物量比较研究[J]. 草业学报, 24(9): 206-215.
王静, 杨持, 王铁娟. 2005. 放牧退化群落中冷蒿种群生物量资源分配的变化[J]. 应用生态学报, 16(12): 2316-2320.
王杨, 徐文婷, 熊高明, 等. 2017. 檵木生物量分配特征[J]. 植物生态学报, 41(1): 105-114.
肖遥, 陶冶, 张元明. 2014. 古尔班通古特沙漠4种荒漠草本植物不同生长期的生物量分配与叶片化学计量特征[J]. 植物生态学报, 38(9):929-940.
杨刚, 杨智明, 王思成, 等. 2003. 盐池四墩子试区草原围栏封育效果调研[J]. 农业科学研究, 24(1): 22-24.
闫玉春, 唐海萍, 辛晓平, 等. 2009. 围封对草地的影响研究进展[J]. 生态学报, 29(9): 5039-5046.
严月, 朱建军, 张彬, 等. 2017. 草原生态系统植物地下生物量分配及对全球变化的响应[J]. 植物生态学报, 41(5): 585-596.
杨勇, 刘爱军, 李兰花, 等. 2016. 围封对内蒙古典型草原群落特征及土壤性状的影响[J]. 草业学报, 25(5): 21-29.
宇万太, 于永强. 2001. 植物地下生物量研究进展[J]. 应用生态学报,12(6): 927-932.
云锦凤, 米福贵. 1989. 冰草属牧草的种类与分布[J]. 中国草地学报,(3): 14-17.
张景慧, 祁瑜, 王艳, 等. 2012. 放牧对贝加尔针茅草原植物个体和群落生物量分配的影响[J]. 北京师范大学学报(自然科学版), 48(3):270-275.
赵彬彬, 牛克昌, 杜国祯. 2009. 放牧对青藏高原东缘高寒草甸群落 27种植物地上生物量分配的影响[J]. 生态学报, 29(3): 1596-1606.
赵有璋. 2004. 种草养羊技术[M]. 北京: 中国农业出版社.
郑伟, 于辉. 2013. 围栏封育对伊犁绢蒿种群构件生长和生物量分配的影响[J]. 草地学报, 21(1): 42-49.
周兵, 闫小红, 肖宜安, 等. 2015. 不同生境下入侵植物胜红蓟种群构件生物量分配特性[J]. 生态学报, 35(8): 2602-2608.
Effects of Enclosure on Biomass Allocation Characteristics of Agropyron mongolicum Population in Desert Steppe
ZHAO Panpan1,2, SHAO Wenshan1,2, JIN Changqing1,2, SONG Lixiao1,2, GONG Shipei1,2, LI Guoqi1,2*
1. Breeding Base for State Key Laboratory of Land Degradation and Ecological Restoration in Northwest China, Ningxia University, Yinchuan 750021, China;2. Key Laboratory for Recovery and Restoration of Degraded Ecosystem in North-western China of Ministry of Education, Ningxia University,Yinchuan 750021, China
This study explored the effect of enclosure on the resources allocation pattern and ecological strategy of plant in the desert steppe region in Ningxia. The biomass and allocation pattern of each modules and the correlation among module biomass were measured in the unsealed and enclosed plots. The results showed that: (1) Height, clump width, coverage and total biomass of Agropyron mongolicum population were significantly increased after enclosure with the rates of increase being 78.96% (P=0.040),63.50% (P=0.013), 50.89% (P=0.032), 205.38% (P=0.022), respectively, while the density did not change significantly (P=0.330). (2)Enclosure had a significant effect on the biomass of the modules of A. mongolicum population. Compared with the outside fence,root, leaf and spike biomass increased significantlywith the rates of increase being 183.52% (P=0.020), 334.09% (P=0.011), 381.25%(P=0.005), respectively. The root biomass that included inside and outside of the fence were the largest(Inside is 38.53g, outside is 13.59g), and the biomass of the spike was the smallest(Inside is 6.16g,outside is 1.28g). (3) After enclosure, the allocation proportion of the biomass of A. mongolicum changed in different degrees. The biomass allocation ratio of reproductive branches and leaves increased significantly, with the rates of increase being 31.45% (P=0.020) and 4.20% (P=0.000), respectively. While the biomass allocation ratio of nutritional branches and root shoot ratio decreased significantly with the rates of decrease being 281.62%(P=0.002) and 13.33% (P=0.023), respectively. (4) Except the biomass of vegetative branches and clump width had significant negative correlation relationship (P=0.039), among the modular biomass of A. mongolicum showed an extremely significant positive correlation, and there was a significant or very significant positive correlation between biomass and plant height, clump width. The enclosure had a significant effect on the biomass and distribution pattern of the modules of A. mongolicum population. It is suggested that enclosure is one of the important measures for this population of A. mongolicum to connect flexibility and to restore its role in the community.
Agropyron mongolicum; enclosure; population module; biomass allocation
10.16258/j.cnki.1674-5906.2017.12.004
S812.29; X173
A
1674-5906(2017)12-2024-06
赵盼盼, 邵文山, 靳长青, 宋立肖, 龚诗佩, 李国旗. 2017. 围封对荒漠草原沙芦草种群构件生物量分配特性的影响[J]. 生态环境学报, 26(12): 2024-2029.
ZHAO Panpan, SHAO Wenshan, JIN Changqing, SONG Lixiao, GONG Shipei, LI Guoqi. 2017. Effects of enclosure on biomass allocation characteristics of Agropyron mongolicum population in desert steppe [J]. Ecology and Environmental Sciences, 26(12):2024-2029.
国家自然科学基金项目(31540007)
赵盼盼(1991年生),女,硕士研究生,主要从事植物生态学、土壤生态学研究。E-mail: 1751327789@qq.com
*通信作者:李国旗,研究员,主要从事植物生态学等方面的研究。E-mail: guoqilee@163.com
2017-09-11
猜你喜欢
草地学报(2022年9期)2022-09-29 11:22:48
天津农林科技(2022年2期)2022-04-19 10:48:14
河南畜牧兽医(2022年3期)2022-04-13 09:12:58
绿色科技(2021年10期)2021-06-23 03:13:06
东北石油大学学报(2021年1期)2021-03-25 07:03:40
现代养生·上半月(2020年11期)2020-11-13 03:44:37
散文选刊·下半月(2019年2期)2019-03-15 06:32:46
草业学报(2019年2期)2019-02-25 01:57:16
山东农业科学(2018年7期)2018-10-22 07:01:10
新疆地质(2016年4期)2016-02-28 19:18:47
