
基于骨架化模型的染色体形态表征
2017-12-26 08:26王一鸣
沈阳师范大学学报(自然科学版) 2017年4期
高 明, 王 婷, 王一鸣, 田 宁
(沈阳师范大学 物理科学与技术学院, 沈阳 110034)
材料科学
基于骨架化模型的染色体形态表征
高 明, 王 婷, 王一鸣, 田 宁
(沈阳师范大学 物理科学与技术学院, 沈阳 110034)
染色体作为遗传物质的重要载体,其空间构象(主要指染色体数量、结构、定位等形态信息)与包括基因表达、表观遗传在内的一些基本的细胞生命活动关联紧密。染色体形态改变是引起人类某些重大疾病(如唐氏综合症、恶性肿瘤等)的重要原因之一。为实现染色体形态的准确、量化分析,针对光学成像设备采集染色体图像含冗杂信息过多的问题,基于骨架化理论,通过二值转换及距离变换等计算方法,抽取染色体的拓扑结构,以减少冗余信息。进一步,提出利用骨架长度、端粒数目及着丝粒分界比,数字化表征染色体的拓扑形态。因此,利用自动化手段建立了一种量化表征染色体形态信息的方法,这将为染色体识别及形态研究提供重要思路。
骨架化; 染色体; 拓扑形态
0 引 言
染色体作为遗传物质的重要载体,其生物形态与表观修饰,遗传与变异,及重大疾病的产生关联紧密[1-3]。染色体形态主要通过荧光标记技术(如荧光原位杂交)结合光学成像技术而获得[4-5]。并通过染色体图像的体积、表面积、形状等几何指标来表征染色体的生物形态[2]。这些几何指标只是一种半定量的形态表征手段,它们的精确度往往受到光学成像技术分辨率的限制。因此需要更完善的方法来全面表征染色体的生物形态[6]。
在自动化研究中,骨架提取及边缘检测在图像识别领域中应用广泛[7-10]。其中利用图像骨架来表征图像形态,是有效剔除冗杂信息,保留图像拓扑结构特征的有效技术手段[8]。骨架化方法已广泛应用于植物根茎模拟、叶片脉络分析、肺血管重建等领域[11-13]。鉴于此,本文以骨架化理论为基础,获得染色体的拓扑结构(即染色体骨架),并以长度、端点数目、着丝粒分界比等指标数字化表征染色体骨架,以区分不同染色体的形状特征。本文提出的染色体骨架分析,将为染色体识别及形态研究提供重要的途径。
1 实验方法
1.1 样品制备
1) 将雌性ICR小鼠于处死前4 h注射3 μg/g的秋水仙素,4 h后脱颈椎处死,摘取后肢股骨;
2) 用低渗液(0.075 mol/L KCl)反复冲洗骨髓腔,收集骨髓细胞;
3) 再将骨髓细胞移入预温至37 ℃的低渗液至8 mL,置于37℃水浴锅内30 min;
4) 然后,加入1 mL新制固定液(甲醇∶冰醋酸=3∶1),混匀后以1 500 r/min离心5 min;
5) 弃上清,再加入5 mL固定液,混匀固定15 min,离心5 min后,再弃上清;
6) 再加入0.3 mL固定液制成细胞悬混液;
7) 在干净、湿冷载玻片上20 cm处滴2滴细胞悬液;
8) 气干后,DAPI染色5 min,清洗后,即可显微镜下观察。
1.2 染色体骨架化
1) 图像预处理
光学系统成像时,在数据采集与传输的过程中,常受到外界干扰,从而对光学图像引入噪声及失真。这些噪声或图像失真使得人们对荧光图像的观察受到影响,对图像信息的理解受到干扰。成像系统采集的图像g,与真实图像f的关系为g=f×h+n。其中h为成像系统的点扩散函数,n为噪声。鉴于此,本文图像预处理过程,首先为去噪。荧光显微镜产生的主要噪声为椒盐噪声,中值滤波是有效去除椒盐噪声的方法[1]。随后进行盲反卷积操作。根据成像设备的参数(激发波长为358 nm,物镜为尼康100倍油镜,物镜油折射率为1.515),构建系统的点扩散函数,再盲反卷积,以尽量恢复真实的荧光图像。
2) 图像分割
光学图像获得后,需要将图片中前景与背景分离开,以便研究前景图像的形态特点。图像分割是指将数字图像根据不同的特征,细分为若干个图像子区域的过程。每个子区域对应的是一个具有某种共同视觉特性的像素集合。本文以人机交互的方式设定分割阈值,即人眼的视觉识别与图像数字直方图的分界协同确定阈值,并将灰度大于阈值的像素点设定为前景图像,小于阈值的设定为背景图像。
3) 二值化
图像的二值化就是将图像上的所有像素点的灰度值分别设置为0或255这两个不同的值,这样就将整个图像的视觉效果呈现出只有黑和白的(二值)图像。在这里将要处理图像的前景像素的灰度值均设置为255,图像的背景像素的灰度值均设置为0,使灰度图像变为黑白(二值)图像。
4) 骨架提取
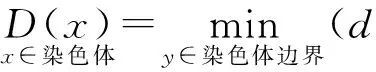
5) 图像后处理
由不规则的染色体边界及图像噪声等因素的干扰,所抽取的染色体骨架存在着很多毛刺(异化分支)。这种畸变(毛刺)将影响染色体拓扑形态的表征。本文以分支长度和方向为指标,判定原骨架中各分支是否为毛刺,以保证修剪后骨架质量更能准确地表征染色体拓扑形态。分支长度的划分标准一般为小于最长分支1/3的视为毛刺。分支方向的划分标准为分叉中夹角小于90°的一般视为毛刺。自动化修剪后骨架仍需人工检验,以摒除计算误差。
1.3 染色体骨架分析
1) 骨架长度
对二值图像骨架提取后的骨架进行长度的计算。结合单个像素代表的几何大小(0.84 μm×0.84 μm),计算染色体骨架的8邻域长度,获得染色体骨架尺寸。
2) 端粒数目
处理后的图像骨架由单个像素点连接而成,每个染色体骨架端点数目代表染色体的端粒数目。(端点在8邻域范围内只有一个像素点)。分别对每个染色体骨架进行观察、记录并整理出端粒数。
3) 着丝粒分界比
不同染色体的着丝粒位置也各有不同,对图像进行骨架提取的染色体骨架中分叉点代表染色体的着丝粒位置。(分叉点在8邻域范围内有≥3个像素点)。一条染色体骨架中,端点到分叉点的距离之比,定义为着丝粒分界比。
2 实验结果
2.1 染色体图像预处理、分割、二值化结果
依实验方法,原始图片(图1a)经2×2中值滤波去除噪声(图1b),而后经盲反卷积修复(图1c),图像质量得到了极大的改善。以灰度64为阈值分界,分割染色体图像的前景与背景,图片中灰度值64~255的像素选定为染色体区域,即为前景目标;灰度值0~63的像素设定为背景目标,图1d显示分割结果。随后,将分割图像进行二值变换,灰度图转变为二值图(图1e)。

图1 染色体原始图像的预处理(a~c)、分割(d)及二值转换(e)Fig.1 Preprocessing(a~c), segmentation(d) and binaryzation(e) of original image of chromosomes
2.2 染色体骨架提取及毛刺去除结果
由上节骨架提取算法,从染色体二值图像中可获得染色体骨架,可以看到染色体的初提取骨架存在很多毛刺(图2a),这些毛刺是一种非结构性分支,由图像噪声及细节引入。以分支长度和方向为指标,设置毛刺划分门限,并以人类视觉观察为最终较验体系,对初提取骨架进行毛刺筛查及剔除,毛刺去除后的染色体最终骨架见图2b和图2c。可见,最终骨架可正确反映染色体臂长及着丝粒位置,是染色体结构的准确体现。
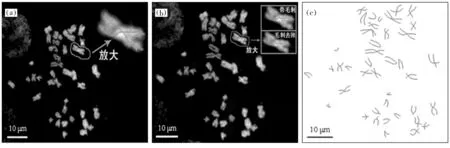
图2 染色体骨架初提取示意(a)及毛刺去除效果(b,c)Fig.2 Initial skeleton extraction of chromosomes (a) and deburring (b, c)
2.3 染色体骨架分析结果
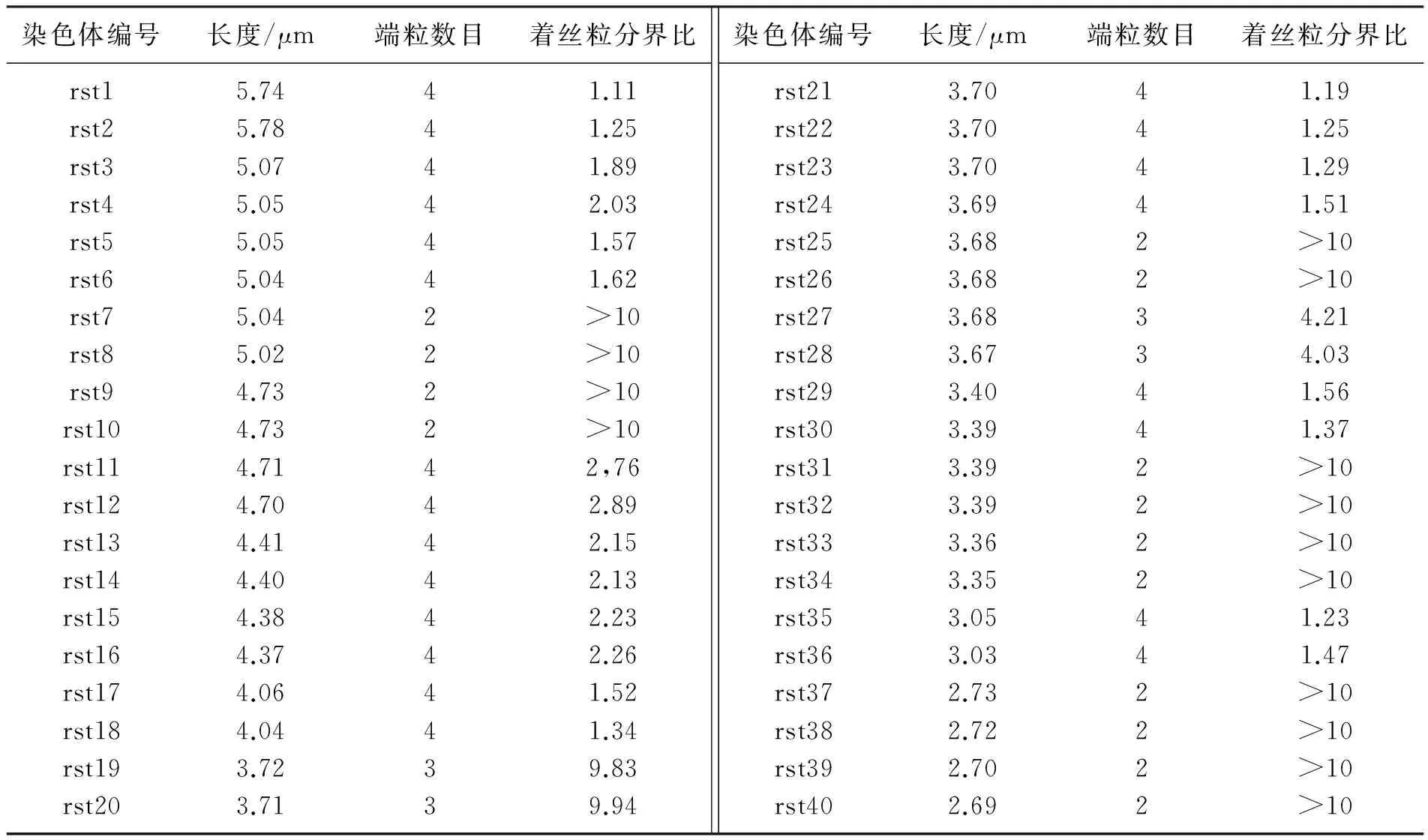
本文以长度、端粒数目及着丝粒分界比来分析染色体骨架(图3),从而确定染色体的形态特点。本文中,以染色体骨架长度长短的顺序依次命名染色体(图4),并在表1中显示了ICR雌性小鼠中期骨髓细胞40条染色体骨架的形态分析结果。
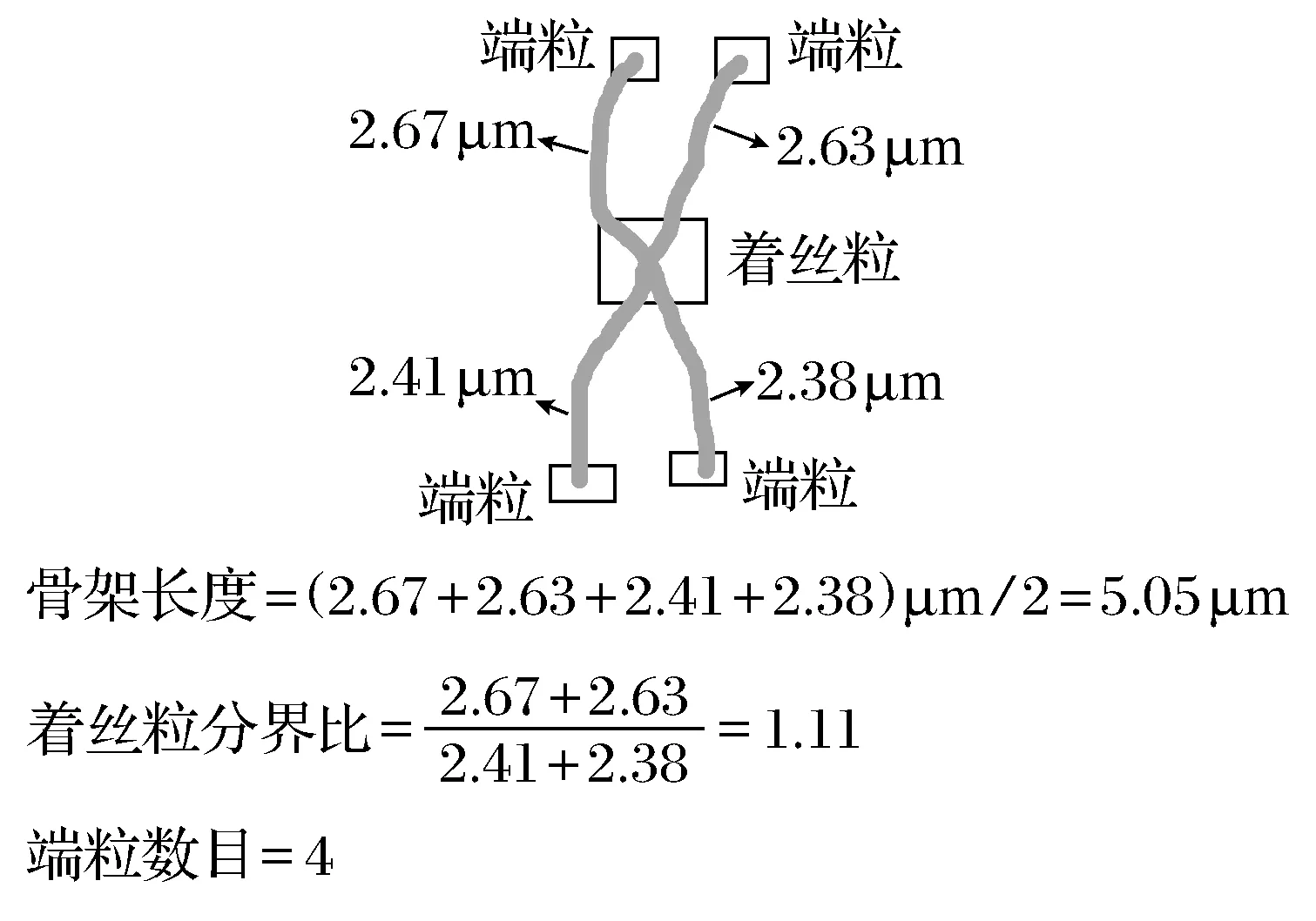
图3 染色体骨架分析示意Fig.3 Illustration of chromosome skeleton analysis
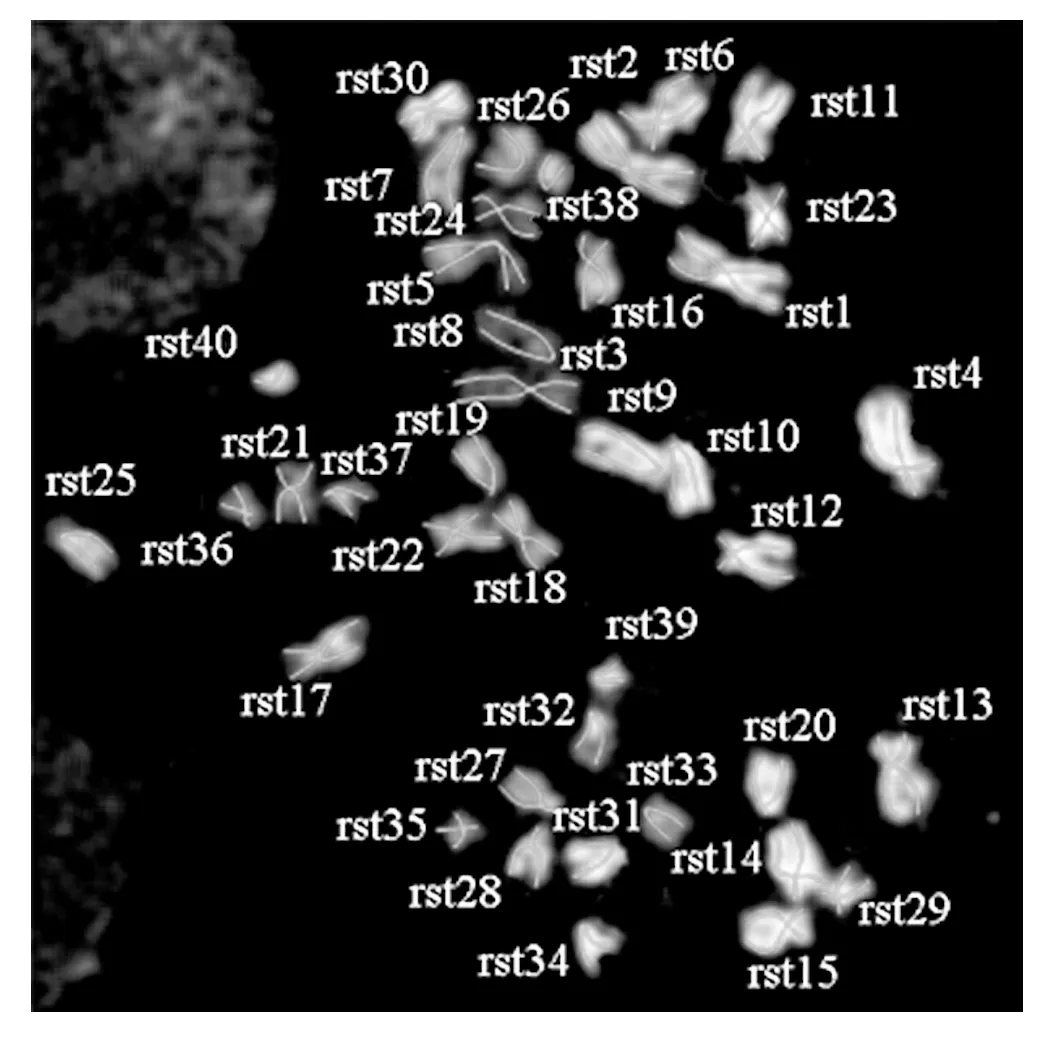
图4 依骨架长度命名染色体Fig.4 Chromosomes named after skeleton length
染色体编号长度/μm端粒数目着丝粒分界比rst15.7441.11rst25.7841.25rst35.0741.89rst45.0542.03rst55.0541.57rst65.0441.62rst75.042>10rst85.022>10rst94.732>10rst104.732>10rst114.7142,76rst124.7042.89rst134.4142.15rst144.4042.13rst154.3842.23rst164.3742.26rst174.0641.52rst184.0441.34rst193.7239.83rst203.7139.94染色体编号长度/μm端粒数目着丝粒分界比rst213.7041.19rst223.7041.25rst233.7041.29rst243.6941.51rst253.682>10rst263.682>10rst273.6834.21rst283.6734.03rst293.4041.56rst303.3941.37rst313.392>10rst323.392>10rst333.362>10rst343.352>10rst353.0541.23rst363.0341.47rst372.732>10rst382.722>10rst392.702>10rst402.692>10
3 讨 论
染色体是遗传物质在细胞分裂中期的存在形式,是染色质紧密蜷缩的结果,其作为遗传物质的重要载体, 其形态研究对发育、遗传及变异等基础研究有重要作用,在临床应用中, 也是白血病等恶性血液病的重要诊断依据[1,14]。生物医学中, 染色体形态研究的常用方法是基于Giemsa染色的G带带型分析[15-16],该方法比较成熟,可准确呈现染色体组型, 但该方法需要较多的生化操作, 且由于成像及染色的影响容易造成带纹压缩误差。 因此,本文尝试提出一种更简洁方便的染色体形态研究方法。 利用骨架化理论分析ICR小鼠中期骨髓细胞的染色体拓扑结构,并以骨架长度、端粒数目及着丝粒分界比(着丝粒位置)标记染色体形态。 一副图像的骨架是这幅图像的重要形态特征,是图像拓扑结构的直观描述, 通常以计算图像中轴线的方式获得骨架。 利用骨架化方法获得染色体的拓扑信息,是利用计算机技术辅助人类视觉观察的重要改进, 这提高了染色体识别的工作效率,将染色体形态研究推向自动化。 本文获得的ICR雌性小鼠染色体骨架形态包括X型、V型及U型, 其中X型对应着中着丝粒和近中着丝粒染色体,V型和U型对应的是近端着丝粒和端着丝粒染色体。 着丝粒分界比为1~1.70的染色体定义为中着丝粒染色体;1.71~3.00的定义为近中着丝粒染色体; 3.01~7.00的定义为近端着丝粒染色体;大于7.00的定义为端着丝粒染色体[16]。按此定义,本文数据显示ICR雌性小鼠骨髓细胞中共有40条染色体,长度范围为2.69~5.74 μm,其中14个为中着丝粒,8个为近中着丝粒,2个为近端着丝粒,16个为端着丝粒。 不同品系的实验小鼠在基因表达、遗传与变异、及恶性肿瘤研究中均具有非常重要的应用。 ICR小鼠是药物筛查及病理研究中常用的动物模型,非常广泛地应用于药理、毒理、肿瘤等科学研究中[1,6-7]。本文对ICR雌性小鼠染色体形态的研究也为基于该品系小鼠的相关研究应用提供重要参考。
本文利用去噪、盲反卷积技术对染色体图像进行预处理,并通过三维自动分割技术分离出染色体前景图像;基于骨架化理论,应用距离变换算法获得了染色体骨架;最后,通过长度、端粒数目及着丝粒分界比量化表征了染色体的拓扑形态。该方法可以准确提取染色体的结构化骨架,并能数字化染色体形态。这对于基于染色体形态的临床诊断及基础研究有重要实用价值。
[ 1 ]田宁. 双光子成像量化观测小鼠卵母细胞染色体三维构象[D]. 北京:清华大学, 2013.
[ 2 ]WANG T,HAN J,DUAN X,et al. The toxic effects and possible mechanisms of Bisphenol A on oocyte maturation of porcine in vitro[J]. Oncotarget, 2016,7(22):32554-32565.
[ 3 ]HE Y,LI X D,WANG D N,et al. Acute promyelocytic leukaemia with a PML-RARA insertional translocation and a chromosome 21 abnormality in XYY syndrome: Case report[J]. The Journal of International Medical Research, 2014,42(6):1363-1373.
[ 4 ]CHEN K H,XU S C,CHEN H Y,et al. Utility of fluorescence in situ hybridization for ploidy and p57 immunostaining in discriminating hydatidiform moles[J]. Biochemical and Biophysical Research Communications, 2014,446(2):555-560.
[ 5 ]ZHU C C,ZHANG Y,DUAN X,et al. Toxic effects of HT-2 toxin on mouse oocytes and its possible mechanisms[J]. Archives of Toxicology, 2016,90(6):1495-1505.
[ 6 ]TIAN N,ZHANG L,ZHENG J H,et al. Three-dimensional quantitative analysis of chromosomes in the oocytes of aging mice during meiosis I in vitro[J]. Theriogenology, 2013,79(2):249-256.
[ 7 ]朵建英,王卫,佟巍,等. 肠道病毒71型(EV71)对ICR小鼠的感染[J]. 中国比较医学杂志, 2009,19(5):41-46.
[ 8 ]韩国强,田绪红,李志垣,等. 三维图像骨架化方法综述[J]. 小型微型计算机系统, 2007,28(9):1695-1699.
[ 9 ]姜炳旭,刘杰,孙可. Sobel边缘检测的细化[J]. 沈阳师范大学学报(自然科学版), 2010,28(4):503-506.
[10]WANG C F,GUI C P,LIU H K,et al. An Image Skeletonization-Based Tool for Pollen Tube Morphology Analysis and Phenotyping[J]. J Integr Plant Biol, 2013,55(2):131-141.
[11]李马并臻,周学成,张常玲,等. 基于骨架模型的植物根系三维构型可视化方法[J]. 计算机工程与设计, 2014,35(11):3913-3917.
[12]郭斯羽,董红霞,张翌. 一种用于植物叶片图像骨架提取的去毛刺方法[J]. 电子测量与仪器学报, 2013,27(1):52-56.
[13]陈刚,吕煊,王志成,等. 肺CT图像的血管骨架化方法[J]. 计算机科学, 2013,40(5):274-278.
[14]MILICA S,BILJANA TODOTIC Z,ZELJKA T,et al. Additional chromosome abnormalities in Philadelphia-Chromosome-Positive Chronic Myeloid Leukaemia[J]. Chromosome Research, 2013,21(1):S104-S104.
[15]MARTA K G,ALEKSANDRA G,OLGA S,et al. Karyotyping of Chinchilla lanigera Mol.(Rodentia, Chinchillidae)[J]. Caryologia, 2015,68(2):138-146.
[16]王曦,李红霞,李慧玲,等. 615小鼠染色体组型与G带带型分析[J]. 中国比较医学杂志, 2007,17(8):467-469.
Morphologicalcharacterizationofchromosomesbasedonskeletonmodel
GAOMing,WANGTing,WANGYiming,TIANNing
(College of Physics Science and Technology, Shenyang Normal University, Shenyang 110034, China)
Chromosome is the important carrier of genetic material, the spatial configuration(namely chromosomal numbers, structure and localization) of which has a close link with the fundamental biological processes in cell such as gene expression and epigenetic inheritance. Chromosomal morphology aberrations are related to the serious disease(such as down’s syndrome, malignant tumor) which threaten human health. To realize the accurate and quantitative analysis of chromosomal morphologies, this work extracted the topology structure of chromosomes based on the skeletonization theory of binary image conversion and distance transform, to overcome the problem of too much miscellaneous information caused by optical imaging equipment. Further, topological morphology of chromosomes was quantitatively characterized by skeleton length, telomere number and centromere boundary ratio. Based on the automation, this paper established a method of quantitatively characterizing the chromosomal morphology, and this paper will provide important thought for chromosome recognition and morphological study.
skeletonization; chromosome; topological morphology
2017-07-18。
辽宁省科技厅自然科学基金资助项目(2015020715)。
高 明(1964-),男,辽宁沈阳人,沈阳师范大学教授,博士;
田 宁(1985-),女,辽宁沈阳人,沈阳师范大学副教授,博士。
1673-5862(2017)04-0395-05
Q63
A
10.3969/ j.issn.1673-5862.2017.04.003
猜你喜欢
生物技术进展(2022年1期)2022-02-11
中国金属通报(2021年6期)2021-07-01
高技术通讯(2021年2期)2021-04-13
装备制造技术(2020年4期)2020-12-25
装备制造技术(2019年12期)2019-12-25
科学之谜(2018年9期)2018-12-17
测控技术(2018年10期)2018-11-25
计算机技术与发展(2017年12期)2017-12-20
制造技术与机床(2017年10期)2017-11-28
计算机应用(2016年10期)2017-05-12
