
青冈栎次生林种群结构及动态特征
2017-12-10 18:39:24曾思齐龙时胜仇建友俞军剑
中南林业科技大学学报 2017年11期
胡 满,曾思齐,龙时胜,仇建友,俞军剑
(1.中南林业科技大学,湖南 长沙 410004;2.安化县林业局马路林业站,湖南 安化 413506)
青冈栎次生林种群结构及动态特征
胡 满1,曾思齐1,龙时胜1,仇建友2,俞军剑2
(1.中南林业科技大学,湖南 长沙 410004;2.安化县林业局马路林业站,湖南 安化 413506)
青冈栎作为我国南方亚热带地区主要建群树种之一,对南方生态系统的稳定起着至关重要的作用。通过样地调查和数据统计方法,分析其种群年龄结构特征,编制其种群的静态生命表,对死亡率、存活、消失率曲线进行分析,并通过时间序列预测、种群数量化动态方法的运用,揭示天然次生林中青冈栎种群结构及动态特征。结果表明:青冈栎种群属于增长型,种群的动态变化指数值为0.0103,表明该种群趋于稳定型;存活曲线趋于
Deevey-Ⅱ型,各龄级种群有着相近的死亡率;时间序列预测分析表明,在未来10、20、30、40 a后青冈栎种群老龄个体逐渐增多,幼龄株数逐渐减少,维持种群长期稳定性困难,因此,采取适当的人工措施是保持青冈栎种群天然更新和种群稳定的关键。
青冈栎;种群结构;静态生命表;存活曲线;时间序列预测
种群以植物种群结构作为一大基本特征,不仅能反映某一种群的动态变化和趋势,在一定程度上,还能折射出其所在环境条件以及群落地位[1-2]。其动态变化,体现出一定环境中,种群生物特征的适应性,是对种群生活史特征、基本结构予以了解的有效工具[3]。
青冈栎Cyclobalanopsis glauca为壳斗科青冈属常绿落叶乔木,在我国长江以南广泛分布[4],但是由于不合理经营利用,大部分青冈栎林都属于自然更新形成的天然次生林。目前对于青冈栎次生林的研究主要集中于竞争关系[5]、林分结构[6]与种苗培育[7-8],关于青冈栎种群结构动态方面的研究较少。以湖南省平江县中南林业科技大学芦头实验林场青冈栎次生林为研究对象,研究青冈栎的种群结构及其动态特征,为青冈栎次生林的经营管理以及合理保护、以及恢复策略等的制定奠定了参考依据。
1 研究区概况
研究区位于湖南省平江县中南林业科技大学芦头实验林场,该林场处于岳阳市东南面,地理位置为 113°51′52″E ~ 113°58′24″E,28°31′17″N ~28°38′00″N,中山地貌,海拔在 120 ~ 1 300 m,林场拥有丰富的动植物资源以及多种珍稀濒危动植物。有种子植物154科606属1 200种,被子植物147科588 属1 174种。
2 材料与方法
2.1 样地设置与调查
在研究地区分别选择具有代表性的地段,设置5块20 m×20 m的标准地,对达到起测径阶(D≥5 cm)的乔木树种做每木检尺,并记录乔木的树种、胸径、树高、冠幅、坐标等因子;未达到起测径阶(D<5 cm)的树种,调查其树种、树高、地径、坐标,并在每个样地中设置4块5 m×5 m的小样方,在基径处伐倒,测定其年龄。同时对样地的基本信息(经纬度、海拔、坡度、坡位、坡向、地形、郁闭度、枯落物厚度等)进行调查并记录(见表1)。
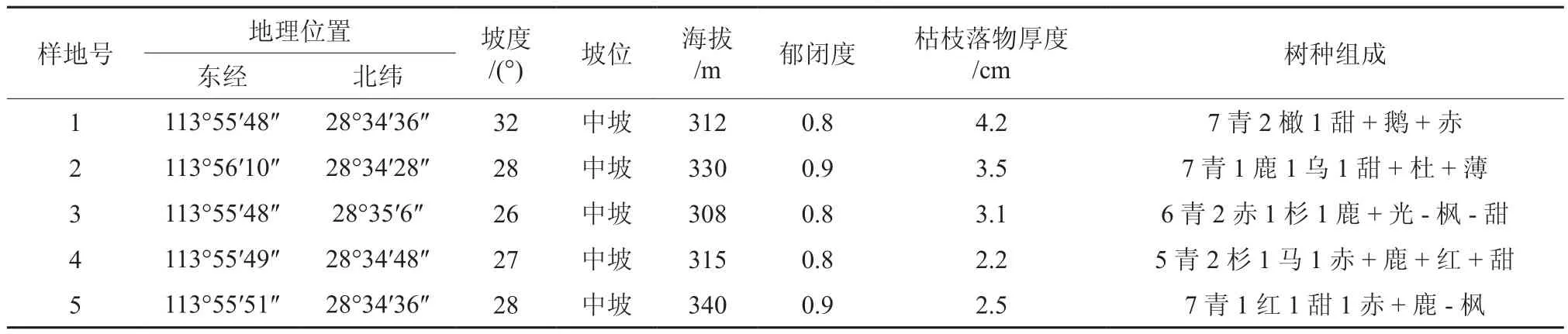
表1 样地基本信息†Table 1 Basic information of experimental plots
2.2 龄级划分
国内大部分学者学者都采用径级结构替代年龄结构的方法[9-11],本研究采用年龄-基径(BD)回归与解析木结合的方法,获取青冈栎的具体年龄。以年龄(A)为因变量,基径(BD)为自变量,通过年龄-基径回归方法,拟合得到方程:
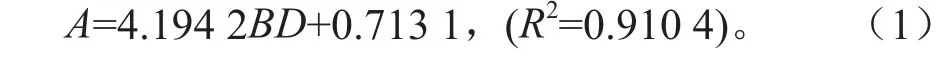
通过解析木资料,以年龄(A)为自变量,胸径(D)为因变量,拟合得到Richard方程结果最优:
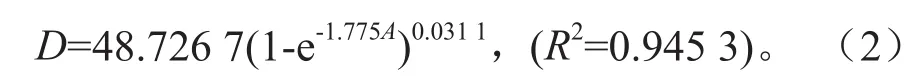
根据式(1)和(2),可以得到每株青冈栎的年龄,根据《国家森林资源连续清查湖南省第七次复查操作细则》,栎类天然林的龄级划分为20 a,但从调查的青冈栎年龄分布情况来看,以20 a一个龄级进行划分并不合理,因此,本研究以5 a一个龄级对青冈栎龄级进行划分,可划分为11个龄级:Ⅰ龄级(A≤5 a)、Ⅱ龄级(5 a≤A<10 a)、Ⅲ龄级(10 a≤A<15 a)、Ⅳ龄级(15 a≤A<20 a)、Ⅴ龄级(20 a≤A<25 a)、Ⅵ龄级(25 a≤A<30 a)、Ⅶ龄级(30 a≤A<35 a)、Ⅷ龄级(35 a≤A<40 a)、Ⅸ龄级(40 a≤A<45 a)、Ⅹ龄级(45 a≤A<50 a)、Ⅺ龄级(A≥50 a)。
2.3 种群动态数量化方法
种群动态数量化方法[12]一般用来定量描述种群动态:

式中,Vn为种群n到n+1级的个体数量变化;Sn+1分别为第n和n+1级种群个体数。
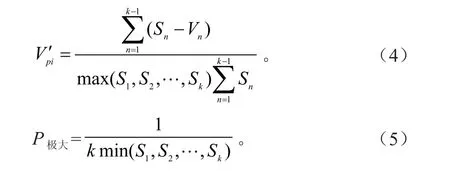
式中,V′pi为整个种群结构的数量动态变化指数;k为大小级数量;S为大小级个体数。Vn和V′pi取正、负、零值时,分别对个体数量稳定、衰退、增长的动态关系予以体现。P极大为遭受外界干扰时,种群结构对外界干扰的敏感性指数。
2.4 静态生命表的编制
结合种群不同龄级的株数,来对该种群的静态生命表进行编制,详细编制方法见参考文献[13]。在编制青冈栎种群生命表时,会出现死亡率为负数的情况,这是由于青冈栎属于天然次生林,静态生命表的3个假设并不能同时满足,负值情况虽与数学假设不符,但依然可以提供有用的生态数据。对此,江洪等人在编制云杉静态生命表时采用了匀滑技术[14],本研究也将使用这种方法进行。
2.5 种群数量动态预测
此处通过时间序列分析的运用[15-16],来对青冈栎种群未来数量加以预测、模拟。

式(6)中:Mt表示在未来n年时t龄级的种群大小,n表示需要预测的未来时间年限,t为龄级,Xk为当前k龄级的种群大小。
3 结果与分析
3.1 青冈栎种群分布现状及结构特征
结合下图可以发现,此种群龄级的形状为金字塔型,顶部相对狭窄,而基部较宽。在统计之后发现,I到IV龄级个体数量相对较多,约为群体数量的85.49%,Ⅴ-Ⅷ龄级占整个种群个体总数的13.28%,Ⅸ-Ⅺ龄级占整个种群个体总数的1.23%。

图1 青冈栎次生林种群龄级结构Fig.1 Population age structure of Cyclobalanopsis glauca secondary forest
基于种群动态数量化方法来对青冈栎种群各龄级之间的动态变化进行分析(见表2),结果表明:青冈栎种群在Ⅰ龄级向Ⅱ龄级的发育过程中,其动态指数值V1=-0.167 4<0,说明处于Ⅰ龄级的个体数少于下一阶段的个体数。若外界环境对种群进行随机干扰,则动态指数值V′pi=0.010 3,趋近于0,相应的敏感性指数P极大=0.05,说明青冈栎次生林中青冈栎种群总体呈现为增长型种群,但种群增长幅度不大,而且种群结构容易受到外界环境的干扰。
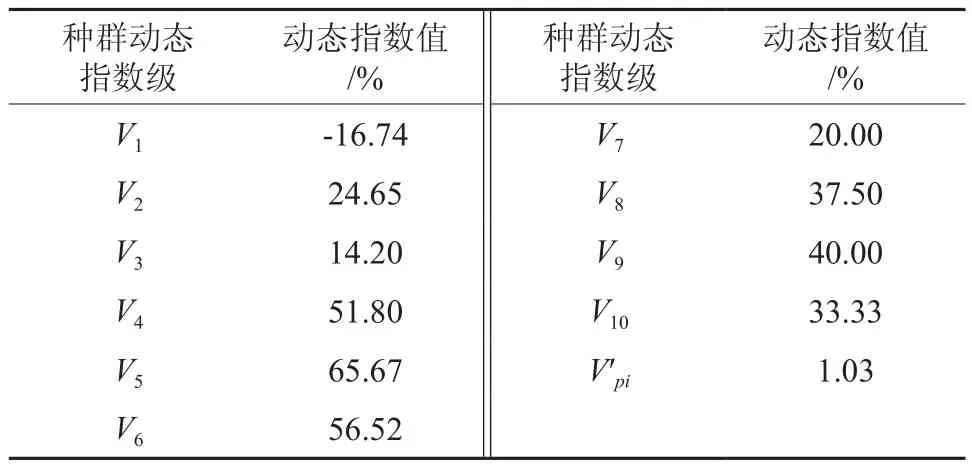
表2 青冈栎次生林种群龄级结构的动态变化指数Table 2 Dynamic index of age structure of secondary forest of Cyclobalanopsis glauca
3.2 静态生命表
青冈栎种群属于天然次生林,为避免死亡率出现负值的情况,通过匀滑技术的运用,来匀滑处理该种群数据,自变量、因变量分别为各龄级、存活株数,构建得到方程如下:
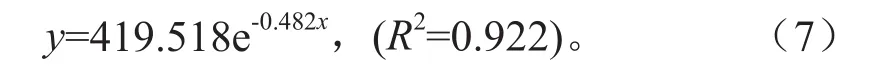
由式(7)可以得到匀滑后得到各龄级的植株数为ax,最终编制的青冈栎种群静态生命表(见表3)。
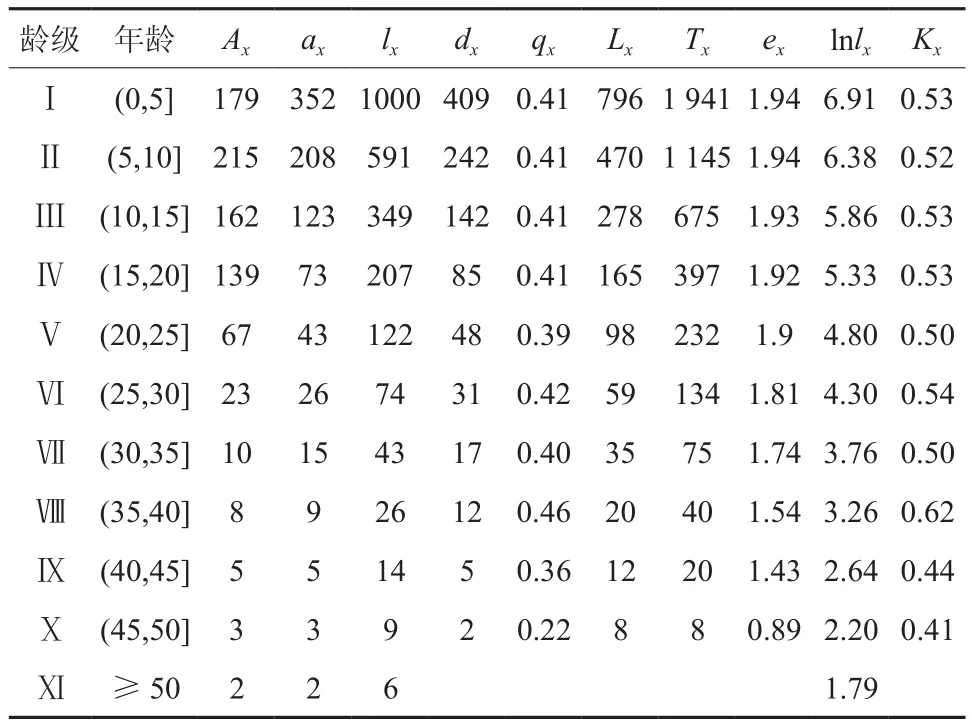
表3 青冈栎种群静态生命表†Table 3 Static life table of Cyclobalanopsis glauca population
3.3 存活曲线、死亡率及消失率曲线
存活曲线,即以存活个体数量作为基础,可对在各龄段,树木个体的存活状态进行反映[17]。本文以各龄级的标准化存活数量进行存活曲线的绘制(图2)。Deevey存活曲线的类型为3种,即曲线凸型、曲线对角型、曲线凹型。第1种表示在达到生理寿命前,死亡的个体仅为少数,即几乎全部均可达到生理寿命;第2种即各年龄具有相同的死亡数;第3种,即死亡率较高的龄段为幼年期[18]。
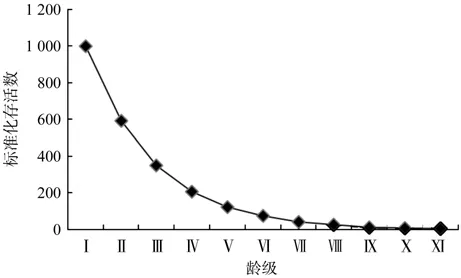
图2 青冈栎种群存活曲线Fig.2 Survivorship curve of Cyclobalanopsis glauca population
从图2中可以看出,曲线早期斜率较大,环境的自然选择强度大,因此死亡率较高,通过筛选的幼树仅为4.3%,并进入到VII龄段。在对存活曲线与II还是III型相符进行检验时,Loucks以及Hett分别通过指数方程式YNx和幂函数式YNo2种数学模型进行检验[19]。拟合得到的结果如下:
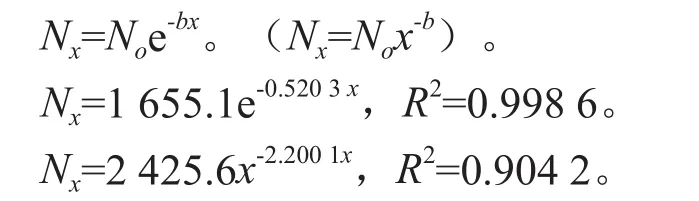
从两个模型的R2值可以看出,幂函数模型的R2值(0.904 2)比指数模型的R2值(0.998 6)要低,因此,青冈栎种群的存活曲线更趋于Deevey-Ⅱ型,呈对角线型,各年龄死亡数相同。

图3 青冈栎种群死亡率及消失率曲线Fig.3 Mortality and vanish rate curve of Cyclobalanopsis glauca population
结合图3可以发现,该种群的消失率、死亡率曲线变化近似一致。在VII龄级前,两者的变化幅度并不大,在Ⅷ龄级的时候,死亡率和消失率相对于之前有一个明显的上升,在Ⅷ龄级之后的发育过程中,消失率、死亡率持续下降,并于X龄级时,分别达到0.41以及0.22。
3.4 时间序列预测
时间序列预测是研究种群未来年龄结构和发展趋势的有效途径。青冈栎种群的时间序列预测以各龄级的基础数据,按照一次平均推移法预测青冈栎种群各龄级在未来10、20、30、40 a后各龄级个体数(见图4)。图4中,青冈栎种群各龄级数量都都呈现小幅度的增长趋势,但是老龄个体却逐渐增多。因此,由于幼龄个体的缺乏,若不采取人工干预措施,青冈栎种群未来必然趋于衰退趋势。
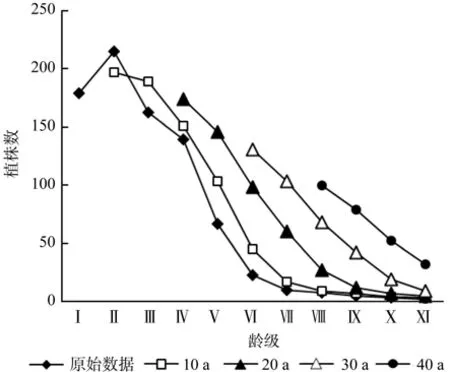
图4 青冈栎种群时间序列预测Fig.4 Time sequence analysis of Cyclobalanopsis glauca population
4 结论与讨论
通过对青冈栎种群的年龄结构分析,天然次生林中青冈栎种群的年龄结构呈非典型金字塔型,种群总体呈现增长趋势,但不具备显著增长性幅度,为稳定型种群,针对外界干扰存在较强的敏感度。
通过针对该种群消失率、死亡率、存活曲线、生命表的分析,最终发现:天然次生林中,该种群的存活曲线为Deevey-II型,在VIII龄级前,种群具有相似的死亡率,且死亡率较高,在Ⅷ龄级之后死亡率才有所下降。分析原因可能与林分郁闭度密切相关,青冈栎种群在Ⅰ—Ⅳ龄级个体数量虽多,但是郁闭度太大导致个体大量死亡,只有一部分能进入到Ⅴ—Ⅷ龄级,此阶段由于个体竞争压力较大加上郁闭度过高,使得死亡率偏高,而因IX到XI龄级,在年龄增长的同时,其逐步进入成熟龄阶段,针对环境的抗性、环境适应能力有所增强,死亡率开始逐渐下降。
从时间序列预测来看,在未来10、20、30、40 a后,青冈栎种群各龄级个体数都呈现小幅度的增长趋势,老龄个体持续增加,而幼龄株数不断下降,表明因可更新幼龄个体的欠缺,若不加强针对这一阶段的抚育管理,未来种群必将出现衰退问题。
青冈栎次生林种群结构及各阶段的动态特征,与其更新方式、更新能力、年龄结构和树高结构等因素密切相关,这些因素之间也存在相互影响。要详细分析青冈栎种群的生命表、以及各龄级的存活曲线和死亡率等数量特征,需要对同一生长发育条件下的各龄级的林分,设置固定样地进行观测,而本次研究所选取的五块样地,都处于中龄林和近熟林阶段,龄级比较集中,研究成果具有阶段性。因此,要形成完整的青冈栎次生林种群结构,后续研究还需增加幼龄林、成熟林林分,形成完整的龄级序列,并对这些林分进行长期的定期观测。
[1]李俊清.森林生态学[M].北京:高等教育出版社, 2006:186-192.
[2]王峥峰,安树青,朱学雷,等.热带森林乔木种群分布格局及其研究方法的比较[J].应用生态学报,1998,9(6):575-580.
[3]李肇晨.海南霸王岭陆均松种群结构及天然更新特征研究[D].海口:海南大学,2015.
[4]刘剑斌.闽北中亚热带青冈次生林高度结构分析[J].亚热带农业研究,2014,10(4):247 .
[5]曾思齐,龙时胜,肖化顺,等.南方地区青冈栎次生林种内与种间竞争研究[J].中南林业科技大学学报,2016,36(10):1-8.
[6]刘 畅,邓送求,方顺清,等.石栎、青冈栎和杉木混交林分的空间结构[J].林业科技开发,2011,25(1):35-39.
[7]梁 贵,项文化,赵仲辉,等.湘中丘陵区石栎—青冈栎常绿阔叶林土壤钾含量空间异质性及其影响因子研究[J].中南林业科技大学学报,2015,35(7):89-92;
[8]Weiner J.Neighbourhood interference amongst Pinus rigida individuals[J]. Journal of Ecology,1984, 72:183 -195.
[9]解婷婷,苏培玺,周紫鹃,等.荒漠绿洲过渡带沙拐枣种群结构及动态特征[J].生态学报,2014,34(15):4274.
[10]王电杰,温国胜,童在康,等.浙江省大鹿岛木麻黄种群静态生命表分析[J].内蒙古农业大学学报(自然科学版),2011,32(4): 146.
[11]李晓笑,王清春,崔国发,等.濒危植物梵净山冷杉野生种群结构及动态特征[J].西北植物学报,2011,31(7):1480.
[12]陈晓德.植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214-217.
[13]刘金福.格氏栲(Castanopsis kawakamiiHayata)种群结构与动态规律研究[D].北京:北京林业大学,2004 :41-45.
[14]江 洪.云杉种群生态学[M].北京: 中国林业出版社, 1992:8-40.
[15]申仕康,马海英,工跃华,等.濒危植物猪血术(Euryodendron excelsumH. T. Chang)自然种群结构及动态[J].生态学报,2008, 28(5): 2404-2412.
[16]郭其强,卢 杰,罗大庆,等.西藏色季拉山阳坡林线方枝柏种群结构特征研究[J].西北林学院学报,2010,25(2):l5-l8.
[17]卢 杰,郭其强,郑维列,等.藏东南高山松种群结构及动态特征[J].林业科学,2013,26 (8):158.
[18]郭 微,上官铁梁,王志明,等.灵空山油松种群年龄结构与动态分析[J].植物科学学报,2013,31(2):133.
[19]Silvertown J W. Introduction to Plant Population Ecology[M].London: Longman Group United Kingdom, 1982.
Structure and dynamic characteristics of Cyclobalanopsis glauca secondary forests
HU Man1, ZENG Siqi1, LONG Shisheng1, QIU Jianyou2, YU Junjian2
(1. Central South University of Forestry and Technology, Changsha 410004, Hunan, China 2. Malu Forestry Station of Anhua Forestry Bureau, Anhua 413506, Hunan, China)
Cyclobalanopsis glauca, one of the main species of the subtropical region in southern China, plays a vital role in the stability of the southern ecosystem. Through the method of sample survey and data statistics, the age structure of the population was analyzed,and the static life table of the population was compiled. The mortality, survival and disappearance rate curves were analyzed. By means of time series prediction and population quantitative dynamic method, The population structure and dynamic characteristics ofCyclobalanopsis glaucain natural secondary forest were revealed. The results showed that the dynamic change index ofCyclobalanopsis glaucawas 0.010 3, indicating that the population was stable. The survival curve was similar to Deevey-Ⅱ, and the mortality of each age group had similar mortality. Time series prediction The results showed that the number of young plants decreased gradually and the long-term stability of the population was stable. Therefore, the appropriate arti fi cial measures were to keep the natural regeneration and population stability ofCyclobalanopsis glaucapopulation in the future.
Cyclobalanopsis glauca; population structure; static life table; survival curve; time sequence analysis
S792.189
A
1673-923X(2017)11-0110-05
10.14067/j.cnki.1673-923x.2017.11.018
2017-06-28
国家林业公益性行业科研专项(201504301)
胡 满,硕士研究生
曾思齐,教授,博士生导师;E-mail:zengsiqi@csuft.edu.cn
胡 满,曾思齐,龙时胜,等.青冈栎次生林种群结构及动态特征[J].中南林业科技大学学报,2017, 37(11): 110-114.
[本文编校:吴 毅]
猜你喜欢
人类学学报(2023年5期)2023-04-29 19:50:21
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
河南农业大学学报(2020年2期)2020-05-22 02:56:30
山西林业科技(2019年1期)2019-05-27 12:24:52
运动(2017年1期)2017-09-03 10:39:20
绿色科技(2016年23期)2017-03-15 15:41:56
科教导刊·电子版(2016年25期)2016-11-16 10:24:35
江苏农业科学(2016年5期)2016-07-23 15:59:31
科技视界(2015年26期)2015-09-11 15:21:45
广西农学报(2015年4期)2015-09-11 02:55:46
