
烟草精氨酸脱羧酶的蛋白质特性分析
2017-11-17 06:53:24李雁斌靳双珍杨玉坤王德勋户艳霞范志勇苏家恩
湖南农业科学 2017年10期
李雁斌,靳双珍,杨玉坤,王德勋,户艳霞,范志勇,苏家恩
(1.云南省烟草公司大理州公司,云南 大理 671000;2.云南省烟草公司大理州公司剑川县分公司,云南 大理 671300)
烟草精氨酸脱羧酶的蛋白质特性分析
李雁斌1,靳双珍1,杨玉坤2,王德勋1,户艳霞1,范志勇1,苏家恩1
(1.云南省烟草公司大理州公司,云南 大理 671000;2.云南省烟草公司大理州公司剑川县分公司,云南 大理 671300)
为了研究参与烟草多胺合成的精氨酸脱羧酶的蛋白特性,运用生物信息学的方法分析了烟草精氨酸脱羧酶的理化特性、系统进化、氨基酸序列、保守区域和二级结构。结果表明:烟草精氨酸脱羧酶的等电点为4.99~6.78,氨基酸数为558~733,疏水性为-0.050~0.079,脂肪指数为88.04~92.08,不稳定指数为39.54~44.14;经进化树分析,烟草精氨酸脱羧酶分为2组,组1为NtCAD1、NtCAD2和NtCAD5,组2为NtCAD3和NtCAD4;经序列比对分析,NtCAD1、NtCAD2和NtCAD5的同源性较高,NtCAD3和NtCAD4的同源性较高,且组2均具有1个精氨酸脱羧酶保守序列和2个丙氨酸消旋酶/Ⅳ组脱羧酶保守序列;烟草精氨酸脱氢酶各成员的二级结构所占比例均为α-螺旋>无规则卷曲>β-折叠>β-转角。
烟草;精氨酸脱羧酶;蛋白特性;生物信息学
烟草是以收获烟叶组织为主的经济产物,烟叶在整个烟草植株上相对敏感的部位,易受到生物和非生物胁迫的危害,影响其产量与质量,进而损害烟农的经济收益。在生物或非生物胁迫条件下,多胺在植物防御反应中发挥了重要作用。例如,多胺具有调节离子平衡的作用,主要为K+、Na+、Ca+等离子[1];多胺可通过与核酸物质的结合,调节应激蛋白的翻译和转录,参与氨基酸序列空间结构的稳定[2]。从原始底物精氨酸到生成多胺需要经过多步酶促反应,精氨酸脱氢酶是多胺合成反应的第一个限速酶[3]。目前,关于精氨酸脱羧酶的研究,主要集中在康乃馨[4]、桃树[5]、水稻[6]、棉花[3]等植物上,对于烟草精氨酸脱羧酶的研究,尤其是分子水平的研究报道较为鲜见。笔者从烟草精氨酸脱羧酶的理化特性、系统进化、氨基酸序列、保守区域和二级结构等方面进行分析,以期揭示烟草精氨酸脱羧酶的蛋白特性,为探索该蛋白的结构功能奠定基础。
1 材料与方法
1.1 烟草精氨酸脱氢酶基因家族成员的获取
在Pfam[7]的数据库中获取具有精氨酸脱氢酶的序列文件(PF06314),且在NCBI[8]的蛋白数据库检索烟草精氨酸脱氢酶的氨基酸序列。以PF06314文件为参考,运用HMMER[9]程序筛选已获取的氨基酸序列,并除去重复序列,确定烟草精氨酸脱氢酶基因家族成员。
1.2 烟草精氨酸脱氢酶的蛋白质特性分析
采用protParam[10]的理化特性分析工具,分析烟草精氨酸脱羧酶各成员的等电点、氨基酸数、亲水性、脂肪指数和不稳定指数。利用ProbCons 1.12[11]在线服务,对烟草精氨酸脱氢酶的氨基酸序列进行分析,并通过PhyML 3.1[12]工具将解析后的数据进行聚类分析,其迭代次数为100,获得进化树。以Clustal[13]软件对精氨酸脱氢酶的氨基酸序列进行多序列比对,并根据MEME[14]工具分析保守区域在序列上的分布。通过SOPMA[15]分析蛋白质结构的组成及其比例。
2 结果与分析
2.1 精氨酸脱氢酶蛋白家族成员的筛选及理化特性
通过在线工具分析烟草精氨酸脱氢酶,结果表明如表1所示。烟草精氨酸脱氢酶具有5个家族成员,其等电点均小于7,以NtCAD4的等电点最大,为6.78,而NtCAD1的值最小,为4.99;各成员蛋白序列的氨基酸数目波动范围较大,处于558(NtCAD3)至733(NtCAD1)之间;NtCAD1、NtCAD2和NtCAD5的亲水性均小于0,表明这3个蛋白质具有亲水性,NtCAD3和NtCAD4的亲水性大于0,说明这2个蛋白具有疏水性;NtCAD3和NtCAD4的脂肪指数相对较大,具有较高耐热性;因NtCAD2和NtCAD5的不稳定指数小于40,属于稳定性蛋白,而NtCAD1、NtCAD3和NtCAD4的不稳定指数大于40。各成员蛋白质的氨基酸组成以丝氨酸(Ser)所占比例较大(除NtCAD1外)。综上所述,烟草精氨酸脱氢酶各成员的理化特性存在差异,但NtCAD1、NtCAD2和NtCAD5较为相似,NtCAD3和NtCAD4较为相似。
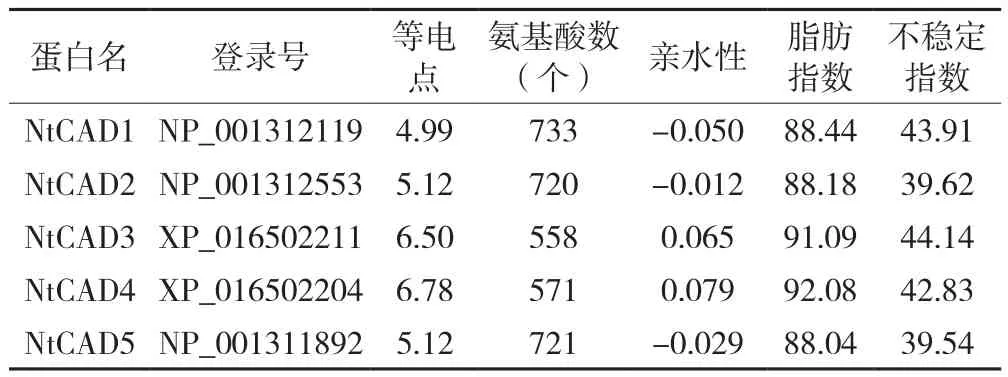
表1 烟草精氨酸脱氢酶蛋白家族成员的信息
2.2 精氨酸脱氢酶的系统进化树
以拟南芥的NP_195197和NP_179243蛋白序列,水稻的 XP_015631971、XP_015631972、XP_015631973、XP_015633833和XP_015643038蛋白序列与烟草精氨酸脱氢酶各成员的蛋白序列进行同源比较,通过PhyML进行聚类,结果如图1所示。依据进化树的排布,将烟草精氨酸脱氢酶成员划分为两组,1组的成员为NtCAD1、NtCAD2和NtCAD5,2组成员为NtCAD3和NtCAD4;依据各分支的亲缘关系,1组的烟草精氨酸脱氢酶与拟南芥的亲缘关系较近,可信度为64%;2组的成员与水稻的亲缘关系较近,可信度的100%。烟草精氨酸脱氢酶在单子叶植物和双子叶植物植物中均有分布,说明该蛋白的分化较早,分化时间先于单子叶和双子叶。
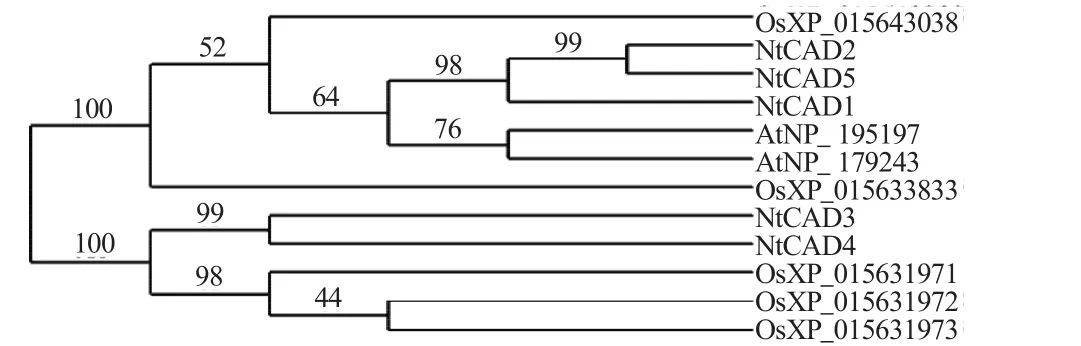
图1 烟草精氨酸脱氢酶的系统进化树
2.3 精氨酸脱氢酶的序列比对
将烟草精氨酸脱氢酶的5个成员进行序列比对,结果如图2所示。5个成员的蛋白序列中,既含有相同的氨基酸残基,又存在氨基酸残基数量和排列顺序的差异,以NtCAD1、NtCAD2和NtCAD5的同源性较高,NtCAD3和NtCAD4的同源性较高。通过对序列保守区域的进一步分析,每个序列均至少含有3种保守基序,分别为A、B和C;其中,NtCAD1、NtCAD2和NtCAD5蛋白的保守区域A位于68~107,保守区域B位于223~272,保守区域C位于490~540;而NtCAD2和NtCAD5的保守区域a位于601~648、保守区域b位于258~309,保守区域c位于355~409;保守区域A、C、a和c均具有丙氨酸消旋酶/Ⅳ组脱羧酶,保守区域B和b为精氨酸脱羧酶保守结构域。NtCAD1、NtCAD2和NtCAD5的丙氨酸消旋酶/Ⅳ组脱羧酶保守区域分别分布在蛋白序列的两端,即A保守域位于N-端,C保守域为C-端,而NtCAD3和NtCAD4 的保守域均位于C-端,且保守域a位于C-末端。
2.4 精氨酸脱氢酶的二级结构
蛋白二级结构的组成为蛋白三维结构及功能奠定基础,因此采用在线工具分别预测烟草精氨酸脱氢酶β-转角、无规则卷曲、α-螺旋和β-折叠的二级结构,结果如表2、图3所示。NtCAD1、NtCAD2和NtCAD5的N-端二级结构主要为无规则卷曲,NtCAD1的C-端以α-螺旋为主,NtCAD2和NtCAD5的C-端二级结构的排布无明显规律;NtCAD3和NtCAD4蛋白序列的N-端二级结构主要为α-螺旋,C-端以β-折叠为主。由表2可知,烟草精氨酸脱氢酶各成员的二级结构所占比例均为α-螺旋>无规则卷曲>β-折叠>β-转角,而α-螺旋多集中在蛋白序列中部。
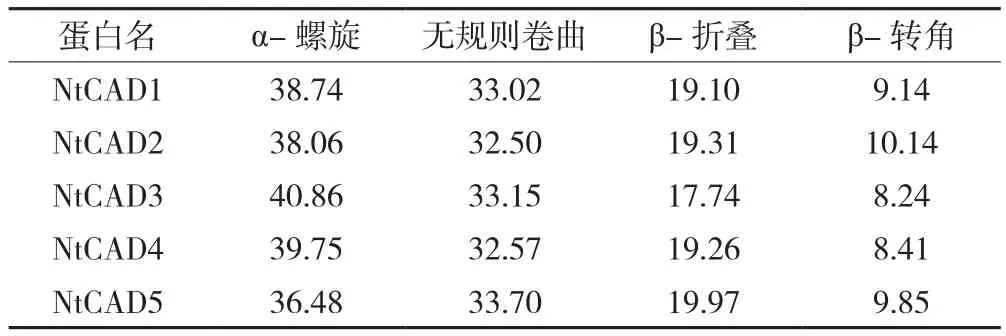
表2 烟草精氨酸脱氢酶各二级结构的比例 (%)

图2 烟草精氨酸脱氢酶的序列比对
3 结论与讨论
研究了烟草精氨酸脱氢酶的理化特性、进化树、氨斟酸序列、二级结构,结果显示,烟草精氨酸脱氢酶成员NtCAD1、NtCAD2和NtCAD5为一组,NtCAD3和NtCAD4为另外一组,尽管如此,各成员的功能和特异性表达仍存在差异[3]。经比对序列的保守区域分析,各成员蛋白序列除具有精氨酸脱羧酶保守结构域外,同时具有丙氨酸消旋酶/Ⅳ组脱羧酶的保守结构域。因此,该成员也属于磷酸吡哆醛(PLP)依赖型家族,参与丙氨酸空间结构的转化[16-17]。经二级结构分析,烟草精氨酸脱氢酶成员的二级结构既有共性也存在差异,使烟草精氨酸脱氢酶的功能具备多样性[18]。

图3 烟草精氨酸脱氢酶的二级结构
[1] Alcázar R,Tiburcio A F. Polyamines:molecules with regulatory functions in plant abiotic stress tolerance[J]. Planta,2010,231(6):1237-1249.
[2] Walters D. Resistance to plant pathogens:possible roles for free polyamines and polyamine catabolism[J]. New Phytologist,2010,159(1):109-115.
[3] 李亚栋,默辉娟,王省芬,等. 棉花精氨酸脱羧酶基因GhADC1克隆与表达分析[J]. 棉花学报,2013,25(4):291-299.
[4] Chang K S,Sun H L,Hwang S B,et al. Characterization and translational regulation of the arginine decarboxylase gene in carnation(Dianthus caryophyllus L.)[J]. Plant Journal for Cell & Molecular Biology,2000,24(1):45-56.
[5] Liu J H,Ban Y,Wen X P,et al. Molecular cloning and expression analysis of an arginine decarboxylase gene from peach(Prunus persica)[J]. Gene,2009,429(1-2):10-17.
[6] Akiyama T,Jin S. Molecular cloning and characterization of an arginine decarboxylase gene up-regulated by chilling stress in rice seedlings[J].Journal of Plant Physiology,2007,164(5):645-654.
[7] Finn R D,Coggill P,Eberhardt R Y,et al. The Pfam protein families database: towards a more sustainable future [J]. Nucleic Acids Research,2016,44(D1):279-285.
[8] Edgar R,Domrachev M,Lash A E. Gene Expression omnibus:NCBI gene expression and hybridization array data repository[J]. Nucleic Acids Research,2002,30(1):207-210.
[9] Marchin M,Kelly P T,Fang J. Tracker:continuous HMMER and BLAST searching[J]. Bioinformatics,2005,21(3):388-389.
[10] Liu S,Zhao Y,Zhang X,et al. Bioinformatic analysis of fanction and structure of putative nucleocapsid protein sequences of SARS-CoV[J].Progress in Biotechnology,2003,40(6):454.
[11] Do C B,Brudno M,Batzoglou S. PROBCONS:probabilistic consistency-based multiple alignment of amino acid sequences[J].Genome Research,2004,15(2):330-340.
[12] Guindon S. PHYML online—a web server for fast maximum likelihoodbased phylogenetic inference[J]. Nucleic Acids Research,2005,33(Web Server issue):557-559.
[13] Larkin M A,Blackshields G,Brown N P,et al. Clustal W and Clustal X version 2.0[J]. Bioinformatics,2007,23(21):2947-2948.
[14] Bailey T L,Elkan C. The value of prior knowledge in discovering motifs with MEME[J]. Proc Int Conf Intell Syst Mol Biol,1995,(Bailey):21-29.
[15] Geourjon C,Deléage G. SOPMA :significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments[J]. Computer Applications in the Biosciences Cabios,1995,11(6):681-684.
[16] Mo H,Pua E. Up-regulation of arginine decarboxylase gene expression and accumulation of polyamines in mustard(Brassica juncea)in response to stress[J]. Physiologia Plantarum,2002,114(3):439-449.
[17] Hao Y J,Kitashiba H,Honda C,et al. Expression of arginine decarboxylase and ornithine decarboxylase genes in apple cells and stressed shoots[J]. Journal of Experimental Botany,2005,56(414):1105-1115.
[18] Hummel I,Bourdais G,Gouesbet G,et al. Differential gene expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in Arabidopsis thaliana:characterization of transcriptional regulation during seed germination and seedling development[J]. New Phytologist,2004,163(3):519-531.
Analysis of Protein Characteristics of Arginine Decarboxylase in Tobacco
LI Yan-bin1,JIN Shuang-zhen1,YANG Yu-kun2,WANG De-xun1,HU Yan-xia1,FAN Zhi-yong1,SU Jia-en1
(1. Dali Tobacco Company, Yunnan Tobacco Corporation, Dali 671000, PRC; 2. Jianchuan Tobacco Branch Company,Dali Tobacco Company, Yunnan Tobacco Corporation, Dali 671300, PRC)
In order to test the protein properties of arginine decarboxylase involved in tobacco polyam ine synthesis, the bio-informatics method was used in this study to analyze the physicochemical properties, phylogenetics, amino acid sequence alignment, conservative region and secondary structure of arginine decarboxylase in tobacco. The results showed that, in tobacco arginine decarboxylase, the isoelectric point was 4.99 to 6.78, the number of amino acids was 558 to 733, the hydrophobicity was -0.050 to 0.079, the fat index was 88.04 to 92.08, and the instability index was 39.54 to 44.14; in analysis of the phylogenetic tree, tobacco arginine decarboxylase divided into 2 groups, group 1 was NtCAD1,NtCAD2 and NtCAD5, and group 2 was NtCAD3 and NtCAD4; the sequence homology among NtCAD1, NtCAD2 and NtCAD5 was higher, the sequence homology between NtCAD3 and NtCAD4 was higher, and the two groups had one arginine decarboxylase conserved sequence and two alanine racemase/IVgroup decarboxylase conservative sequences; and the proportion of secondary structure of each member of arginine dehydrogenase in tobacco was that of α-helix > that of random curl > that of β-sheet > that of β-turn.
tobacco; arginine decarboxylase; protein property; bio-informatics
Q557
A
1006-060X(2017)10-0001-05
10.16498/j.cnki.hnnykx.2017.010.001
2017-08-18
中国烟草总公司云南省公司资助项目(2015YN20;2016 YN10)
李雁斌(1987-),女,云南南涧县人,助理农艺师,主要从事烟叶生产工作。
苏家恩
(责任编辑:成 平)
猜你喜欢
天津市教科院学报(2021年5期)2021-11-10 07:32:40
生物学通报(2021年9期)2021-07-01 03:24:44
江苏农业科学(2016年8期)2017-02-15 19:54:11
中外医疗(2015年11期)2016-01-04 03:58:50
中国当代医药(2015年30期)2015-03-01 02:08:13
淮阴工学院学报(2014年5期)2014-09-10 09:15:56
生物质化学工程(2014年4期)2014-07-05 16:20:28
食品工业科技(2014年13期)2014-03-11 18:16:53
食品科学(2013年24期)2013-03-11 18:30:35
河南医学研究(2012年3期)2012-08-15 00:48:04
