
以防治油菜菌核病为目标的核盘菌乙酰乳酸合酶抑制剂的筛选
2017-11-14 09:46温雪玮陈泱泱武斌胡东维梁五生
浙江大学学报(农业与生命科学版) 2017年5期
温雪玮,陈泱泱,武斌,胡东维,梁五生
以防治油菜菌核病为目标的核盘菌乙酰乳酸合酶抑制剂的筛选
温雪玮,陈泱泱,武斌,胡东维,梁五生*1
(浙江大学农业与生物技术学院生物技术研究所/农业部作物病虫分子生物学重点实验室,杭州310058)
为开发新的油菜菌核病防治药剂,选取7种乙酰乳酸合酶(acetolactate synthase,ALS)潜在抑制剂(氯嘧磺隆、苄嘧磺隆、氯磺隆、甲嘧磺隆、咪唑乙烟酸、灭草喹和磺胺噻唑)作为3类ALS酶抑制剂(磺酰脲类、咪唑啉酮类、磺胺噻唑类)的代表,对其防治油菜菌核病的性能进行测试。结果显示,1.0 mg/L灭草喹、磺胺噻唑、氯嘧磺隆、苄嘧磺隆和氯磺隆对核盘菌ALS酶活性有较强的抑制作用,而相同剂量的甲嘧磺隆的抑制作用微弱,咪唑乙烟酸无抑制作用。相应地,半最大效应浓度(half-maximal effective concentration,EC50)值显示,核盘菌的菌丝生长对灭草喹、磺胺噻唑、氯嘧磺隆、苄嘧磺隆和氯磺隆的抑制作用较敏感,而对甲嘧磺隆和咪唑乙烟酸的敏感性较弱。氯嘧磺隆、苄嘧磺隆、氯磺隆、灭草喹和甲嘧磺隆对油菜叶片ALS酶活性有强烈的抑制作用,咪唑乙烟酸和磺胺噻唑的抑制作用明显比上述5种抑制剂弱。氯嘧磺隆、苄嘧磺隆、氯磺隆、灭草喹、甲嘧磺隆和咪唑乙烟酸处理对油菜幼苗生长发育有明显的负面影响,主要表现为生长速率下降,植株明显比对照矮小,叶片变小,叶色变黄。磺胺噻唑处理对油菜幼苗生长发育的负面影响明显比上述6种抑制剂弱。选择对核盘菌ALS酶活性和菌落生长的抑制作用都较强、而对油菜ALS酶活性的抑制作用和对油菜幼苗生长的负面影响最弱的磺胺噻唑进行抗核盘菌侵染试验。结果显示,磺胺噻唑处理显著地降低了核盘菌对油菜叶片的侵染程度,对油菜菌核病产生了较明显的防治效果。上述研究结果表明,磺胺噻唑可作为分子结构基础来进一步开发以核盘菌ALS酶为作用靶标的油菜菌核病防治剂。
乙酰乳酸合酶;菌核病;核盘菌;油菜;磺胺噻唑
核盘菌(Sclerotinia sclerotiorum)是一种可对农业产生严重负面影响的植物病原真菌。该真菌可侵染超过400种植物,其中包括不少重要的农作物和蔬菜[1-4]。油菜是我国主要的油料作物,也是世界范围内重要的油料作物。核盘菌是油菜的主要病害之一,可导致油菜菌核病(rapeseed Sclerotinia disease)。该病在我国一般年份的发病率为10%~30%,导致5%~30%的产量损失;流行年份在一些严重田块的发病率可超过80%[5-6]。
由于目前尚无核盘菌高抗油菜品种,油菜种植者仍主要依赖杀菌剂来防治该病害。多年来我国一直使用以多菌灵和甲基托布津为主的苯并咪唑类杀菌剂来防治油菜菌核病[7]。由于长期、反复地使用多菌灵,目前已在许多地区监测到抗多菌灵的核盘菌菌株[7-9]。因此,有必要指导油菜种植者更加科学地使用已有杀菌剂,另外还需要不断开发新的油菜菌核病防治药剂。
乙酰乳酸合酶(acetolactate synthase,ALS),又称乙酰羟基酸合酶(acetohydroxyacid synthase),是植物和微生物细胞中催化支链氨基酸生物合成过程中第一步反应的酶[10-12]。通过抑制植物细胞中的ALS酶活性可以破坏支链氨基酸生物合成,阻断蛋白质合成,干扰细胞分裂,结果影响植物生长,甚至可能导致植物死亡[12-13]。由于动物细胞内不含ALS酶,在田间施用ALS酶抑制剂作为除草剂对人、畜等动物安全,故此酶是除草剂研发中的一种重要靶标[14-15]。近年来,以人体病原细菌和真菌的ALS酶作为靶标开发新的人用抗生素和杀菌剂的研究报道逐渐增多[16-21],但尝试利用ALS酶抑制剂来防治植物真菌病害的研究目前只见到1篇文献报道[22]。
我们从NCBI数据库中搜取核盘菌ALS酶的氨基酸序列(XP_001592310.1)和油菜ALS酶的氨基酸序列(CAA77613.1),通过序列比对后发现两者的同源性只有36.48%(图1)。如此低的同源性暗示理论上有可能筛选到对核盘菌ALS酶作用较强、但对油菜ALS酶作用较弱的抑制剂,用于防治油菜菌核病。本文报道了我们对这一可能性进行探索的试验结果。

图1 核盘菌和油菜的ALS酶氨基酸序列比较Fig.1 Comparison of the amino acid sequences of acetolactate synthases of Sclerotinia sclerotiorum and rapeseed
1 材料与方法
1.1 试验材料
核盘菌分离物(7-3)分离自油菜种植地,之前已用于进行其他一些研究[23-25]。油菜品种为浩油11。
核盘菌的培养按照之前报道的方法进行[23-25]。在用固体平板培养时,将马铃薯葡萄糖琼脂(potato dextrose agar,PDA)培养基倒至直径为9 cm的培养皿中制成PDA平板,将核盘菌菌核接种于PDA平板表面中心位置,然后置于暗处22℃下培养。继代培养时用灭过菌的5 mm内径打孔器从PDA平板上培养的核盘菌菌落外缘打取菌碟接种于新的PDA平板上,使菌丝面与培养基接触,置于暗处22℃下培养。在用液体培养基培养时,如上用打孔器从PDA平板上培养的核盘菌菌落外缘打取菌碟接种到用三角瓶盛装的马铃薯葡萄糖(potato dextrose,PD)培养基中,然后置于暗处22℃下振荡培养。
油菜苗的培养按照常规方法进行。将油菜种子用水冲洗干净后在蒸馏水中浸泡1 d,然后置于湿润的滤纸上,放入人工气候箱中光照萌发(22℃,光/暗=16 h/8 h)。用塑料杯(开口直径为6.8 cm)分装适量的营养土,将种子萌发后获得的油菜幼苗植于营养土中,每杯土栽1株苗,将幼苗放入人工气候箱中光照培养(22 ℃,光/暗=16 h/8 h)。
1.2 核盘菌菌丝ALS酶的提取
用PD培养基培养核盘菌6 d,取菌丝用2层纱布过滤并尽量挤去培养基,称取适量菌丝,置于用液氮预冷过的研钵中,依次加入适量石英砂和液氮,将菌丝研磨成粉末,然后按菌丝鲜质量∶缓冲液体积=1∶2的比例加入ALS酶提取缓冲液[0.1 mol/L磷酸钾缓冲液(pH 7.5),1.0 mmol/L焦磷酸氯化硫胺素,0.1 mmol/L FAD,3.0 mmol/L MgCl2],充分混匀,冰浴20 min后用2层纱布过滤,将滤液在4℃、1.2万r/min条件下离心20 min,收集上清液,即核盘菌菌丝ALS粗酶液。按参考文献[26]的方法测定提取的粗酶液中蛋白质浓度。
1.3 油菜叶片ALS酶的提取
从油菜幼苗上剪取适量叶片,用水冲洗干净后吸去表面水分,称鲜质量,置于用液氮预冷过的研钵中,加入适量石英砂,再加液氮研磨为粉末,按叶片鲜质量∶缓冲液体积=1∶5的比例加入ALS酶提取缓冲液(组成同核盘菌菌丝ALS酶的提取),充分混匀,冰浴20 min后用2层纱布过滤,将滤液在4℃、1.2万r/min条件下离心20 min,收集上清液,即油菜叶片ALS粗酶液。按参考文献[26]的方法测定提取的粗酶液中蛋白质浓度。
1.4 ALS酶潜在抑制剂对核盘菌和油菜ALS酶的抑制效力测定
研究表明,不同物种的ALS酶之间可能有差异,抑制剂的抑制效果有物种特异性[27-32]。在试验前因不明确本文测试的ALS酶抑制剂对核盘菌ALS酶及后续油菜ALS酶的抑制效力,所以将它们称为“核盘菌和油菜ALS酶的潜在抑制剂”。经过调研文献,选取以下7种ALS酶潜在抑制剂进行试验:氯嘧磺隆、苄嘧磺隆、氯磺隆、甲嘧磺隆、咪唑乙烟酸、灭草喹和磺胺噻唑。其中:氯嘧磺隆、苄嘧磺隆、氯磺隆和甲嘧磺隆作为磺酰脲类ALS酶抑制剂的代表,咪唑乙烟酸和灭草喹作为咪唑啉酮类ALS酶抑制剂的代表,磺胺噻唑作为磺胺噻唑类ALS酶抑制剂的代表。以上7种ALS酶潜在抑制剂均购自上海阿拉丁生化科技股份有限公司。
[33]报道的方法测定ALS酶的活性并略作改进,即用乙酰乳酸脱羧酶取代硫酸进行脱羧反应。反应分3类:1)对照反应管。取试管,加入1 mL前面提取的粗酶液和50 μL 30%硫酸,摇匀,再加入1 mL反应缓冲液[50 mmol/L磷酸钾缓冲液(pH 7.0),200 mmol/L丙酮酸钠,0.5 mmol/L焦磷酸氯化硫胺素,0.03 mmol/L FAD,2.0 mmol/L MgCl2],以及10 μL稀释100倍的乙酰乳酸脱羧酶溶液,摇匀,37℃水浴反应1 h。2)正常反应管。取试管,加入1 mL粗酶液,1 mL反应缓冲液,10 μL稀释100倍的乙酰乳酸脱羧酶溶液和50 μL蒸馏水,摇匀,37℃水浴反应1 h。3)加抑制剂反应管。取试管,加入1 mL粗酶液,适量ALS酶潜在抑制剂溶液,1 mL反应缓冲液,10 μL稀释100倍的乙酰乳酸脱羧酶溶液,加适量蒸馏水使反应液体积一致,并使抑制剂终质量浓度为1 mg/L,摇匀,37℃水浴反应1 h。上述3类反应管水浴反应后每管都依次加入0.5%肌酸500 μL、9% α-萘酚溶液500 μL(用4 mol/L NaOH配制),60℃水浴加热15 min,室温下放置15 min,5 000 r/min离心5 min,收取上清液。将从对照反应管得到的上清液用来调零,测定从正常反应管和加抑制剂反应管得到的上清液在530 nm处的吸光度值D(530 nm)。通过D(530 nm)、反应液中的蛋白质量和反应时间计算酶活性及抑制剂对ALS酶活性的抑制率。以U表示酶活性单位,1 U指每毫克蛋白质每小时使反应液在530 nm处吸光度的变化值为1.0。抑制剂对ALS酶活性的抑制率=(正常反应管的酶活性-加抑制剂反应管的酶活性)/正常反应管的酶活性×100%。测定试验重复3次,每次试验中每类反应管都至少平行测定3份。
1.5 核盘菌对ALS酶潜在抑制剂的敏感性测定
将抑制剂分别溶解后取适量加入PDA培养基中,倒至9 cm直径培养皿中,每种ALS酶潜在抑制剂都制成含不同浓度抑制剂的敏感性测定PDA平板,用灭过菌的5 mm内径打孔器从继代培养PDA平板上培养2 d的核盘菌菌落外缘打取菌碟,接种于敏感性测定PDA平板中心,然后置于暗处22℃下培养,观察菌丝生长情况。每种浓度至少重复3份。当无ALS酶潜在抑制剂PDA平板(对照平板)上的菌落快长满平板时,从2个互相垂直的方向测量所有PDA平板上的菌落直径,所得数据用于计算EC50值(抑制50%菌丝生长的有效浓度)。EC50值的计算按参考文献[22-23]的方法进行。
1.6 ALS酶潜在抑制剂对油菜幼苗生长发育的影响试验
培养油菜幼苗至适当大小(约3~5叶期),用镊子夹取脱脂棉吸取ALS酶潜在抑制剂溶液(1.0 mg/L),轻柔涂抹油菜幼苗第1片真叶的上表面,将幼苗置于人工气候箱中,22℃下光照培养。每天观察记录幼苗的生长发育情况。试验重复3次,在每次试验中每种处理至少重复5株。
1.7 ALS酶潜在抑制剂对核盘菌侵染油菜的防治效果测定
按参考文献[1]的方法进行核盘菌侵染油菜离体叶片试验。取油菜叶片,先后用自来水和蒸馏水冲洗干净。取直径11 cm的玻璃培养皿,铺上2层粗滤纸,加入适量无菌水使滤纸充分润湿。然后将叶片铺于湿润的滤纸上,用医用无菌脱脂棉吸取无菌水包裹叶柄保湿,在非叶脉位置用针尖刺一小孔,用灭过菌的5 mm内径打孔器从继代培养PDA平板上培养2 d的核盘菌菌落外缘打取菌碟,接种于油菜离体叶片的小孔上,使菌丝面与叶片接触。将接种了菌碟的叶片分成2组:一组用1.0 mg/L ALS酶潜在抑制剂溶液进行叶面喷雾处理,使叶面均匀分布细小液滴;另一组用无菌蒸馏水对叶面进行类似的喷雾处理,作为有接菌但无抑制剂处理的对照。另外取一组叶片作为空白(无菌)接种对照,即用灭菌的5 mm内径打孔器从PDA无菌平板上打取无菌琼脂圆块,接种于油菜离体叶片的小孔上。将上述放有试验叶片的培养皿盖上玻璃盖后置于人工气候箱中,22℃下光照培养,1 d后拍照,并测量最宽处和最窄处的长度来计算病斑幅度[病斑幅度=(最宽处长度+最窄处长度)/2]。试验重复3次,在每次试验中每组处理或对照至少接种10片油菜叶。
2 结果与分析
2.1 ALS酶潜在抑制剂对核盘菌ALS酶活性的抑制作用
检测结果(图2)表明:当抑制剂质量浓度为1.0 mg/L时,氯嘧磺隆、苄嘧磺隆、氯磺隆、甲嘧磺隆、灭草喹和磺胺噻唑这6种抑制剂均对核盘菌ALS酶活性有一定的抑制作用,且以灭草喹的抑制作用最强,甲嘧磺隆的抑制作用最弱,仅有微弱的抑制作用;而1.0 mg/L咪唑乙烟酸对核盘菌ALS酶活性无抑制作用。
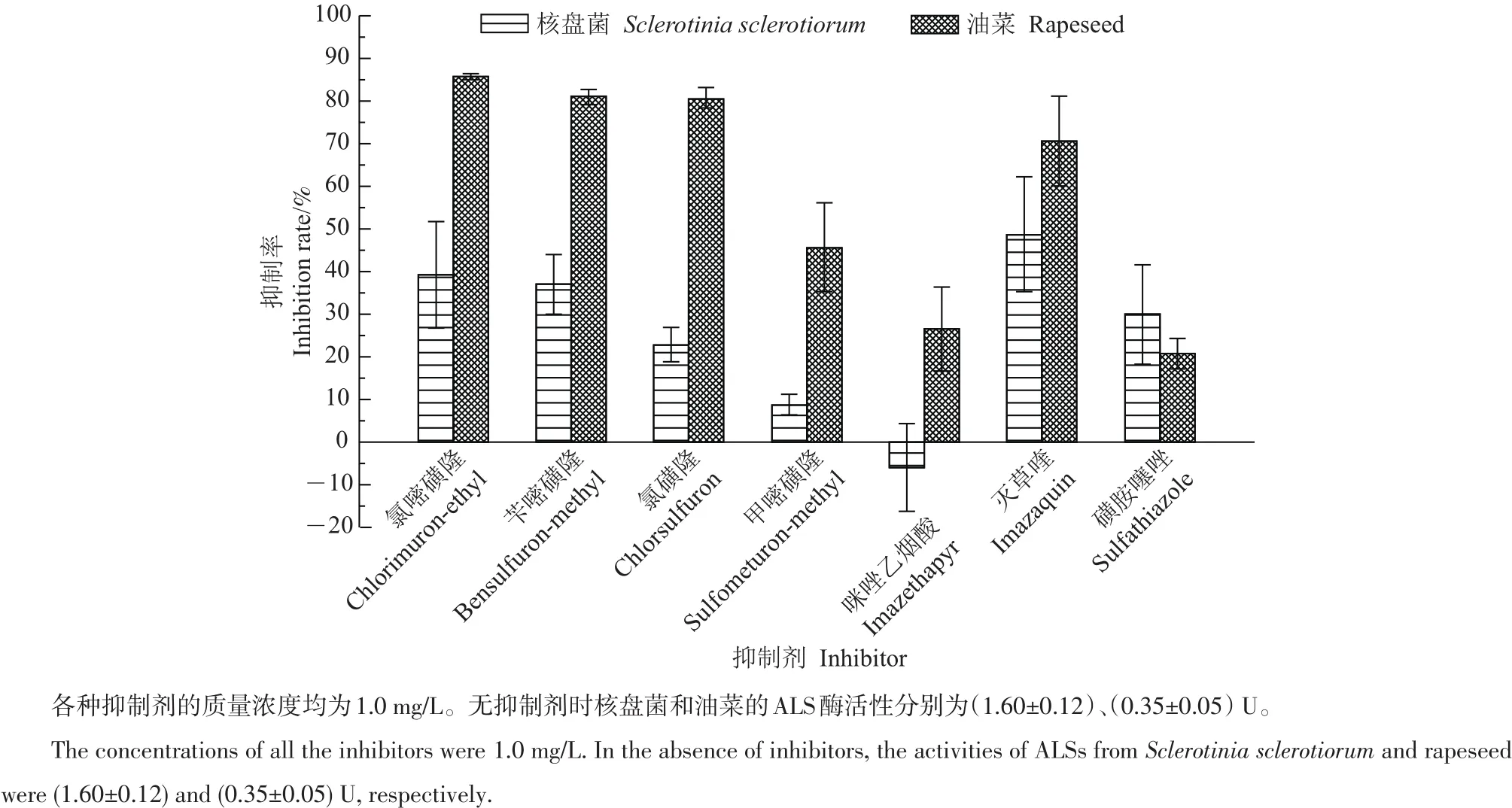
图2 7种ALS酶潜在抑制剂对核盘菌和油菜ALS酶活性的抑制作用Fig.2 Inhibitory effects of seven potential acetolactate synthase(ALS)inhibitors on the activity of ALS from Sclerotinia sclerotiorum and rapeseed
2.2 ALS酶潜在抑制剂对核盘菌菌落生长的抑制作用
为了探明本试验7种ALS酶潜在抑制剂对核盘菌菌丝生长的影响,利用PDA平板测试了这些抑制剂在不同质量浓度条件下对核盘菌菌落生长的影响。结果表明,灭草喹和磺胺噻唑在0.1 mg/L时即有明显的抑制作用,氯嘧磺隆、苄嘧磺隆和氯磺隆自1.0 mg/L起有明显的抑制作用,而甲嘧磺隆和咪唑乙烟酸在50 mg/L时的抑制作用仍很弱(图3)。此外,对7种ALS酶潜在抑制剂对核盘菌的EC50值测算结果表明:灭草喹、磺胺噻唑、氯嘧磺隆、苄嘧磺隆、氯磺隆对核盘菌的EC50值小于10 mg/L,表明核盘菌对这5种抑制剂相对较敏感;而甲嘧磺隆和咪唑乙烟酸对核盘菌的EC50值都大于50 mg/L,表明核盘菌对这2种抑制剂相对不太敏感(表1)。这与它们对核盘菌ALS酶活性抑制作用强弱的变化趋势一致。说明核盘菌对这些抑制剂的敏感性强弱与它们抑制核盘菌ALS酶活性的作用强弱相关。
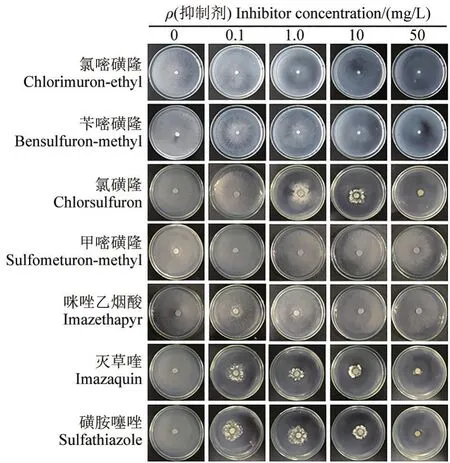
图3 7种ALS酶潜在抑制剂对PDA平板上核盘菌菌落生长的影响Fig.3 Effects of seven potential acetolactate synthase inhibitors on the growth of Sclerotinia sclerotiorum colony on potato dextrose agar plates
2.3 ALS酶潜在抑制剂对油菜ALS酶活性的抑制作用
检测结果表明,7种ALS酶潜在抑制剂对油菜叶片ALS酶活性均有一定的抑制作用,其中氯嘧磺隆、苄嘧磺隆、氯磺隆、灭草喹和甲嘧磺隆这5种抑制剂的抑制作用较强,而咪唑乙烟酸和磺胺噻唑的抑制作用较弱,尤其是磺胺噻唑的抑制作用最弱(图2)。比较它们对油菜和核盘菌ALS酶的抑制作用可见,除磺胺噻唑外,其余6种ALS酶潜在抑制剂对油菜ALS酶的抑制作用都明显强于对核盘菌ALS酶的抑制作用。

表1 7种ALS酶潜在抑制剂对核盘菌的EC50值Table 1 EC50values of seven potential acetolactate synthase inhibitors to Sclerotinia sclerotiorum
2.4 ALS酶潜在抑制剂对油菜幼苗生长发育的影响
试验结果(图4)表明:氯嘧磺隆、苄嘧磺隆、氯磺隆、灭草喹、甲嘧磺隆和咪唑乙烟酸这6种ALS酶潜在抑制剂在1.0 mg/L时即对油菜幼苗的生长发育有明显的负面影响,主要表现为生长速率下降,植株明显比对照矮小,叶片变小,叶色变黄;而磺胺噻唑对油菜幼苗的生长发育虽然也有一定程度的负面影响,但其影响程度明显比上述6种抑制剂弱。比较各抑制剂对油菜ALS酶活性的抑制作用强弱和对油菜幼苗生长发育的影响程度可以看出,各抑制剂对油菜幼苗生长发育的影响程度与抑制油菜ALS酶活性的作用强弱基本相关。

图4 7种ALS酶潜在抑制剂对油菜幼苗生长发育的影响Fig.4 Effects of seven potential acetolactate synthase inhibitors on the growth and development of rapeseed seedlings
2.5 ALS酶潜在抑制剂对核盘菌侵染油菜叶片的影响

图5 磺胺噻唑处理对核盘菌侵染油菜叶片的影响Fig.5 Effect of sulfathiazole treatment on the infection of Sclerotinia sclerotiorum to rapeseed leaves
油菜菌核病防治剂需要对核盘菌有较好的灭杀或抑制作用,但不能影响油菜的生长发育。上述结果显示,在7种ALS酶潜在抑制剂中,核盘菌对灭草喹、磺胺噻唑、氯嘧磺隆、苄嘧磺隆和氯磺隆敏感,其中磺胺噻唑对油菜ALS酶活性的抑制作用和对油菜幼苗生长发育的负面影响最弱,因此,磺胺噻唑用于防治油菜菌核病的潜力最大。为此,我们测试了磺胺噻唑处理对核盘菌侵染油菜叶片的影响。结果(图5)显示:核盘菌菌丝侵染油菜叶片的能力非常强,1 d内即可诱发明显的病斑;而在油菜叶片接种核盘菌后喷施磺胺噻唑(1 mg/L),可显著降低核盘菌侵染油菜叶片所诱发的病斑幅度。这表明磺胺噻唑处理对核盘菌侵染油菜的进程有明显的抑制作用。因此,磺胺噻唑处理可对油菜菌核病产生较明显的防治效果。
3 讨论与结论
动物细胞内不含ALS酶。迄今关于ALS酶的研究绝大多数是以植物或细菌为研究对象,另外有极少量以真菌为材料。研究植物ALS酶主要是为了研发以ALS酶为靶标的除草剂[15]。细菌ALS酶研究主要以大肠杆菌为材料,研究ALS酶的活性调控机制[29]。近年来以人体病原细菌ALS酶为靶标开发人用抗生素逐渐引起关注。例如结核分枝杆菌(Mycobacterium tuberculosis)是一种可引发人体结核病(如肺结核)的病原细菌。JUNG等[21]最近从6 800种化合物中筛选到15种对结核分枝杆菌ALS酶具有较强抑制作用的化合物,它们在20 μmol/L时对结核分枝杆菌ALS酶活性的抑制率即大于80%,其中5种三唑并嘧啶类化合物的抑制效果尤其突出,其半最大抑制效应浓度(half-maximal inhibitory concentration,IC50)值为 0.4~1.24 μmol/L,更重要的是它们对一些从临床病样中分离得到的具有广谱耐药性的结核分枝杆菌株系也有显著的抑制作用,其MIC50值(50%minimal inhibitory concentration,50%最低抑菌质量浓度)只有0.2~0.8 mg/L。这一质量浓度水平与常用的抗结核病药物异烟肼和利福平的MIC50值浓度水平相当,因此上述筛选得到的ALS酶抑制剂有望用于开发以ALS酶为作用靶标的抗结核病药物。迄今为止,对真菌ALS酶的研究报道仍非常少,但与细菌ALS酶研究发展趋势类似,以人体病原真菌ALS酶为靶标开发人用杀菌剂近年来逐渐引起关注。比如白色念珠菌(Candida albicans)是一种人体病原真菌,可侵犯人体的皮肤、黏膜而引发炎症,甚至可导致内脏或全身感染。LEE等[16]研究证实了几种磺酰脲类化合物对白色念珠菌的ALS酶和菌落生长有抑制作用,其中乙基-2-{[(4-碘-6-甲氧基嘧啶-2-基)氨甲酰]氨磺酰}苯甲酸酯的抑制作用最强,其MIC90值(90%minimal inhibitory concentration,90%最低抑菌质量浓度)为0.7 mg/L。
涉及植物病原真菌ALS酶的研究目前只见到2篇文献报道[22,34]。DU等[34]报道了稻瘟病菌(Magnaporthe oryzae)的ALS酶为稻瘟病菌实现其致病性所必需。叶朝燕等[22]测试了14种嘧啶氧苄胺类化合物对镰刀菌(Fusarium graminearum)ALS酶活性及菌落生长的抑制效果,这也是涉及利用ALS酶抑制剂来防治植物真菌病害的唯一研究报道。这一报道以及上文所述以ALS酶为靶标的人用抗生素和杀菌剂的研究进展启示我们利用ALS酶抑制剂来防治油菜菌核病可能具有较大潜力。由于植物细胞内含有ALS酶,因此研发以ALS酶为靶标的植物真菌病害防治剂需要确保不影响植物的ALS酶。目前研究已证实,ALS酶与其抑制剂之间存在紧密的结构对应关系[27-32],ALS酶一个氨基酸的变化即可能改变其对某种抑制剂的敏感性。比如LEE等[28]报道,诱导大麦(Hordeum vulgare L.)ALS酶编码基因的一个碱基发生替换,导致ALS酶的653位丝氨酸突变成天门冬酰胺,结果大麦从对咪唑啉酮类除草剂甲氧咪草烟敏感变为不敏感;BROSNAN等[31]报道早熟禾(Poa annua L.)ALS酶编码基因的2个碱基发生替换,导致ALS酶的205位丙氨酸突变成苯丙氨酸,结果早熟禾产生对多类除草剂(咪唑啉酮类、磺酰脲类、三唑并嘧啶类、磺酰基氨基-碳酰-三唑啉酮类、嘧啶硫苯甲酸类)的抗性;HUANG等[32]报道反枝苋(Amaranthus retroflexus L.)ALS酶的376位天冬氨酸突变成谷氨酸,反枝苋对除草剂咪唑乙烟酸从敏感变为不敏感。比对核盘菌ALS酶和油菜ALS酶的氨基酸序列后发现,两者的同源性只有36.48%。因此,从理论上有可能筛选到对油菜ALS酶作用较弱、但对核盘菌ALS酶作用较强的抑制剂,用于防治油菜菌核病。本研究对7种ALS酶潜在抑制剂(氯嘧磺隆、苄嘧磺隆、氯磺隆、甲嘧磺隆、咪唑乙烟酸、灭草喹和磺胺噻唑)防治油菜菌核病的可能性进行了试验。结果发现,灭草喹、磺胺噻唑、氯嘧磺隆、苄嘧磺隆和氯磺隆对核盘菌ALS酶活性和核盘菌菌落生长都具有较好的抑制作用,而这5种抑制剂中磺胺噻唑对油菜ALS酶活性的抑制作用和对油菜幼苗生长发育的影响都最弱。抗核盘菌侵染试验的结果显示,喷施磺胺噻唑显著地降低了核盘菌对油菜叶片的侵染程度。这些结果表明,磺胺噻唑用于防治油菜菌核病具有较大的潜力。由于磺胺噻唑目前已被作为医用抗菌药物,用于防治肺炎球菌、脑膜炎双球菌、淋球菌和溶血性链球菌等人体病原菌的感染。基于农用和医用抗菌素分开的原则,磺胺噻唑本身不宜作为油菜菌核病的防治剂使用,但本试验结果表明,磺胺噻唑可作为分子结构基础来进一步开发以核盘菌ALS酶为作用靶标的油菜菌核病防治剂。
在对核盘菌ALS酶或油菜ALS酶的抑制效力方面,本试验结果显示,2种咪唑啉酮类ALS酶潜在抑制剂(咪唑乙烟酸和灭草喹)的抑制效力差异显著,4种磺酰脲类ALS酶潜在抑制剂(氯嘧磺隆、苄嘧磺隆、氯磺隆和甲嘧磺隆)的抑制效力也具有一定差异。这说明ALS酶抑制剂的侧链基团对其抑制效力具有重要影响,也再次证明ALS酶与其抑制剂之间存在紧密的结构对应关系。目前对这一结构对应关系已有一些研究[25-30],但仍了解不够,因此,对其继续进行系统深入的研究将有助于研发更多更佳的以ALS酶为靶标的油菜菌核病防治药剂,以及其他植物真菌病害的防治药剂。
参考文献(References):
[1] WANG C G,YAO J,DU X Z,et al.The Arabidopsis mediator complex subunit16 is a key component of basal resistance against the necrotrophic fungal pathogen Sclerotinia sclerotiorum.Plant Physiology,2015,169(1):856-872.
[2] BOLAND G J,HALL R.Index of plant hosts of Sclerotinia sclerotiorum.Canadian Journal of Plant Pathology,1994,16(2):93-108.
[3] BARDIN S D,HUANG H C.Research on biology and control of Sclerotinia diseases in Canada.Canadian Journal of Plant Pathology,2001,23(1):88-98.
[4] YOUNG C S,CLARKSON J P,SMITH J A,et al.Environmental conditions influencing Sclerotinia sclerotiorum infection and disease development in lettuce.Plant Pathology,2004,53(4):387-397.
[5] 汪雷,刘瑶,丁一娟,等.油菜菌核病研究进展.西北农林科技大学学报(自然科学版),2015,43(10):85-93.WANG L,LIU Y,DING Y J,et al.Advance in Sclerotinia stem rot of rapeseed.Journal of Northwest A&F University(Natural Science Edition),2015,43(10):85-93.(in Chinese with English abstract)
[6] LI G Q,HUANG H C,MIAO H J,et al.Biological control of sclerotinia diseases of rapeseed by aerial applications of the mycoparasite Coniothyrium minitans.European Journal of Plant Pathology,2006,114(4):345-355.
[7] 周锋,王彦芬,张小磊,等.油菜菌核病病原菌对多菌灵抗药性的研究进展.湖南农业科学,2012(17):82-84.ZHOU F,WANG Y F,ZHANG X L,et al.Advances in resistance of Sclerotinia sclertiorum to carbendazim.Hunan Agricultural Sciences,2012(17):82-84.(in Chinese with English abstract)
[8] 杨敬辉,潘以楼,朱桂梅,等.油菜菌核病菌对多菌灵和乙霉威的抗药性机理.植物保护学报,2004,31(1):74-78.YANG J H,PAN Y L,ZHU G M,et al.Mechanism of resistance of Sclerotinia sclertiorum to carbendazim and diethofencarb.Acta Phytophylacica Sinica,2004,31(1):74-78.(in Chinese with English abstract)
[9] 李伟,李伟,周益军,等.江苏省油菜菌核病菌对多菌灵的敏感性.中国油料作物学报,2007,29(1):63-68.LI W,LI W,ZHOU Y J,et al.Sensitivity of Sclerotinia sclerotiorum isolates to carbendazim in Jiangsu Province.Chinese Journal of Oil Crop Sciences,2007,29(1):63-68.(in Chinese with English abstract)
[10]MCCOURT J A,DUGGLEBY R G.Acetohydroxyacid synthase and its role in the biosynthetic pathway for branched-chain amino acids.Amino Acids,2006,31(2):173-210.
[11]LIU Y D,LI Y Y,WANG X Y.Acetohydroxyacid synthases:Evolution,structure,and function.Applied Microbiology and Biotechnology,2016,100(20):8633-8649.
[12]GALILI G,AMIR R,FERNIE A R.The regulation of essential amino acid synthesis and accumulation in plants.Annual Review of Plant Biology,2016,67:153-178.
[13]RAY T B.Site of action of chlorsulfuron:Inhibition of valine and isoleucine biosynthesis in plants.Plant Physiology,1984,75(3):827-831.
[14]MCCOURT J A,PANG S S,KING-SCOTT J,et al.Herbicidebinding sites revealed in the structure of plant acetohydroxyacid synthase.Proceedings of the National Academy of Sciences of the USA,2006,103(3):569-573.
[15]POWLES S B,YU Q.Evolution in action:Plants resistant to herbicides.Annual Review of Plant Biology,2010,61:317-347.
[16]LEE Y T,CUI C J,CHOW E W L,et al.Sulfonylureas have antifungal activity and are potent inhibitors of Candida albicans acetohydroxyacid synthase.Journal of Medicinal Chemistry,2013,56(1):210-219.
[17]KREISBERG J F,ONG N T,KRISHNA A,et al.Growth inhibition of pathogenic bacteria by sulfonylurea herbicides.Antimicrobial Agents and Chemotherapy,2013,57(3):1513-1517.
[18]RICHIE D L,THOMPSON K V,STUDER C,et al.Identification and evaluation of novel acetolactate synthase inhibitors as antifungal agents.Antimicrobial Agents and Chemotherapy,2013,57(5):2272-2280.
[19]PUE N,GUDDAT L W.Acetohydroxyacid synthase:A target for antimicrobial drug discovery.Current Pharmaceutical Design,2014,20(5):740-753.
[20]GOKHALE K, TILAK B. Mechanisms of bacterial acetohydroxyacid synthase(AHAS)and specific inhibitors of Mycobacterium tuberculosis AHAS as potential drug candidates against tuberculosis.Current Drug Targets,2015,16(7):689-699.
[21]JUNG I P,HA N R,LEE S C,et al.Development of potent chemical antituberculosis agents targeting Mycobacterium tuberculosis acetohydroxyacid synthase.International Journal of Antimicrobial Agents,2016,48(3):247-258.
[22]叶朝燕,张阳,汪冒君,等.镰刀菌乙酰乳酸合成酶抑制剂的筛选及抗菌性分析.浙江大学学报(理学版),2015,42(2):166-171,220.YE C Y,ZHANG Y,WANG M J,et al.Screening and fungistatic activity ofacetolactate synthase inhibitors for Fusarium graminearum.Journal of Zhejiang University(Science Edition),2015,42(2):166-171,220.(in Chinese with English abstract)
[23]XU T,YAO F,LIANG W S,et al.Involvement of alternative oxidase in the regulation of growth,development,and resistance to oxidative stress ofSclerotinia sclerotiorum.Journalof Microbiology,2012,50(4):594-602.
[24]XU T,WANG Y T,LIANG W S,et al.Involvement of alternative oxidase in the regulation of Sclerotinia sclerotiorum sensitivity to fungicides of azoxystrobin and procymidone.Journalof Microbiology,2013,51(3):352-358.
[25]梁五生,徐婷,姚飞,等.交替氧化酶可增强核盘菌对多菌灵的敏感性.中国农学通报,2016,32(6):193-197.LIANG W S,XU T,YAO F,et al.Enhancing effect of alternative oxidase on sensitivity of Sclerotinia sclerotiorum to carbendazim.Chinese Agricultural Science Bulletin,2016,32(6):193-197.(in Chinese with English abstract)
[26]BRADFORD M M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding.Analytical Biochemistry,1976,72(1/2):248-254.
[27]KALME S,PHAM C N,GEDI V,et al.Inhibitors of Bacillus anthracis acetohydroxyacid synthase.Enzyme and Microbial Technology,2008,43(3):270-275.
[28]LEE H,RUSTGI S,KUMAR N,et al.Single nucleotide mutation in the barley acetohydroxy acid synthase(AHAS)gene confers resistance to imidazolinone herbicides.Proceedings of the National Academy of Sciences of the USA,2011,108(21):8909-8913.
[29]GEDI V,YOON M Y.Bacterial acetohydroxyacid synthase and its inhibitors:A summary of their structure,biological activity and current status.The FEBS Journal,2012,279(6):946-963.
[30]EVANS D L,JOSHI S V.Elucidating modes of activation and herbicideresistancebysequenceassemblyand molecular modelling of the acetolactate synthase complex in sugarcane.Journal of Theoretical Biology,2016,407:184-197.
[31]BROSNAN J T,VARGAS J J,BREEDEN G K,et al.A new amino acid substitution(Ala-205-Phe)in acetolactate synthase(ALS)confers broad spectrum resistance to ALS-inhibiting herbicides.Planta,2016,243(1):149-159.
[32]HUANG Z F,CHEN J Y,ZHANG C X,et al.Target-site basis for resistance to imazethapyr in redroot amaranth(Amaranthus retroflexus L.).Pesticide Biochemistry and Physiology,2016,128:10-15.
[33]SINGH B K,STIDHAM M A,SHANER D L.Assay of acetohydroxyacid synthase.Analytical Biochemistry,1988,171(1):173-179.
[34]DU Y,ZHANG H F,HONG L,et al.Acetolactate synthases MoIlv2 and MoIlv6 are required for infection-related morphogenesis in Magnaporthe oryzae.Molecular Plant Pathology,2013,14(9):870-884.
Screening of acetolactate synthase inhibitors against Sclerotinia sclerotiorum for the purpose of controlling rapeseed Sclerotinia disease.Journal of Zhejiang University(Agric.&Life Sci.),2017,43(5):589-598
WEN Xuewei,CHEN Yangyang,WU Bin,HU Dongwei,LIANG Wusheng*
(Ministry of Agriculture Key Laboratory of Molecular Biology of Crop Pathogens and Insects/Institute of Biotechnology,College of Agriculture and Biotechnology,Zhejiang University,Hangzhou 310058,China)
acetolactate synthase;Sclerotinia disease;Sclerotinia sclerotiorum;rapeseed;sulfathiazole
S 432.44
A
10.3785/j.issn.1008-9209.2017.02.201
Summary Rapeseed is an oil crop with quite large growing areas in China,and also one of the main oil crops in the world.As one of the most serious diseases in rapeseed,Sclerotinia disease,caused by the fungal pathogen Sclerotinia sclerotiorum,can result in significant yield loss.At present,no rape cultivars are available that are resistant to S.sclerotiorum.Using fungicides is the main measure to control rapeseed Sclerotinia disease in China.Carbendazim,as a common fungicide,has been frequently applied to control rapeseed Sclerotinia disease for long.Increasing evidence of fungicide resistance in populations of S.sclerotiorum has been found.Therefore,new fungicides are constantly being requested to control rapeseed Sclerotinia disease.Acetolactate synthase(ALS)is the enzyme that catalyzes the first step in the branched-chain aminoacid biosynthesis pathway in plants and microbes,and is the target of some herbicides.Recently,some studies have reported the development of new antibiotics to control bacterial pathogens or new fungicides to control fungal pathogens of human beings with ALS as the action target.However,there are few studies on developing new fungicides to control fungal pathogens of plants with ALS as the action target.
国家自然科学基金(31370279);浙江省自然科学基金(LY13C140001);浙江省教育厅科研计划项目(Y201327327)。
梁五生(http://orcid.org/0000-0001-9360-1187),Tel:+86-571-86433024,E-mail:liangws@zju.edu.cn
(First author):温雪玮(http://orcid.org/0000-0002-0886-3930),E-mail:1018559277@qq.com
2017-02-20;接受日期(Accepted):2017-04-28
In the present study,seven potential ALS inhibitors(chlorimuron-ethyl,bensulfuron-methyl,chlorsulfuron,sulfometuronmethyl,imazethapyr,imazaquin and sulfathiazole)were used to determine the effectiveness of controlling rapeseed Sclerotinia disease.Chlorimuron-ethyl,bensulfuron-methyl,chlorsulfuron and sulfometuron-methyl were chosen as the representatives of sulfonylurea-type ALS inhibitors.Imazethapyr and imazaquin were the representatives of imidazolinone-type ALS inhibitors,and sulfathiazole was the representative of sulfathiazole-type ALS inhibitors.
The results indicated that the activity of ALS in S.sclerotiorum could not been inhibited by 1.0 mg/L imazethapyr.In contrast,the treatment with 1.0 mg/L chlorimuron-ethyl,bensulfuron-methyl,chlorsulfuron,sulfometuron-methyl,imazaquin and sulfathiazole showed more or less inhibitory effects on the ALS activity of S.sclerotiorum.Among them,imazaquin had the strongest inhibitory effect on ALS,but sulfometuron-methyl showed the weakest inhibitory effect.Imazaquin and sulfathiazole could effectively suppress the growth of S.sclerotiorum colony on potato dextrose agar plates at the concentration of 0.1 mg/L.Chlorimuron-ethyl,bensulfuron-methyl and chlorsulfuron had inhibitory effect at the concentration of 1.0 mg/L.However,sulfometuron-methyl and imazethapyr showed weak inhibitory effect on the growth of S.sclerotiorum colony even at the concentration of 50 mg/L.The half-maximal effective concentrations(EC50)of imazaquin,sulfathiazole,chlorimuron-ethyl,bensulfuron-methyl and chlorsulfuron were determined to be less than 10 mg/L,but EC50values of sulfometuron-methyl and imazethapyr were determined to be more than 50 mg/L.All the seven potential ALS inhibitors showed inhibitory effects on the activity of ALS in rapeseed leaves at the concentration of 1.0 mg/L.The inhibitory effects of chlorimuron-ethyl,bensulfuronmethyl,chlorsulfuron,imazaquin and sulfometuron-methyl were much stronger than those of imazethapyr and sulfathiazole.The inhibitory effect of sulfathiazole was the weakest among the seven inhibitors.Significant damaging influences were observed on rapeseed seedlings by using chlorimuron-ethyl,bensulfuron-methyl,chlorsulfuron,imazaquin,sulfometuron-methyl and imazethapyr at the concentration of 1.0 mg/L,such as decreasing growth rate,dwarfing,reducing leaf size and leaf yellowing,but no significant influences were observed by treating rapeseed seedlings with sulfathiazole.The use of sulfathiazole could exert remarkable inhibitory effects against ALS activity and colony growth of S.sclerotiorum without giving rise to harmful influence on rapeseed seedlings.Thus,the effect of sulfathiazole on S.sclerotiorum infection to rapeseed was tested.The result indicated that the treatment with sulfathiazole significantly reduced the infection scale of S.sclerotiorum in rapeseed leaves.
In conclusion,the present research indicated that S.sclerotiorum was sensitive to imazaquin,sulfathiazole,chlorimuron-ethyl,bensulfuron-methyl and chlorsulfuron other than sulfometuron-methyl and imazethapyr.On the other hand,rapeseed was sensitive to all the seven ALS inhibitors but imazethapyr and sulfathiazole.It is suggested that sulfathiazole treatment is an effective strategy to control rapeseed Sclerotinia disease caused by S.sclerotiorum.Therefore,this study concludes that sulfathiazole is a potential molecular structure basis for controlling rapeseed Sclerotinia disease with ALS of S.sclerotiorum as an action target.
猜你喜欢
四川农业科技(2022年2期)2022-11-21
今日农业(2021年21期)2021-11-26
今日农业(2021年14期)2021-10-14
今日农业(2021年7期)2021-07-28
四川蚕业(2021年3期)2021-02-12
中华养生保健(2020年3期)2020-11-16
上海蔬菜(2020年5期)2020-10-20
农药科学与管理(2019年9期)2019-11-23
科技视界(2016年26期)2016-12-17
英语学习(2015年12期)2015-02-01
