
四倍体彩色马蹄莲的生长及对低温胁迫的生理响应
2017-11-14 09:46贺水莲吴景芝陆燕许春梅吴红芝
浙江大学学报(农业与生命科学版) 2017年5期
贺水莲,吴景芝,陆燕,许春梅,吴红芝,3*
四倍体彩色马蹄莲的生长及对低温胁迫的生理响应
贺水莲1,吴景芝1,陆燕2,许春梅1,吴红芝1,3*1
(1.云南农业大学园林园艺学院,昆明650201;2.昆明市西山区农林局,昆明650100;3.连云港西诺花卉种业有限公司,江苏东海222331)
以彩色马蹄莲栽培品种Tanswan和Liberty的四倍体及二倍体组培苗为材料,在同一环境条件下进行炼苗移栽,并采取完全相同的栽培管理措施,比较分析不同倍性彩色马蹄莲植株的成苗情况、形态学特征、种球干物质积累、休眠性及抗寒性特征。结果表明:与二倍体相比,四倍体彩色马蹄莲倍性稳定,植株成苗率相对较高;叶片增大,叶色加深,表现出明显的器官巨大性;种球营养物质积累量增多;生育期提前,休眠期变短。低温胁迫后,Tanswan和Liberty 2个彩色马蹄莲品种四倍体植株的抗寒性和抗低温伤害能力比二倍体更强,植株成活率分别增加23.59%和26.74%,显著高于其对应的二倍体植株。通过对低温胁迫后彩色马蹄莲中游离脯氨酸含量及超氧化物歧化酶、过氧化物酶活性的比较发现,马蹄莲抗寒性与脯氨酸含量变化成反比,揭示了四倍体彩色马蹄莲抗寒性增强的生理机制。综上表明,彩色马蹄莲四倍体具有很好的生产推广及应用潜力。
彩色马蹄莲;多倍体;生长特征;形态学特征;低温抗性
彩色马蹄莲(Zantedeschia hybrida Spr.)又称彩色海芋,原产南非,为天南星科(Araceae)马蹄莲属(Zantedeschia)多年生草本球根花卉[1-2]。形态高雅、色彩艳丽、花叶俱赏的彩色马蹄莲用途广泛,除可用作优良的切花、盆花外,还是花束、花篮,以及花坛、花镜等的重要材料[3]。自20世纪90年代进入国内市场后,彩色马蹄莲深受消费者青睐,被誉为21世纪“花卉之星”[4]。但由于目前彩色马蹄莲品种多为国外即将淘汰的老品种,经多代无性繁殖后其种球生活力、抗病性等内在品质下降,且种球在重复使用过程中受病虫害、土壤、气候等逆境影响退化十分严重,极大地影响了彩色马蹄莲的生产和质量[5-7]。此外,彩色马蹄莲种植技术难度大、投入高、成品率低等普遍观念[8]也极大地制约了彩色马蹄莲产业在中国的发展。因此,培育观赏性状优良、抗病、生长势强的新品种对我国彩色马蹄莲产业具有重要意义[9-11]。
与二倍体相比,植物多倍体一般具有花朵增大、着色加深、花瓣增厚、耐贮运、抗病性增强等特点,因此,多倍体品种往往具有更好的商品品质。而染色体鉴定结果表明,目前市场上的彩色马蹄莲品种均是二倍体。笔者之一吴红芝等[10]通过多倍体育种技术获得了彩色马蹄莲四倍体,但对于其是否比二倍体生长更强健、具有更好的抗性及其内在生理机制等问题尚未进行深入研究。因此,本研究以彩色马蹄莲多倍体变异组培苗为材料,以相同品种彩色马蹄莲二倍体组培苗为对照,对彩色马蹄莲四倍体植株的形态学、生长发育、低温响应等进行研究,以期为彩色马蹄莲多倍体新品种的培育提供理论依据。
1 材料与方法
1.1 试验时间与地点
试验时间为2013年3月—2014年12月;试验在云南农业大学园林园艺学院的温室内进行。
1.2 试验材料
以彩色马蹄莲Tanswan和Liberty的四倍体及二倍体组培苗为试验材料。其中,四倍体均由二倍体诱导变异产生。
1.3 炼苗移栽
将在同等培养条件下、生长发育相似的彩色马蹄莲Tanswan和Liberty的二倍体和四倍体组培苗同时移出,揭去覆膜,在室温下进行3~4 d的炼苗。炼苗结束后,仔细清洗以去掉苗上的培养基,之后将其移栽到基质中,并标明品种及倍性。采用100 cm×60 cm×40 cm规格的塑料筐为种植框,种植基质分别为:V(草炭)∶V(珍珠岩)=6∶4(A1组);V(草炭)∶V(珍珠岩)=7∶3(A2组)。设3个重复,每个处理每个倍性的组培苗各取50株。
1.4 水肥及光照管理
遮阴和水肥管理对彩色马蹄莲组培苗的移栽成活率及生长势有极大影响[12]。本研究在幼苗期施用表1所示的高氮复合肥以促进叶片生长,并覆遮阳密度为50%的银色遮阳网一层;待叶片生长基本停止后施用表2所示的高磷高钾复合肥以促进种球干物质积累,同时揭去遮阳网;当地上部分开始有枯黄迹象时逐渐减少浇水次数和浇水量以促进后期种球膨大。
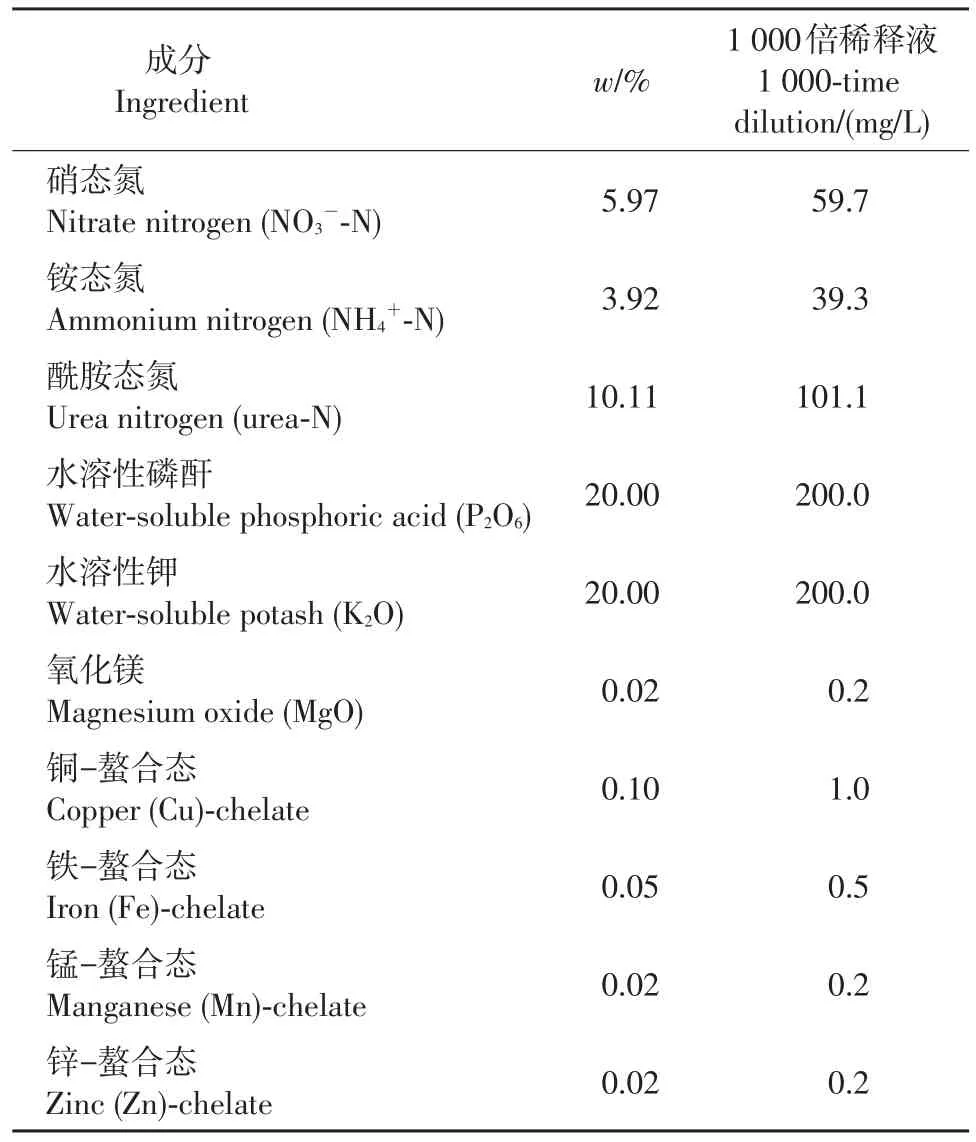
表1 N-P-K(20-20-20+1 MgO+TE)平衡的苗期肥Table 1 Seedling fertilizer of N,P,K balance(20-20-20+1 MgO+TE)

表2 富含P、K(15-50-35+2 MgO+TE)的开花肥Table 2 Flowering fertilizer with rich P,K(15-50-35+2 MgO+TE)
1.5 形态指标及测定方法
移栽后,待小苗生长至60 d时观察记录株高、茎粗、最大叶长、最大叶宽、叶形指数、叶色、存活率及多倍体植株倍性稳定性;种球采收后测量其鲜质量、干质量、干鲜比。各项指标测定方法如下:植株倍性采用根尖染色体鉴定法;在植株形态特征中,株高为茎基部到最长叶尖的长度,茎粗为用绳子缠绕茎后绳的长度,最大叶长为最大叶片的叶基部到叶尖的距离,最大叶宽为最大叶片的最宽部位的长度,叶形指数为最大叶长与最大叶宽之比,叶色用目测法观察。干鲜比=种球干质量/种球鲜质量。其中,种球鲜质量为采收时的质量,种球干质量为在室温下风干15 d后的质量。成活率=成活株数/播种数×100%。
1.6 抗寒性鉴定
以不同倍性的Tanswan和Liberty组培苗移栽后2个月、生长健壮的幼苗为材料,每个品种挑选40株,移入人工光照培养箱,在平均温度为0℃、光照时间为14 h的条件下进行低温胁迫处理,1周后统计成活率。同时,分别采集对照与低温处理的植株叶片测定其生理生化指标。过氧化物酶(peroxidase,POD)用愈创木酚法测定,超氧化物歧化酶(superoxide dismutase,SOD)用硝基四氮唑蓝(nitroblue tetrazolium,NBT)显色法测定[13];脯氨酸用茚三酮显色法测定[14]。
1.7 休眠特性检测
将秋季地上部分茎叶枯死进入休眠后的彩色马蹄莲种球(未采收)连同栽培框、基质一起,共同放置于昆明冬季自然低温下,于第2年3月中旬气温回暖时将种球(T2代)连同种植框移到温室内,恢复浇水以促进种球萌发,观察统计其萌发生长情况。
1.8 种球采收
在叶片完全变黄、枯萎时进行种球采收。将种球从种植框内小心挖出,避免机械损伤;采挖后先在有防雨设施、通风良好、无阳光直射的地方将种球阴干,让土块自行脱落,再进行清洗、分级和杀菌处理。种球阴干期间要及时加强通风,使其尽快干燥。
2 结果与分析
2.1 四倍体植株的成苗率分析
彩色马蹄莲四倍体组培苗经5代继代培养后,其性状和倍性保持稳定。但与组培环境条件较稳定不同,变异苗在栽培过程中可能会因环境条件的变化发生有丝分裂异常。因此,在移栽2个月后,随机挑选10个植株进行根尖染色体制片观察。结果表明,彩色马蹄莲四倍体的染色体数目为64,是二倍体的2倍(图1~2)。说明彩色马蹄莲四倍体植株在自然栽培条件下倍性稳定。

图1 彩色马蹄莲二倍体的染色体(2n=32)(16×100倍)Fig.1 Chromosome of the colored calla lily diploid(2n=32)(16×100 times)

图2 彩色马蹄莲四倍体的染色体(4n=64)(16×100倍)Fig.2 Chromosome of the colored calla lily tetraploid(4n=64)(16×100 times)
为了研究不同倍性彩色马蹄莲的成活率,在不同基质中开展了移栽试验。将带有3~4个健康叶片、不同倍性的彩色马蹄莲组培苗移栽到透气性大小不同的基质中(A1:基质草炭含量较低,珍珠岩含量较高,透气性高;A2:基质草炭含量较高,珍珠岩含量较低,透气性低),分别观察统计彩色马蹄莲组培苗的成活率及生长情况。结果(表3)表明:在A1基质栽培条件下的彩色马蹄莲组培苗成活率显著高于A2基质栽培条件,说明基质的透气性对彩色马蹄莲根系生长十分重要;在相同基质栽培条件下四倍体植株的成活率明显高于二倍体植株;比较同一基质中相同倍性苗的成活率发现,彩色马蹄莲组培苗的移栽成活率与品种有关。
2.2 多倍体植株形态学特征
当植株株高、茎粗、叶长、叶宽等达到最大值后进行形态特征观测。从表4可以看出,四倍体植株更加健壮,叶形向钝圆发展,其表型数据明显优于二倍体植株。研究认为,叶形指数越靠近1,则叶形越接近椭圆形,其观赏性越好,光合作用面积越大[15]。本研究2个品种的四倍体叶形指数都比二倍体更接近1:说明四倍体植株具有更高的观赏性,能更好地进行光合作用。
2.3 种球干物质的积累
种球是球根花卉贮藏营养的主要器官,其干物质积累量与种球的品质、抗性及适应性密切相关[12]。从表5中可以看出:不同品种间种球的鲜质量、干质量存在较大差异;四倍体的干质量、鲜质量均显著大于二倍体,且Tanswan和Liberty 2个品种四倍体植株的干鲜比分别比二倍体提高了15.3%和13.2%。说明四倍体植株对干物质的积累效率优于二倍体。
2.4 四倍体植株的抗寒性
2013年冬天昆明遭遇雨雪低温天气约1周,种植于简易大棚中(温度约为2~10℃)的T1代彩色马蹄莲受到严重影响,采收后统计发现二倍体的死亡率明显高于四倍体。为了进一步确证彩色马蹄莲四倍体的抗寒性优于二倍体,以移栽成活的不同倍性的马蹄莲组培苗为材料,经0℃低温处理3 d,分析其成活率、游离脯氨酸含量、超氧化物歧化酶(SOD)及过氧化物酶(POD)活性等抗寒相关指标。
2.4.1 成活率
如表6所示,Tanswan和Liberty的四倍体植株成活率分别增加了23.59%和26.74%,显著高于其相应的二倍体植株:表明彩色马蹄莲四倍体植株在抗寒能力方面明显优于二倍体。

表3 彩色马蹄莲四倍体和二倍体组培苗成活率比较Table 3 Comparison on survival rate of tetraploid and diploid of colored calla lily in vitro

表4 彩色马蹄莲四倍体和二倍体植株形态学特征比较Table 4 Comparison on morphological characteristics of tetraploid and diploid of colored calla lily in vitro

表5 彩色马蹄莲四倍体与二倍体种球干物质积累比较Table 5 Comparison on dry matter accumulation in bulbs of tetraploid and diploid of colored calla lily in vitro

表6 低温对彩色马蹄莲四倍体与二倍体幼苗的影响Table 6 Effects of low temperature on tetraploid and diploid seedlings of colored calla lily
2.4.2 游离脯氨酸含量
脯氨酸作为渗透物质,在植物体内起着渗透调节作用[16]。植物在遭遇干旱、低温、盐碱等逆境时,游离脯氨酸含量便会大量积累,并且积累量与植物抗逆性有关[13]。由表7可知,经过低温胁迫处理的植株游离脯氨酸含量都有不同程度的升高。运用SPSS 17.0软件进行显著性分析发现,经低温胁迫处理后的2个品种的游离脯氨酸含量都极显著高于常温。此外,在常温下2个品种二倍体的游离脯氨酸含量极显著高于四倍体,且经低温胁迫处理后,二倍体植株的游离脯氨酸含量增加程度也远大于四倍体。

表7 不同倍性彩色马蹄莲游离脯氨酸含量变化Table 7 Change of free proline content in the diploid and tetraploid of colored calla lily
2.4.3 超氧化物歧化酶(SOD)活性
超氧化物歧化酶(SOD)是需氧生物体内普遍存在的一种含金属的保护性酶。SOD活性和超氧阴离子自由基产生速率都可以表示植物在抗寒过程中的生理状况[17],所以SOD活性的变化可以作为植物抗逆性分析的参考指标。由表8可知,经过低温胁迫处理的植株,其SOD活性都不同程度地高于常温对照。使用SPSS 17.0软件进行显著性分析发现:Liberty四倍体的SOD活性在低温胁迫后极显著高于二倍体,在常温处理下也显著高于二倍体;Tanswan在低温处理及常温处理下四倍体与二倍体之间在统计学上都没有显著差异。根据SOD活性分析可得抗寒性强弱顺序为 Liberty(4n)>Liberty(2n),Tanswan(4n)>Tanswan(2n)。

表8 不同倍性彩色马蹄莲SOD活性变化Table 8 Change of SOD activity in the diploid and tetraploid of colored calla lily
2.4.4 过氧化物酶(POD)活性
POD是膜保护系统的组成部分,是植物体内自由基的有效清除剂[18]。从表9可以看出:经低温胁迫处理后,所有植株的POD活性都有不同程度的上升;Tanswan和Liberty 2个品种的四倍体POD活性增加幅度均大于二倍体。显著性分析结果显示,经低温胁迫后,Liberty和Transwan 2个品种的四倍体的POD活性均极显著高于各自的二倍体。说明经低温胁迫后四倍体彩色马蹄莲的抗氧化能力显著增强。

表9 不同倍性彩色马蹄莲POD活性变化Table 9 Change of POD activity in the diploid and tetraploid of colored calla lily
2.5 四倍体休眠特性
将秋季地上部分茎叶枯死进入休眠后的彩色马蹄莲种球连同基质一起放置于昆明冬季自然低温下,发现第2年四倍体与二倍体种球的萌发时间不同:Liberty和Transwan 2个彩色马蹄莲四倍体种球的萌发时间均早于对应的二倍体(表10)。在同一栽培条件下种植55 d时,2个品种的四倍体已生长成为15~20 cm的健壮植株,而二倍体才刚刚出苗(图3~4)。说明四倍体种球的休眠时间缩短,生育期提前。

表10 彩色马蹄莲四倍体与二倍体T2种球出苗时间比较Table 10 Comparison of bulb germination time between tetraploid and diploid of colored calla lily

图3 Liberty品种Fig.3 Liberty variety

图4 Tanswan品种Fig.4 Tanswan variety
3 讨论
与二倍体植株相比,彩色马蹄莲四倍体植株生长健壮、叶柄较粗、叶色浓绿、叶面积变大、叶片形状钝圆变宽:说明彩色马蹄莲四倍体植株表现出明显的器官巨大性。
植物受到逆境胁迫后,其生理变化错综复杂。在逆境条件下,大部分植物均会积累大量的脯氨酸以保持细胞水分和生物大分子结构的稳定性,从而适应逆境,防止活性氧对膜脂和蛋白质的过氧化作用[19]。本研究结果表明,在受到低温胁迫后,马蹄莲二倍体与四倍体植株游离脯氨酸含量都有明显增加:说明彩色马蹄莲受到低温胁迫后,通过脯氨酸调节的防御反应被积极响应。陈雅君等[20]提出,在低温胁迫下脯氨酸的积累能力与品种的抗寒力呈负相关,认为抗寒性弱的品种在受到低温胁迫后,为适应寒冷,保护体内组织免受冻害,会积累大量的游离脯氨酸,而游离脯氨酸积累较少的品种,其自身相应的抗寒性较强。本研究二倍体马蹄莲的脯氨酸积累量均显著高于四倍体,说明彩色马蹄莲四倍体抗寒性强于二倍体。POD和SOD都是植物抗氧化相关酶,SOD能够歧化O2·-为O2和H2O2,而 POD则催化H2O2形成 H2O,因此,POD和SOD的活性与植物抗逆性呈正相关。通过比较低温胁迫后POD和SOD的活性发现,在低温胁迫下两者活性均有明显上升,且2个品种四倍体的POD和SOD活性上升程度明显高于二倍体:说明彩色马蹄莲四倍体植株比其二倍体植株抗低温伤害能力更强。
通过对休眠特性的研究发现,彩色马蹄莲四倍体种球休眠期明显缩短,这可能是相关基因表达量增加的缘故。彩色马蹄莲四倍体休眠期缩短,不仅在实际应用中能够节省低温贮藏种球的成本,在理论研究中还可以利用彩色马蹄莲四倍体差异表达基因来进一步挖掘控制马蹄莲休眠的相关基因。
早在20世纪90年代,就有利用体细胞加倍获得四倍体彩色马蹄莲的报道[21]。但目前市场上的彩色马蹄莲商业品种均为二倍体,其原因可能是顶端优势明显的彩色马蹄莲在诱变成多倍体后,营养生长旺盛的特点更加突出,对基质、光照、水肥等的需求发生了较大的变化,而种植者依旧采用二倍体的栽培方法,容易使彩色马蹄莲多倍体营养生长过盛、徒长、易感病,因此,四倍体马蹄莲未能进入生产应用。
本研究通过对彩色马蹄莲四倍体与二倍体进行植株形态、成苗情况、种球干物质积累、休眠性及在低温胁迫下的生理特征的分析比较,明确了彩色马蹄莲四倍体具有植株器官巨大性显著、休眠期缩短、抗寒性增强、观赏价值高于二倍体植株等特点,认为四倍体彩色马蹄莲具有很大的推广应用潜力。因此,在未来的工作中可以进一步研究光、温、水、气、肥等对彩色马蹄莲多倍体栽培的影响,探索适宜彩色马蹄莲多倍体的科学栽培技术和病虫害防治技术,为彩色马蹄莲多倍体新品种的开发和推广应用奠定理论基础。
[1] LETTY C.The genus Zantedeschia.Bothalia,1973,11(1/2):5-26.
[2] SINGH Y,VAN WYK A E,BAIJNATH H.Taxonomic notes on the genus Zantedeschia Spreng.(Araceae)in southern Africa.South African Journal of Botany,1996,62(6):321-324.
[3] KUEHNY J.Calla history and culture.HortTechnology,2000,10(2):267-274.
[4] 赵培飞,吴丽芳,郑凌,等.云南彩色马蹄莲的生产与发展前景.湖南农业科学,2002(3):69-70.ZHAO P F,WU L F,ZHENG L,et al.The cultivation and development of colored common calla lily(Zantedeschia spp.)in Yunnan.Hunan Agricultural Sciences,2002(3):69-70.(in Chinese with English abstract)
[5] 闫永庆,王昆,樊金萍.彩色马蹄莲生产管理技术.中国林副特产,2005(3):12-13.YAN Y Q,WANG K,FAN J P.The production and management techniques of Zantedeschia aethiopica.Forest By-Product and Speciality in China,2005(3):12-13.(in Chinese with English abstract)
[6] 赵小菊.彩色马蹄莲杂交品种的栽培与切花生产技术.农业科技与信息,2009(17):34-35.ZHAO X J.Cultivation of Zantedeschia aethiopica hybrids and the production technology of cut flower.Agricultural Science and Technology and Information,2009(17):34-35.(in Chinese)
[7] 马宗新.彩色马蹄莲种球繁育和栽培关键技术研究.农业工程技术:温室园艺,2009(7):54-57.MA Z X.Key techniques of Zantedeschia aethiopica bulb breeding and cultivation.Agricultural Engineering Technology:Greenhouse Horticulture,2009(7):54-57.(in Chinese)
[8] 张勇,王莅,谭国栋,等.不同栽培处理对彩色马蹄莲生长影响的初步研究.天津农业科学,2007,13(4):32-34.ZHANG Y,WANG L,TAN G D,et al.Preliminary study on effect of different cultivation treatments on the growth of colorful Zantedeschia.Tianjin Agricultural Sciences,2007,13(4):32-34.(in Chinese)
[9] 邵果园.黄色马蹄莲(Zantedeschia elliotiana Engler)组培快繁研究及多倍体诱导初探.重庆:西南大学,2007:22-23.SHAO G Y.Primary studies on Zantedeschia ellintiana Engler by tissue rapid propagation and polyploidy induction.Chongqing:Southwest University,2007:22-23.(in Chinese with English abstract)
[10]吴红芝,张锡庆,郑思乡,等.彩色马蹄莲多倍体的诱导.园艺学报,2008,35(3):443-446.WU H Z,ZHANG X Q,ZHENG S X,et al.Polyploid induction of coloured Zantedeschia aethiopica.Acta Horticulturae Sinica,2008,35(3):443-446.(in Chinese with English abstract)
[11] 刘晓静,华小平,陆波,等.彩色马蹄莲品种‘Parfait’多倍体诱导及其生物学特征变化.植物资源与环境学报,2014,23(3):81-85.LIU X J,HUA X P,LU B,et al.Induction of Zantedeschia hybrid‘Parfait’polyploid and change of its biological characteristics.Journal of Plant Resources and Environment,2014,23(3):81-85.(in Chinese with English abstract)
[12]彭峰,陈嫣嫣,郝日明,等.彩色马蹄莲组培苗壮苗生根及移栽措施研究.江苏农业科学,2008(1):126-128.PENG F,CHEN Y Y,HAO R M,et al.Study on the growth and rooting of tissue culture plantlets of Zantedeschia hybrida and their transplanting method.Jiangsu Agricultural Sciences,2008(1):126-128.(in Chinese)
[13] 史树德,孙亚卿,魏磊,等.植物生理学实验指导.北京:中国林业出版社,2011:127-130.SHI S D,SUN Y Q,WEI L,et al.Plant Physiology Experiment Instruction.Beijing:China Forestry Publishing House,2011:127-130.(in Chinese)
[14]苍晶,赵会杰.植物生理学实验教程.北京:高等教育出版社,2013:143-145.CANG J,ZHAO H J.Plant Physiology Experiment Tutorial.Beijing:Higher Education Press,2013:143-145.(in Chinese)
[15]陈冬梅,和光宇,杨继先.彩色马蹄莲栽培技术要点.云南农业,2008(4):13.CHEN D M,HE G Y,YANG J X.The key cultivation techniques for the color calla lily.Yunnan Agriculture,2008(4):13.(in Chinese)
[16]李志亮,王刚,吴忠义,等.脯氨酸与植物抗渗透胁迫基因工程改良研究进展.河北师范大学学报(自然科学版),2005,29(4):404-408.LI Z L,WANG G,WU Z Y,et al.Proline and advance in relative genic engineering improvement for osmotic stress tolerance of plants.Journal of Hebei Normal University(Natural Science Edition),2005,29(4):404-408.(in Chinese with English abstract)
[17]高俊凤.植物生理实验指导.北京:高等教育出版社,2006:211-219.GAO J F.Plant Physiology Experiment Instruction.Beijing:Higher Education Press,2006:211-219.(in Chinese)
[18]陈少裕.膜脂过氧化对植物细胞的伤害.植物生理通讯,1991,27(2):84-90.CHEN S Y.Membrane lipid peroxidation damage to plant cells.Plant Physiology Communications,1991,27(2):84-90.(in Chinese)
[19]唐婉,胡杏,徐婉,等.几种紫薇属植物的抗寒性评价.西北农业学报,2012,21(9):121-126.TANG W,HU X,XU W,et al.Assessing the cold resistance of several crape myrtle(Lagerstroemia L.)species and cultivars.Acta Agriculturae Boreali-Occidentalis Sinica,2012,21(9):121-126.(in Chinese with English abstract)
[20]陈雅君,崔国文,富象乾.低温对苜蓿品种幼苗体内游离脯氨酸含量的影响.中国草地,1996(6):46-47.CHEN Y J,CUI G W,FU X Q.The effect of low temperature on free proline content in Alfalfa seedlings.Grassland of China,1996(6):46-47.(in Chinese with English abstract)
[21]COHEN D,YAO J L.In vitro chromosome doubling of nine Zantedeschia cultivars.Plant Cell,Tissue and Organ Culture,1996,47(1):43-49.
Growth of tetraploid Zantedeschia hybrida and its physiological response to low temperature.Journal of Zhejiang University(Agric.&Life Sci.),2017,43(5):570-578
HE Shuilian1,WU Jingzhi1,LU Yan2,XU Chunmei1,WU Hongzhi1,3*
(1.College of Landscape and Horticulture,Yunnan Agricultural University,Kunming 650201,China;2.Agricultural and Forestry Bureau of Xishan District of Kunming City,Kunming 650100,China;3.Lianyungang Sinoflor Flowers and Plant Co.,Ltd.,Donghai 222331,Jiangsu,China)
colored calla lily(Zantedeschia hybrida);polyploid;growth performance;morphological characteristics;resistance to low temperature
S 68
A
10.3785/j.issn.1008-9209.2016.06.162
Summary The colored calla lily(Zantedeschia hybrida),as the“flower star of the 21st century”,is becoming more and more popular for its special morphology and high ornamental value.But at present,the varieties of colored calla lily in the market are imported from foreign countries like the Netherlands,New Zealand,Japan,etc.Without independent intellectual property rights,the industry of colored calla lily has been seriously restricted in China;therefore,breeding new varietyies of colored calla lily with excellent ornamental traits,resistance to disease and strong growth vigor is of great significance to our country.Polyploids of ornamental plants are usually characterized by their superior growth vigor,flower size,stem sturdiness,and better resistance as compared with diploids.Two tetraploid variations of colored calla lily varieties“Tanswan”and“Liberty”were successfully established in the previous study;however,whether the tetraploid varaitions have more advantages in growth performance,morphological characteristics and resistance to stress than the diploids are still unclear.
In this study,the tissue culture plantlets of the diploid and tetraploid of colored calla lily were transplanted andmanaged at the same environmental condition and cultivation measure.Their ploidy level,survival rate,morphological characteristics(including plant height,stem diameter,maximum leaf length,maximum leaf width,leaf-shape index,leaf color,and uniformity),dry matter in bulb,resistance to stress and dormancy were investigated and analyzed.
国家自然科学基金(31260490);云南省农科教相结合新型农业社会化服务体系试点项目(2014NG002-08);云南省教育厅科学研究基金重大专项项目(ZD2014008);国家大学生创新创业训练计划建设项目(201410676004)。
吴红芝(http://orcid.org/0000-0003-2926-6075),E-mail:hwu1128@163.com
(First author):贺水莲(http://orcid.org/0000-0003-4969-6558),E-mail:heshuilian2006@163.com
2016-06-16;接受日期(Accepted):2017-03-21
The results showed that the chromosome number of two tetraploid variations was 64 after two months of transplanted into the field,suggesting the ploidy of the tetraploid variations are stable.The survival rate of colored calla lily cultured in the medium with more peat moss[V(peat moss)∶V(pelite)=6∶4]was higher than that in the medium of V(peat moss)∶V(pelite)=7∶3,which suggests the permeability of the medium is important to the growth of the colored calla lily.Compared with the diploid plants,the tetraploid plants of colored calla lily grew faster and stronger,had higher survival rate,thicker/darker green leaves,earlier germination time and shorter dormant period.Furthermore,the tetraploid plants had higher survival rate with an increase of 23.59%and 26.74%respectively in“Tanswan”and“Liberty”than the diploids under the low temperature stress,indicating the tetraploids of colored calla lily have higher resistance to low temperature.Under the same environmental condition,the difference in fresh mass and dry mass of the bulbs was quite significant at different ploidy levels.The tetraploids had higher ratio of dry to fresh mass than diploids,which suggests that the tetraploids have stronger ability to accumulate dry matter than the diploids.By comparing and analyzing the content of free proline,activities of superoxide dismutase(SOD)and peroxidase(POD),we found the tetraploids of colored calla lily had higher resistance to low temperature than the diploids;meanwhile,the resistance to low temperature stress of colored calla lily was negative correlation with the changes of free proline content,and the varieties with higher resistance to low temperature stress had a stronger antioxidation system.
In conclusion,the tetraploids of colored calla lily have significant advantages in morphological characteristics,dry matter accumulation in bulb,resistance to stress and dormancy characteristics;therefore,the tetraploid of colored calla lily has great potential to be applied the production in the future.
猜你喜欢
西北农业学报(2022年5期)2022-06-07
热带作物学报(2022年5期)2022-06-01
科学养鱼(2021年12期)2022-01-14
山西农业科学(2021年3期)2021-12-06
东北林业大学学报(2021年12期)2021-02-10
今日农业(2020年14期)2020-12-14
诊断学(理论与实践)(2020年6期)2020-12-09
动漫界·幼教365(小班)(2018年10期)2018-05-14
快乐作文·高年级(2017年9期)2018-01-15
散文诗世界(2016年6期)2016-11-22
