
不同碳源和生长因子条件下粒毛盘菌代谢产物初步研究
2017-11-08 09:15:37赵墨元杨柳青侯国华
微生物学杂志 2017年4期
石 芳, 赵墨元, 2, 杨柳青, 侯国华, 叶 明*
(1.合肥工业大学 食品科学与工程学院,安徽 合肥 230009;2.莫纳什大学 医药、护理与健康科学系,澳大利亚 3168)
不同碳源和生长因子条件下粒毛盘菌代谢产物初步研究
石 芳1, 赵墨元1, 2, 杨柳青1, 侯国华1, 叶 明1*
(1.合肥工业大学 食品科学与工程学院,安徽 合肥 230009;2.莫纳什大学 医药、护理与健康科学系,澳大利亚 3168)
采用GC-MS法对一种粒毛盘菌(Lachnumsp.)在不同碳源、生长因子条件下发酵代谢产物的挥发性成分组成与差异进行分析。结果显示,不同碳源和生长因子条件下产生的代谢产物不同,主要包括有机酸、胺类、烷烃类、酯类、醇类、吡咯等物质。分别以20 g/L的葡萄糖、蔗糖、淀粉为碳源的发酵液中检测到的挥发性代谢产物为7、7、10种;添加1 mg/L 的VC、VB1、甘氨酸、色氨酸作为不同生长因子的发酵液中检测到的挥发性代谢产物分别为6、7、7、12种。结果显示粒毛盘菌YM406发酵代谢产物具有丰富的多样性,并且在不同的培养条件下产生的代谢产物存在一定的差异。
粒毛盘菌;代谢产物;GC-MS;碳源;生长因子
近年来,从微生物次级代谢产物中发现了越来越多的活性物质。微生物次级代谢产物结构复杂多样,具有抗细菌、抗真菌、抗肿瘤、抗病毒等多种生物活性,是微生物药物开发的源泉[1]。此外,微生物代谢产物也可作为菌种鉴定的依据[2]。从微生物中获取新型骨架化合物,一般从两方面进行研究:一是增加微生物次级代谢产物的多样性,二是寻找快速有效的高通量筛选策略,最大限度地获取新的微生物次级代谢产物[3]。目前,人们已经从微生物次级代谢产物中发现了2万多个具有生物活性的化合物[4]。然而在进行微生物次级代谢产物研究时,如何提高微生物次级代谢产物的多样性是一个重要课题[5]。 微生物在常规培养条件下存在未表达的代谢途径,以至于许多代谢产物不能产生。因此,运用各种技术和方法激活这些沉默途径,获得结构多样的代谢产物已成为当今研究的热点[6]。研究表明同一微生物在不同培养条件下的代谢产物不同,如采用分段发酵的方法发现发酵后期温度不同,对乳酸菌繁殖和积累类细菌素有影响,不同的碳源和氮源的组合用量也能产生不同的结果[7];利用GC-MS方法检测发现不同培养和工艺条件下枯草芽胞杆菌的代谢产物不同[8];温度、时间、初始pH、接种量以及油脂添加量等不同发酵条件对发酵代谢产物中共轭亚油酸产量的影响也不同[9],不同温度和酸度条件对链霉菌属微生物的生长及代谢产物的影响不同[10]。而不同菌株在同一培养条件下的代谢产物也不同,如基因型不同的铜绿假单胞菌的菌株挥发性代谢产物是不同的[11]。 粒毛盘菌属(Lachnum)是一类腐生性真菌,个体较小,分布广泛,多生于腐朽的树木、叶茎或根部等处。长期以来,各国学者在粒毛盘菌多样性研究方面做了大量工作[12-17]。有学者在20世纪90年代发现粒毛盘菌在深层发酵条件下能产生脂酮类、异香豆素类等抗菌物质[18-19]。近几年来,本实验室发现一些粒毛盘菌菌株在深层发酵条件下产生大量多糖、色素及多酚等活性物质,其多糖具有抗氧化[20]、抗衰老[21]、降血糖[22]、降血脂[23]等活性,黑色素具有治疗贫血[24]、抗氧化[25]、抗紫外辐射[26]、抗衰老[27]等活性,但在粒毛盘菌代谢产物多样性研究方面则刚刚起步。本文研究了不同碳源和生长因子对粒毛盘菌YM406次级代谢产物的影响,为揭示粒毛盘菌代谢产物多样性、获得更多活性物质研究提供参考。
1 材料与方法
1.1材料
1.1.1 菌种来源 粒毛盘菌YM406子实体采集于安徽黄山,由合肥工业大学微生物资源与应用研究所分离保存。
1.1.2 培养基(g/L) ①活化培养基:葡萄糖20,酵母膏5,蛋白胨5,琼脂15,自然pH;②发酵培养基:葡萄糖20,酵母膏5,蛋白胨5,自然pH。
1.1.3 主要试剂及仪器 蛋白胨、酵母膏、琼脂等试剂购自天津市博迪化工有限公司;SW-CJ-ICU型超净工作台购自上海博迅实业有限公司;TDL-50B型台式离心机购自上海安亭科学仪器厂;250D型数显光照培养箱购自金坛市正基仪器有限公司;GC-MS-2010岛津购自深圳市华得隆科技有限公司。
1.2方法
1.2.1 菌种活化 将斜面保藏的粒毛盘菌菌株接种于活化培养基培养7 d,活化培养物作为摇瓶发酵的种子。
1.2.2 菌株发酵 菌株经平板活化后,用打孔器接种菌龄一致、直径为6 mm的菌块于装有150 mL发酵培养基的250 mL锥形瓶中培养10 d(发酵温度25 ℃,摇床转速160 r/min)。参考文献[28]的实验方法选择碳源与生长因子浓度,分别以20 g/L的蔗糖和淀粉替换葡萄糖研究不同碳源对粒毛盘菌代谢产物的影响,并以不接种的不同碳源培养基作为空白对照;在发酵培养基中分别添加1 mg/L的 VC、VB1、甘氨酸和色氨酸研究不同生长因子对粒毛盘菌代谢产物的影响,并以不接种的不同生长因子培养基作为空白对照。
1.2.3 不同培养条件下的代谢产物检测 分别取不同碳源、生长因子培养条件下的粒毛盘菌发酵液进行离心(6 000 r/min, 20 min),取适量上清液装入进样瓶中,进行GC-MS分析。 ①气相色谱条件: 色谱柱:HP-INNOWax(30 m×0.25 mm×0.25 mm);载气:氦气;柱流量:1.0 mL/min;进样口温度:250 ℃;程序升温:起始温度60 ℃,保持1 min,5 ℃/min 升至240 ℃,保持5 min,再以20 ℃/min升至260 ℃,保持2 min;进样量:1 μL;进样方式:不分流。 ②质谱条件: 电离方式:EI;电子能量:70 eV;离子源温度:230 ℃;接口温度:260 ℃;溶剂延迟:2 min;分析时间:2~45 min;模式:Scan;扫描范围:50~ 550 m/z。
2 结果与分析
2.1不同碳源条件下的代谢产物分析
菌株YM406在不同碳源条件下的空白培养基、发酵后的培养液GC-MS分析色谱图分别见图1、图2。色谱峰谱库比对结果见表1、表2。由表1可知,以葡萄糖为碳源的空白培养基检测出5种不同的化合物,其中醛类3种、有机酸2种。以蔗糖为碳源的空白培养基检测出7种不同的化合物,其中醛类3种、有机酸2种、醇类1种、胺类1种。以淀粉为碳源的空白培养基中仅检测出2种物质,分别为醛和有机酸。由表2可知,以葡萄糖为碳源的发酵液中检测出7种不同的化合物,其中有机酸2种,烷烃类、酯类、醚类、醇类、吲哚类各1种。以蔗糖为碳源的发酵液中检测出7种不同的化合物,其中有机酸3种,烷烃、胺类、醇类、酮类各1种。以淀粉为碳源的发酵液中检测出10种不同的化合物,其中含有胺类3种,烷烃、醇类各2种,有机酸、酯类、脲类各1种。
图1 不同碳源条件下的空白培养基GC-MS图谱 Fig.1 GC-MS spectra of blank medium with different carbon sourcesA: 葡萄糖; B: 蔗糖; C: 淀粉,图2同A: glucose; B: sucrose; C: starch,Figure 2 with

图2 不同碳源条件下的发酵液GC-MS图谱 Fig.2 GC-MS spectra of fermentation broth with different carbon sources
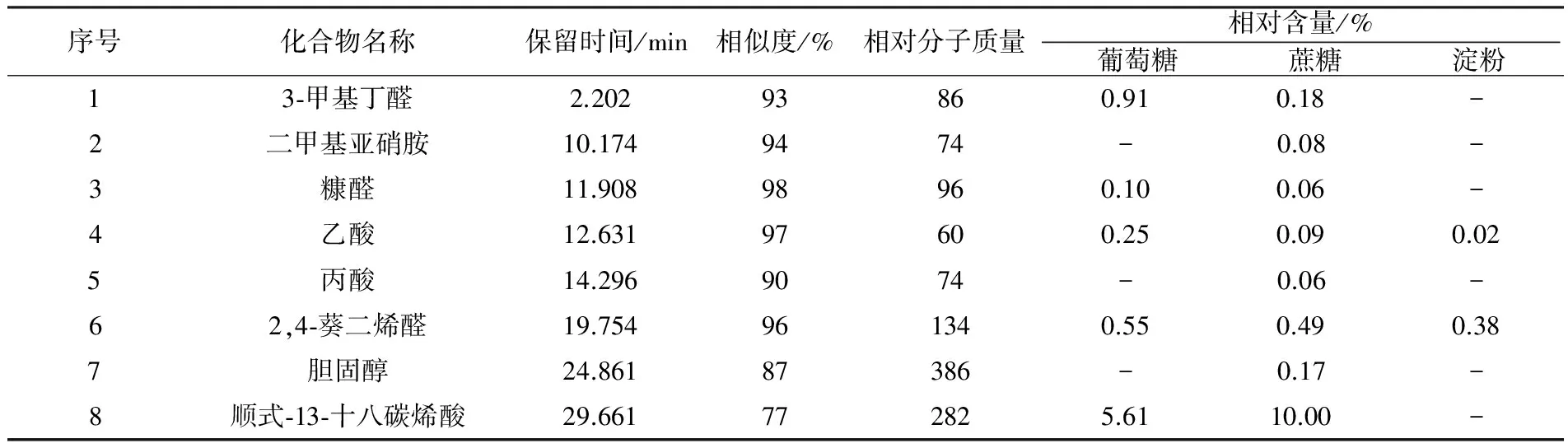
序号化合物名称保留时间/min 相似度/% 相对分子质量相对含量/% 葡萄糖 蔗糖 淀粉 13⁃甲基丁醛2.20293860.910.18-2二甲基亚硝胺10.1749474-0.08-3糠醛11.90898960.100.06-4乙酸12.63197600.250.090.025丙酸14.2969074-0.06-62,4⁃葵二烯醛19.754961340.550.490.387胆固醇24.86187386-0.17-8顺式⁃13⁃十八碳烯酸29.661772825.6110.00-
注:“-”表示未检出,下表同
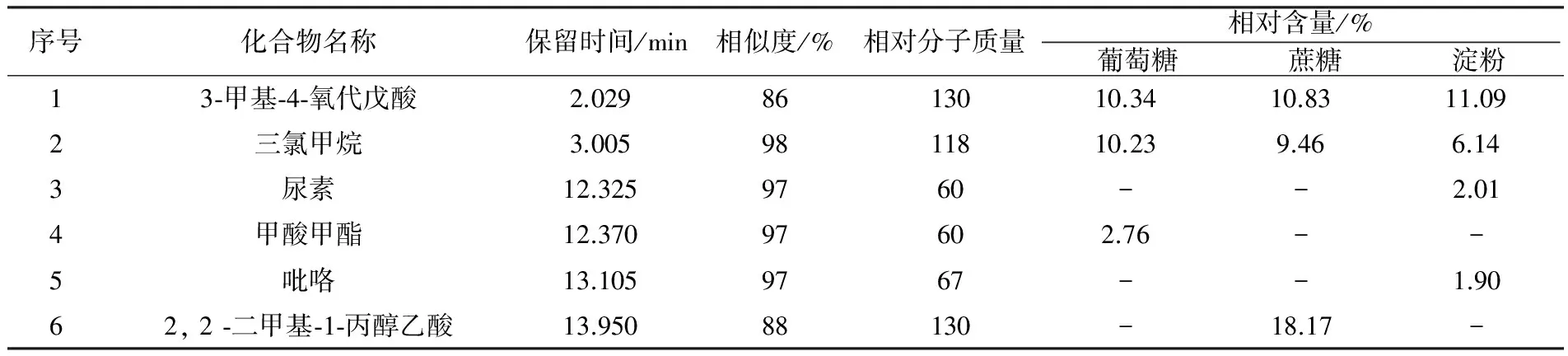
表2 不同碳源条件下的发酵液成分GC-MS分析
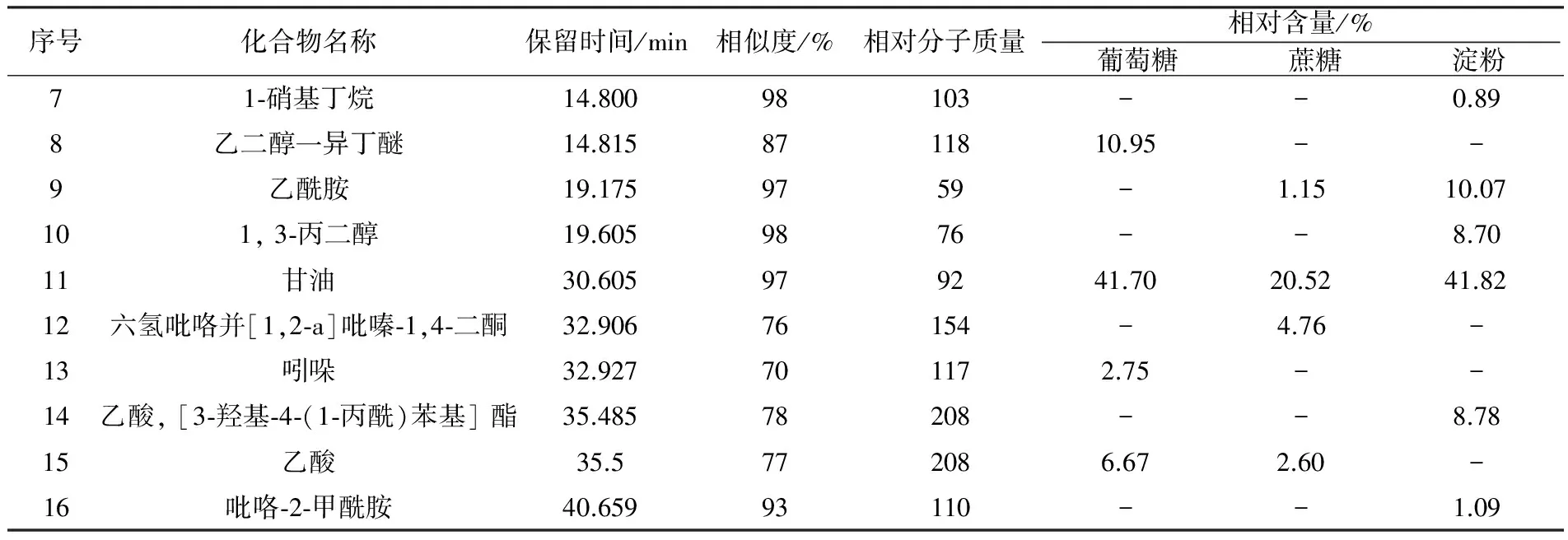
续表2
由此可见,以淀粉为碳源时,发酵液中检测出的挥发性代谢产物种类最多,为10种,其次是以葡萄糖和蔗糖为碳源的发酵液。这3组发酵液中,均检出了甘油、三氯甲烷、3-甲基-4-氧代戊酸3种物质,其中相对含量最高的是甘油。以葡萄糖为碳源时,发酵液中生成了其他两个处理组中未检测到的4种化合物,分别是甲酸甲酯(2.76%)、乙二醇一异丁醚(10.95%)、乙酸(6.67%)和吲哚(2.75%)。以蔗糖为碳源时,发酵液中生成了其他发酵液中未检测到的2种化合物分别为2, 2-二甲基-1-丙醇乙酸(18.17%)和六氢吡咯并[1,2-a] 吡嗪-1,4-二酮(4.76%)。以淀粉为碳源时,发酵液中生成了其他发酵液中未检测到的6种化合物,分别为尿素(2.01%)、吡咯(1.90%)、1-硝基丁烷(0.89%)、1, 3-丙二醇(8.70%)、乙酸、 [3-羟基-4-(1-丙酰)苯基] 酯(8.78%)和吡咯-2-甲酰胺(1.09%)。
2.2不同生长因子条件下的代谢产物分析
菌株YM406在不同生长因子条件下的空白培养基、发酵液的GC-MS分析色谱图分别见图3、图4。对应色谱峰的化合物和相对含量见表3、表4。
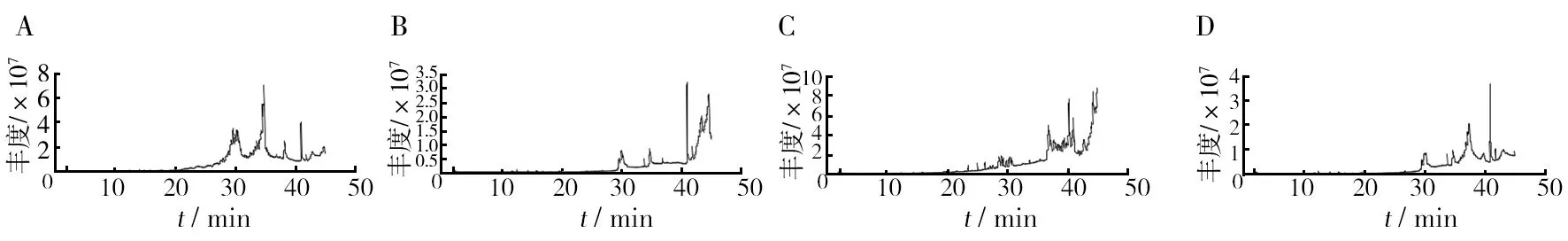
图3 添加不同生长因子的空白培养基GC-MS图谱 Fig.3 GC-MS spectra of blank medium with different growth factorsA: VC; B: VB1; C: 甘氨酸; D: 色氨酸,图4同A: VC; B: VB1; C: Glycine; D: TryptopHan,Figure 4 with
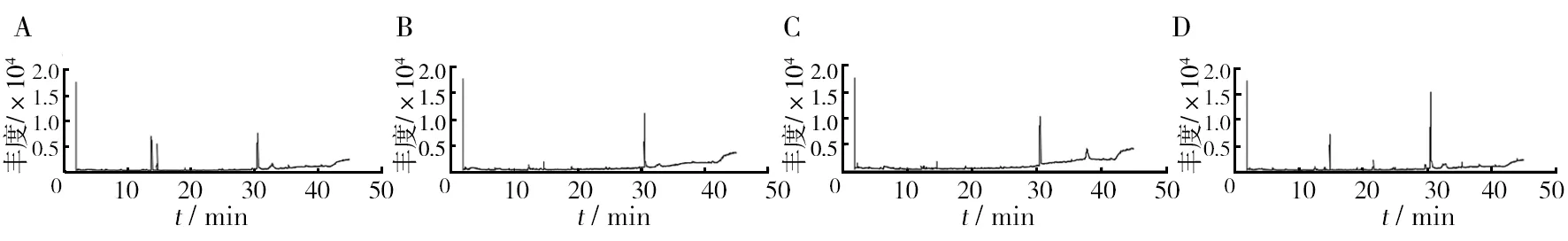
图4 添加不同生长因子的发酵液GC-MS图谱 Fig.4 GC-MS spectra of fermentation with different growth factors
由表3可知,添加VC为生长因子的空白培养基中检测出4种不同的化合物,其中有机酸2种、醛类1种、糖类衍生物1种。添加VB1为生长因子的空白培养基中检测出7种不同的化合物,其中醛类4种、有机酸2种、胺类1种。添加甘氨酸为生长因子的空白培养基中检测出4种物质,分别为有机酸2种、烷烃和酚类各一种。添加色氨酸为生长因子的空白培养基中检测出6种物质,分别为醛类3种、有机酸2种、烷烃1种。由表4可知,添加VC的发酵液中检测出6种不同的化合物,有机酸2种,胺类、酮类、醇类、吲哚类各1种。添加VB1的发酵液中检测出7种化合物,有机酸、醇类各2种,肟类、胺类、无机酸各1种。添加甘氨酸的发酵液中检测出7种不同的化合物,烷烃类、酯类、有机酸类、胺类、酚类、醇类、吡咯类各1种。添加色氨酸的发酵液中检测出12种物质,酯类2种,醇类2种,酮类2种,胺类、有机酸、醚类、胍类、烷烃类、吡啶类各1种。
由表4可见,添加色氨酸为生长因子时,发酵液中挥发性代谢产物种类最多,为12种,其次是添加甘氨酸和VB1的发酵液。这4组发酵液中,均含有2, 2-二甲基-1-丙醇乙酸和甘油2种物质,且VC组中2, 2-二甲基-1-丙醇乙酸相对含量最高,为26.35%。4组相对含量最高的均为甘油,VB1组的甘油相对含量高达67.12%。此外,各组中产生了一些其他组没有的化合物,如添加VC的发酵液中产生了2-吡咯烷酮(1.74%)、吲哚(4.55%);添加VB1的发酵液中产生了乙酸(6.53%)、甲氧基-苯基肟(2.00%)、磷酸(1.83%)、1-戊醇, 2,2-二甲基(1.36%);添加甘氨酸的发酵液中产生了八甲基环四硅氧烷(1.42%)、苯酚(1.05%)、六氢吡咯并[1,2-a]吡嗪-1,4-二酮(30.24%);添加色氨酸的发酵液中产生了二甲基亚硝胺(0.51%)、乙二醇丁醚(0.88%)、胍(1.13%)、苯甲醇(4.89%)、脱氢甲瓦龙酸内酯(1.02%)、4, 4-二甲基庚烷(1.34%)、3-吡啶, 4-氨基-2-乙基-6-甲基(2.70%)、3,5,7,8-四氢化-4,6-哌啶二酮(2.90%)、脱氧精胍菌素(1.5%)。

表3 不同生长因子条件下的空白培养基成分GC-MS分析
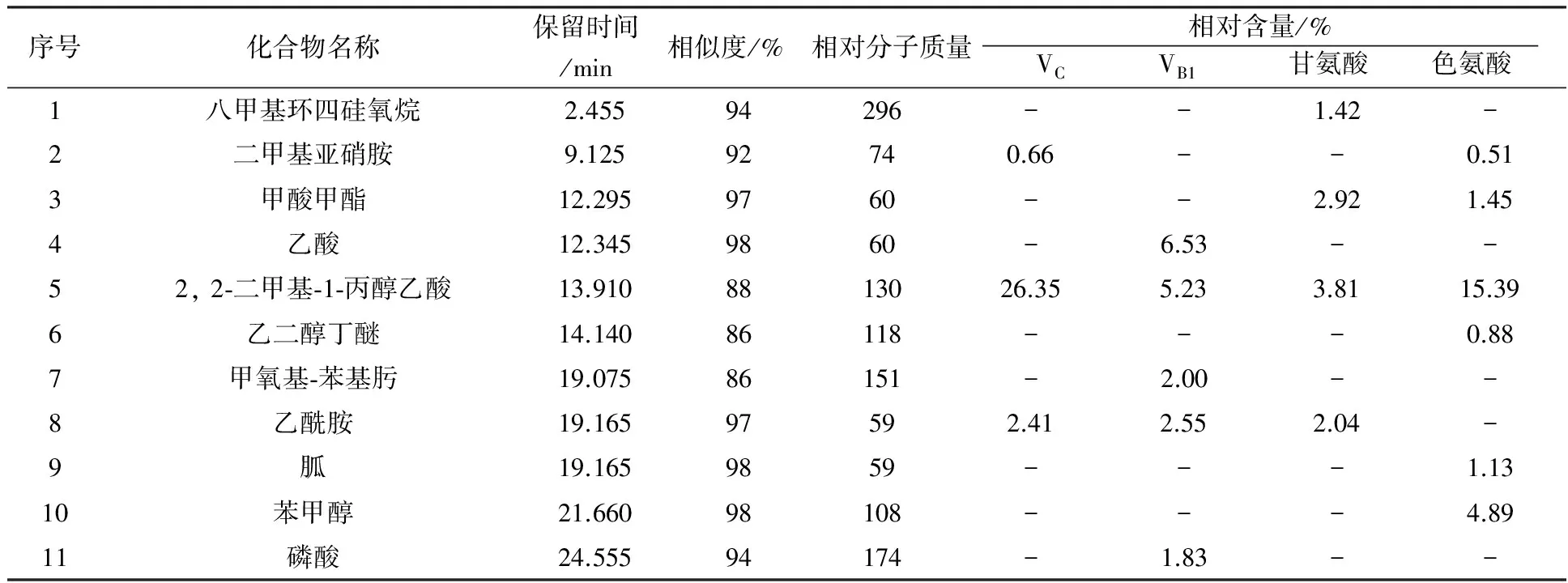
表4 不同生长因子条件下的空白培养基成分GC-MS分析
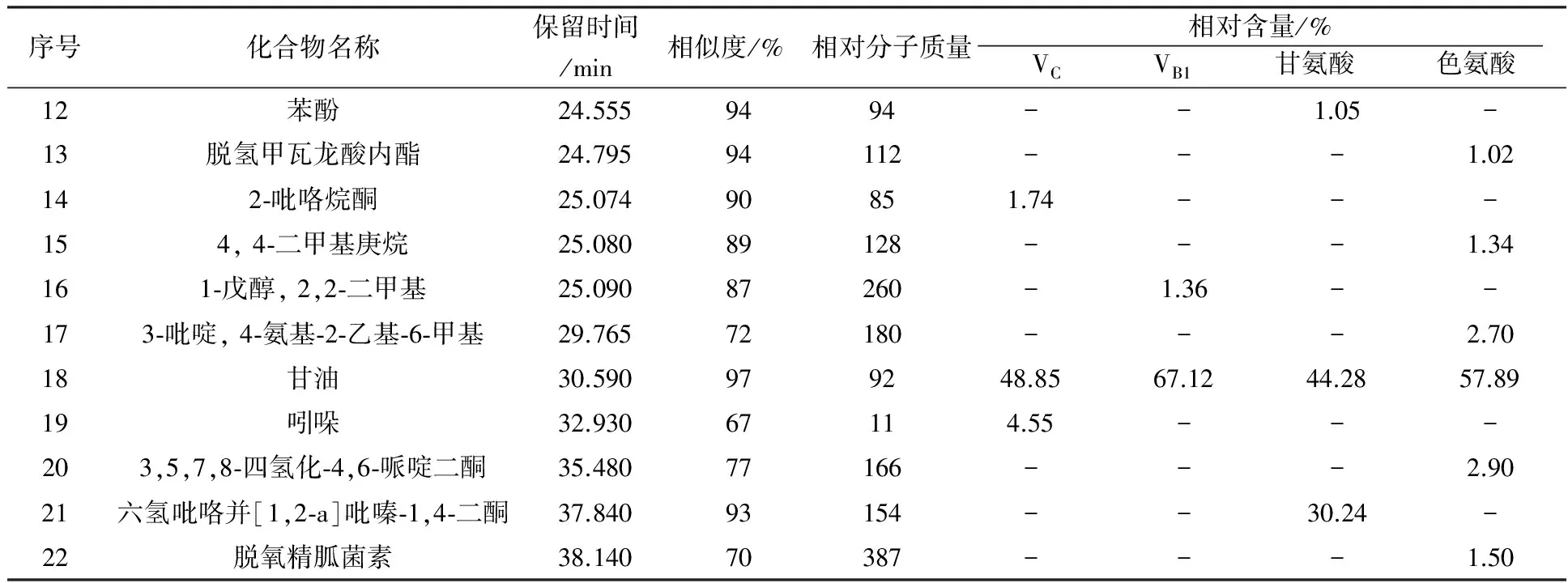
续表4
3 讨 论
微生物能够产生结构新颖、活性多样的次级代谢产物,是天然产物的重要来源之一,且不同的产物往往是经过不同的代谢途径产生的[29]。例如,醛类物质主要是通过不饱和脂肪酸的氧化、脂质氢过氧化物和过氧化物的分解及氨基酸的降解[30]途径形成的,酸类物质主要由脂肪酶水解途径,醇脱氢酶和醛脱氢酶转化途径和糖类物质合成脂肪酸等代谢途径形成,醇类化合物主要由糖酵解、脂肪酸经脂肪氧化作用以及氨基酸分解作用形成[31]。
本研究采用GC-MS分析方法检测了不同碳源和生长因子条件下粒毛盘菌产生的挥发性次级代谢产物,发现不同碳源条件下,粒毛盘菌YM406发酵液中产生的代谢产物种类与含量具有一定差异。以葡萄糖为碳源时,发酵液主要产生醇类(41.70%)、酸类(17.01%)、烷烃类(10.23%)和醚类(10.95%)化合物;以蔗糖为碳源时,发酵液主要产生醇类(20.52%)、酸类(29.00%)和烷烃类(9.46%)化合物;以淀粉为碳源时,发酵液主要产生醇类(50.52%)、酸类(11.09%)、胺类(11.16%)化合物,同时,其发酵液中检测到的挥发性代谢产物种类最多,为10种,且该组中醇类物质相对含量最高,说明以淀粉为碳源时利于粒毛盘菌产醇类物质,而Loughlin Gethins等[32]发现葡萄糖、果糖和乳糖3种碳源对马克斯克鲁维酵母的杂醇产量无显著影响,同时在3组发酵液中发现了一些有应用价值的化合物,如甘油、三氯甲烷、甲酸甲酯,吡咯和尿素,六氢吡咯并[1,2-a]吡嗪-1,4-二酮等。甘油和尿素是重要的化工原料,在医学、工业和商业领域等诸多方面具有广泛的应用[33];甲酸甲酯是极具应用价值的化学品和油品添加剂[34];三氯甲烷和吡咯衍生物是重要的有机合成原料。
不同生长因子条件下的粒毛盘菌YM406发酵液中产生的代谢产物种类与含量也具有一定差异。添加VC为生长因子的发酵液主要产生醇类(48.85%)和酸类(26.35%)化合物;添加VB1为生长因子的发酵液主要产生醇类(67.12%)和酸类(11.76%)化合物;添加甘氨酸为生长因子的发酵液主要产生醇类(44.28%)和酮类(30.24%)化合物;添加色氨酸为生长因子的发酵液主要产生醇类(62.78%)和酸类(15.39%)化合物,同时,其发酵液中检测到的挥发性代谢产物种类最多,为12种。由此可见,添加VB1为生长因子的发酵液产生的醇类物质最多。在4组发酵液中检测出乙酰胺、胍、八甲基环四硅氧烷、乙二醇丁醚、苯甲醇、脱氢甲瓦龙酸内酯和脱氧精胍菌素等具有应用价值的化合物。其中乙酰胺是许多有机物和无机物的优良溶剂,广泛用于各种工业产品中;八甲基环四硅氧烷是广泛应用于有机硅行业的化合物;胍、乙二醇丁醚和苯甲醇均广泛用于工业化学品生产中。脱氧精胍菌素是一种新型免疫抑制剂,可以有效地阻止移植器官排异,逆转急性排异反应,且副反应少,毒性低[35],应用前景非常广阔。
本研究表明,通过改变粒毛盘菌培养基的成分,可增加其代谢产物的多样性,为粒毛盘菌以及其他微生物代谢产物及其应用提供了科学依据。然而本研究只是对1株粒毛盘菌代谢产物中挥发性成分进行了初步分析,粒毛盘菌能产生多少种代谢产物,不同粒毛盘菌代谢产物种类如何,将有待采用更多的分析方法(如LC-MS、代谢组学等)以及更多的菌种(株)进行比较分析。
[1] 武临专,洪斌.微生物药物合成生物学研究进展[J].药学学报,2013,48(2):155-160.
[2] Hung R,Lee S,Bennett JW.Fungal volatile organic compounds and their role in ecosystems[J].Applied Microbiology & Biotechnology,2015,99(8):3395-3405.
[3] 杨宁.固态培养法增加微生物次级代谢产物的多样性[D].大连:大连理工大学,2013.
[4] Berdy J.Bioactive microbial metabolites[J].The Journal of antibiotics,2005,58(1):1-26.
[5] 王开梅,张亚妮,张志刚,等.提高微生物次生代谢产物多样性的策略[J].湖北农业科学,2010,49(12):3207-3210.
[6] 韦洪娟,林贞建,李德海,等.单菌多次级代谢产物方法及其在微生物代谢产物研究中的应用[J].微生物学报,2010,(6):701-709.
[7] 方祥,胡文峰,张辉华.不同发酵条件对乳杆菌 O-2 菌株代谢产物的影响[J].中国微生态学杂志,2004,16(1):15-17.
[8] 杨帆,王和玉,姚翠屏,等.不同工艺条件下枯草芽孢杆菌代谢产物分析对比[J].酿酒科技,2010,(1):104-106.
[9] 祁宏伟,闫晓刚,于维,等.不同培养条件对乳酸菌发酵代谢产物 CLA 的影响[J].安徽农学通报,2011,17(21):38-40.
[10] Sadeghy B, Hatami N.The effect of different conditions of temperature and acidity on antimicrobial metabolites ofStreptomycessp. isolates C-11 and C-26 againstF.subglutinans[J].Archives of PHytopathology & Plant Protection,2014,47(6):711-716.
[11] Shestivska V, P, Spaněl, Dryahina K, et al.Variability in the concentrations of volatile metabolites emitted by genotypically different strains ofPseudomonasaeruginosa[J].Journal of Applied Microbiology,2012,113(3):701-713.
[12] 余知和,庄文颖.中国热带粒毛盘菌属的物种多样性研究[J].生物多样性,2003,11(2):141-146.
[13] 陈辉.安徽粒毛盘菌属及其相关类群资源与多相分类研究[D]. 合肥:合肥工业大学,2007.
[14] 余知和,庄文颖.粒毛盘菌属及其相关属的分子系统学研究[J]. 菌物学报,2003,22(1):42-49.
[15] 叶明,庄文颖.粒毛盘菌属种间关系的分子系统学初探[J]. 菌物学报,2002,21(03):340-345.
[16] 叶明,阚清华,叶生梅,等.中国温带粒毛盘菌初探之一[J].生物学杂志,2004,21(5):18-21.
[17] 赵鹏,庄文颖.ITS作为粒毛盘菌属DNA条形码的探索(英文) [J].菌物学报, 2011,30(6):932-937.
[18] Stadler M,Anke H,Sterner O.New metabolites with nematicidal and antimicrobial activities from the ascomyceteLachnumpapyraceum (Karst.) Karst. VII. Structure determination of brominated lachnumon andmycorrhizinA derivatives[J].Journal of Antibiotics, 1995,48(2):158-161.
[19] Shan R, Stadler M, Sterner O, et al. New metabolites with nematicidal and antimicrobial activities from the ascomyceteLachnumpapyraceum (Karst.) Karst. VIII. Isolation, structure determination and biological activities of minor metabolites structurally related tomycorrhizinA.[J]. Journal of Antibiotics, 1996, 49(5):447-52.
[20] 陈吴西, 蔡敬民, 邱涛,等.LachnumYM328多糖发酵条件优化及抗氧化性[J]. 合肥工业大学学报(自然科学版), 2011, 34(2):300-303.
[21] Ye M, Chen W X, Qiu T, et al. Structural characterisation and anti-ageing activity of extracellular polysaccharide from a strain ofLachnumsp.[J]. Food Chemistry, 2012, 132(1):338-43.
[22] Chen T, Zhang M, Li J, et al. Structural characterization and hypoglycemic activity ofTrichosanthespeel polysaccharide[J]. LWT-Food Science and Technology, 2016, 70:55-62.
[23] Qiu T, Ma X, Ye M, et al. Purification, structure, lipid lowering and liver protecting effects of polysaccharide fromLachnumYM281[J]. Carbohydrate Polymers, 2013, 98(1):922-930.
[24] Song S, Yang L, Ye M, et al. Antioxidant activity of aLachnumYM226 melanin-iron complex and its influence on cytokine production in mice with iron deficiency anemia[J]. Food & Function, 2016, 7(3):1508-1514.
[25] 杜湛湛,张嫱,徐平,等.粒毛盘菌多糖纯化,硫酸酯化修饰及抗氧化活性评价[J].合肥工业大学学报:自然科学版,2016,39(1):134-139.
[26] 郭赓艺.粒毛盘菌 YM404 黑色素的纯化、结构及抗紫外辐射活性研究[D].合肥:合肥工业大学,2013.
[27] Lu Y, Ye M, Song S, et al. Isolation, Purification, and Anti-Aging Activity of Melanin fromLachnumsingerianum[J]. Applied Biochemistry & Biotechnology, 2014, 174(2):762-71.
[28] 邱涛. 粒毛盘菌YM281多糖的发酵、结构及其降脂利肝活性研究 [D].合肥:合肥工业大学,2012.
[29] 梁华正, 张燮, 饶军,等. 微生物挥发性代谢产物的产生途径及其质谱检测技术[J]. 中国生物工程杂志, 2008, 28(1):124-133.
[30] SINK J D. Lipid soluble components of meatflavors/odors and their biochemical origin[J]. Journal of the American Oil Chemists′ Society, 1973, 50(11): 470-474.
[31] 李莹,白凤翎,励建荣.发酵海产品中微生物形成挥发性代谢产物研究进展[J].食品科学,2015,36(15):255-259.
[32] Gethins L, Guneser O, Demirkol A, et al. Influence of carbon and nitrogen source on production of volatile fragrance and flavour metabolites by the yeastKluyveromycesmarxianus[J]. Yeast, 2015, 32(1):67-76.
[33] 宋如,钱仁渊,李成,等.甘油新用途研究进展[J].中国油脂,2008,33(5):40-44.
[34] 武建兵, 吴志伟, 王瑞义,等. 甲醇制备甲酸甲酯、甲缩醛和聚甲醛二甲醚的催化反应研究进展[J]. 燃料化学学报, 2015, 43(07):816-828.
[35] 程晓昆,王娅莉,张惠敏,等.反相离子对色谱法测定脱氧精胍菌素药物中的主成分[J]. 色谱,2012,30(12):1292-1294.
InitialStudyonMetabolitesofLachnumsp.withDifferentCarbonSources&GrowthFactors
SHI Fang1, ZHAO Mo-yuan1, 2, YANG Liu-qing1, HOU Guo-hua1, YE Ming1
(1.Schl.ofFoodSci. &Engin.,HefeiUni.ofTechnol.,Hefei230009; 2.FacultyofMed.,Nursing, &HealthSci.,MonashUni.,Australia3168)
Composition and difference of volatile components in fermentation metabolites ofLachnumYM406 with different carbon sources and growth factors was analyzed by GC-MS. The results showed that the metabolites are various with different carbon sources and growth factors, mainly including organic acids, amines, alkanes, esters, alcohols, pyrroles and other substances. There were respectively 7, 7, 10 kinds of volatile metabolites in fermentation broth with 20 g/L of glucose, sucrose, and starch as carbon source. Meanwhile, in fermentation broth added with 1mg/L VC, VB1, glycine, tryptophan as growth factors, there were 6, 7, 7, 12 kinds of volatile metabolites respectively. These results showed that the fermentation metabolites byLachnumYM406 under different culture conditions were fairly diverse, suggested that strain YM406 had important application value.
Lachnum; metabolites; GC-MS; carbon sources; growth factors
国家自然科学基金项目(31470146)
石芳 女,硕士研究生。研究方向为工业微生物学。E-mail:1024312029@qq.com
* 通讯作者。男,教授,博士生导师。主要从事食品微生物、微生物源活性物质的结构与功效等研究。E-mail:yeming123@sina.com
2016-08-08;
2016-08-30
Q939.97;TS201.2
A
1005-7021(2017)04-0053-07
10.3969/j.issn.1005-7021.2017.04.009
猜你喜欢
能源化工(2022年3期)2023-01-15 02:26:43
西南大学学报(自然科学版)(2022年12期)2022-12-20 02:32:14
中国食用菌(2020年8期)2020-11-08 13:41:50
科学Fans(2020年3期)2020-05-13 14:30:32
环境与发展(2019年10期)2019-11-28 14:25:24
课程教育研究·学法教法研究(2019年10期)2019-05-14 11:02:18
天然产物研究与开发(2018年4期)2018-05-07 06:47:53
生物技术通报(2018年4期)2018-03-31 17:16:11
中成药(2018年1期)2018-02-02 07:20:03
汽车文摘(2014年8期)2014-12-16 01:54:28
