
碱胁迫下燕麦幼苗对外源过氧化氢的生理响应
2017-11-08 07:36刘建新王金成刘秀丽李博萍
麦类作物学报 2017年10期
刘建新,王金成,刘秀丽,李博萍
(甘肃省高校陇东生物资源保护与利用省级重点实验室/陇东学院生命科学与技术学院,甘肃庆阳 745000)
碱胁迫下燕麦幼苗对外源过氧化氢的生理响应
刘建新,王金成,刘秀丽,李博萍
(甘肃省高校陇东生物资源保护与利用省级重点实验室/陇东学院生命科学与技术学院,甘肃庆阳 745000)
为探讨过氧化氢(H2O2)对碱胁迫下植物的生理调节作用,以燕麦品种定莜6号的幼苗为材料,采用沙培方法,设置0、50和150 μmol·L-1三个H2O2浓度水平,研究了外源H2O2对100 mmol·L-1NaHCO3胁迫下燕麦叶片活性氧代谢、渗透溶质积累和幼苗生长的影响。结果表明,NaHCO3胁迫下,与不添加H2O2处理(0 μmol·L-1)相比,50和150 μmol·L-1H2O2处理均不同程度降低了燕麦叶片超氧阴离子、H2O2、丙二醛、可溶性糖含量和细胞质膜的相对透性,明显提高可溶性蛋白质、脯氨酸和类胡萝卜素含量及叶绿素a/b值。 NaHCO3胁迫下,50 μmol·L-1H2O2处理的燕麦叶片超氧化物歧化酶(SOD)活性和谷胱甘肽(GSH)含量下降,过氧化氢酶(CAT)、过氧化物酶(POD)、抗坏血酸过氧化物酶(APX)活性及抗坏血酸(AsA)含量提高;150 μmol·L-1H2O2处理则提高了SOD、POD、APX活性和AsA含量,降低了CAT活性,对GSH含量无明显影响。50 μmol·L-1H2O2显著缓解了NaHCO3胁迫对燕麦幼苗生长的抑制作用,150 μmol·L-1H2O2的缓解效应不明显。
过氧化氢;燕麦;碱胁迫;活性氧代谢;渗透调节物质

过氧化氢(hydrogen peroxide, H2O2)是生物细胞应答各种逆境胁迫的一种信号分子[8],它参与调控脱落酸诱导的拟南芥(Arabidopsisthaliana)气孔关闭过程[9]。外源H2O2处理能够缓解干旱胁迫诱导的活性氧积累对黄瓜(Cucumissativus)叶绿体膜的伤害[10];提高玉米(Zeamays)的耐盐性[11];并通过调节多磷酸肌醇激酶基因 VvIPK2表达,参与葡萄(Vitis)低温胁迫响应[12];提高水稻(Oryzasativa)镉胁迫耐性[13]。外源H2O2能否提高植物耐碱性?目前鲜见这方面的报道。本研究以燕麦品种定莜6号的幼苗为材料,检测施加H2O2后其叶片活性氧代谢、渗透溶质积累等生理指标的变化,探讨碱胁迫下燕麦幼苗对外源H2O2的生理响应,以期为燕麦耐碱栽培提供理论依据。
1 材料与方法
1.1 材料培养与处理
1.1.1 材料培养
实验于2015年4-8月在甘肃省高校陇东生物资源保护与利用省级重点实验室日光温室进行。供试燕麦品种为定莜6号,种子由甘肃省定西市旱作农业科研推广中心提供。选取饱满、大小一致的燕麦种子,用1%次氯酸钠表面消毒15 min,25 ℃催芽,露白后播种在装满珍珠岩的塑料盆(口径20 cm,高14 cm)中,浇水后置日光温室培养,昼/夜温度为(22±5) ℃/(16±6) ℃,相对湿度为65%±15%,光照强度420~680 μmol·m-2·s-1。2015年5月二叶一心期每盆留苗约200株,当幼苗长至三叶一心期时进行实验处理。
1.1.2 实验设计
实验设6个处理,分别为Hoagland营养液(CK)、Hoagland营养液 + 100 mmol·L-1NaHCO3(T1)、 Hoagland营养液 + 100 mmol·L-1NaHCO3+ 50 μmol·L-1H2O2(T2)、Hoagland营养液 + 100 mmol·L-1NaHCO3+ 150 μmol·L-1H2O2(T3)、Hoagland营养液 + 50 μmol·L-1H2O2(T4)、 Hoagland营养液 + 150 μmol·L-1H2O2(T5)。用以上处理液于每天19:00浇灌幼苗根部,每盆浇灌量约300 mL。为了避免盐碱冲击幼苗,NaHCO3胁迫浓度每天递增25 mmol·L-1,4 d后浓度达到100 mmol·L-1时,记为实验处理正式开始。处理7 d后取幼苗倒数第1、第2片功能叶,液氮速冻后-70 ℃保存,用于测定相关生理指标。每处理取50株完整幼苗洗净拭干后称量鲜重。每5盆为1个重复,重复3次,随机区组排列。
1.2 测定指标与方法
采用高俊凤[14]的方法测定超氧阴离子含量和细胞质膜相对透性;参照Sergiev等[15]的方法测定H2O2含量;丙二醛(MDA)含量测定采用李合生[16]的方法;超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)和抗坏血酸过氧化物酶(APX)活性及抗坏血酸(AsA)和谷胱甘肽(GSH)含量采用陈建勋和王晓峰[17]的方法测定;可溶性糖、可溶性蛋白质、游离氨基酸和脯氨酸含量测定分别采用李合生[16]的蒽酮比色法、考马斯亮蓝法、茚三酮染色法和磺基水杨酸法;光合色素含量按高俊凤[14]的方法测定。每个指标重复测定3次,计算平均值。
1.3 数据统计分析
采用Excel 2003整理数据及绘图,用SPSS 20.0方差分析和Duncan法多重比较(P<0.05)。
2 结果与分析
2.1碱胁迫下燕麦幼苗叶片活性氧产生和膜脂过氧化对外源H2O2的响应
与CK相比,NaHCO3胁迫(T1处理)下燕麦幼苗叶片超氧阴离子、H2O2、MDA含量和质膜相对透性均显著提高(表1)。NaHCO3胁迫下,与T1处理相比,添加50 μmol·L-1H2O2(T2处理)后,幼苗超氧阴离子含量无显著变化,但H2O2、MDA含量和质膜相对透性均明显下降;添加150 μmol·L-1H2O2(T3处理)后,幼苗MDA含量和质膜相对透性与T1处理无显著差异,而超氧阴离子和H2O2含量明显降低。单施150 μmol·L-1H2O2(T5处理)使燕麦幼苗叶片MDA含量较CK明显提高,而单施50 μmol·L-1H2O2(T4处理)对超氧阴离子、H2O2、MDA含量和相对质膜透性均无显著影响。这说明外源H2O2可在一定程度上抑制NaHCO3胁迫下燕麦幼苗中活性氧的产生,降低膜脂过氧化水平,减轻质膜的伤害。
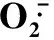
表1 外源H2O2对碱胁迫下燕麦幼苗叶片超氧阴离子、内源过氧化氢、丙二醛和相对质膜透性的影响Table 1 Effect of exogenous H2O2 on , endogenous H2O2, malondialdehyde contents and relative plasma membrane permeability in oat seedling leaves under alkali stress
同列不同字母表示处理间5%水平差异显著。表2同。
Different letters within the same column indicate significant difference among the treatments at 5% level. The same in table 2.
2.2碱胁迫下燕麦幼苗叶片活性氧清除系统对外源H2O2的响应
与CK相比,NaHCO3胁迫(T1处理)显著提高了燕麦幼苗叶片SOD、APX活性和GSH含量,明显降低CAT和POD活性,对AsA含量影响不显著(图1)。NaHCO3胁迫下,与T1处理相比,添加50 μmol·L-1H2O2(T2处理)后,幼苗叶片SOD活性和GSH含量显著下降,而CAT、POD、APX活性和AsA含量明显提高;添加150 μmol·L-1H2O2(T3处理)后,幼苗叶片SOD、POD、APX活性和AsA含量显著提高,CAT活性明显下降,GSH含量则变化不大。单施50 μmol·L-1H2O2处理(T4)的SOD活性与CK无显著差异,CAT、POD活性和GSH含量明显下降,APX活性和AsA含量显著升高;单施150 μmol·L-1H2O2处理(T5)使SOD活性和AsA含量明显升高,CAT、POD活性和GSH含量显著下降,APX活性则与CK差异不明显。这表明,外源H2O2可提高NaHCO3胁迫下燕麦幼苗的活性氧清除能力,有助于减轻碱胁迫的伤害。
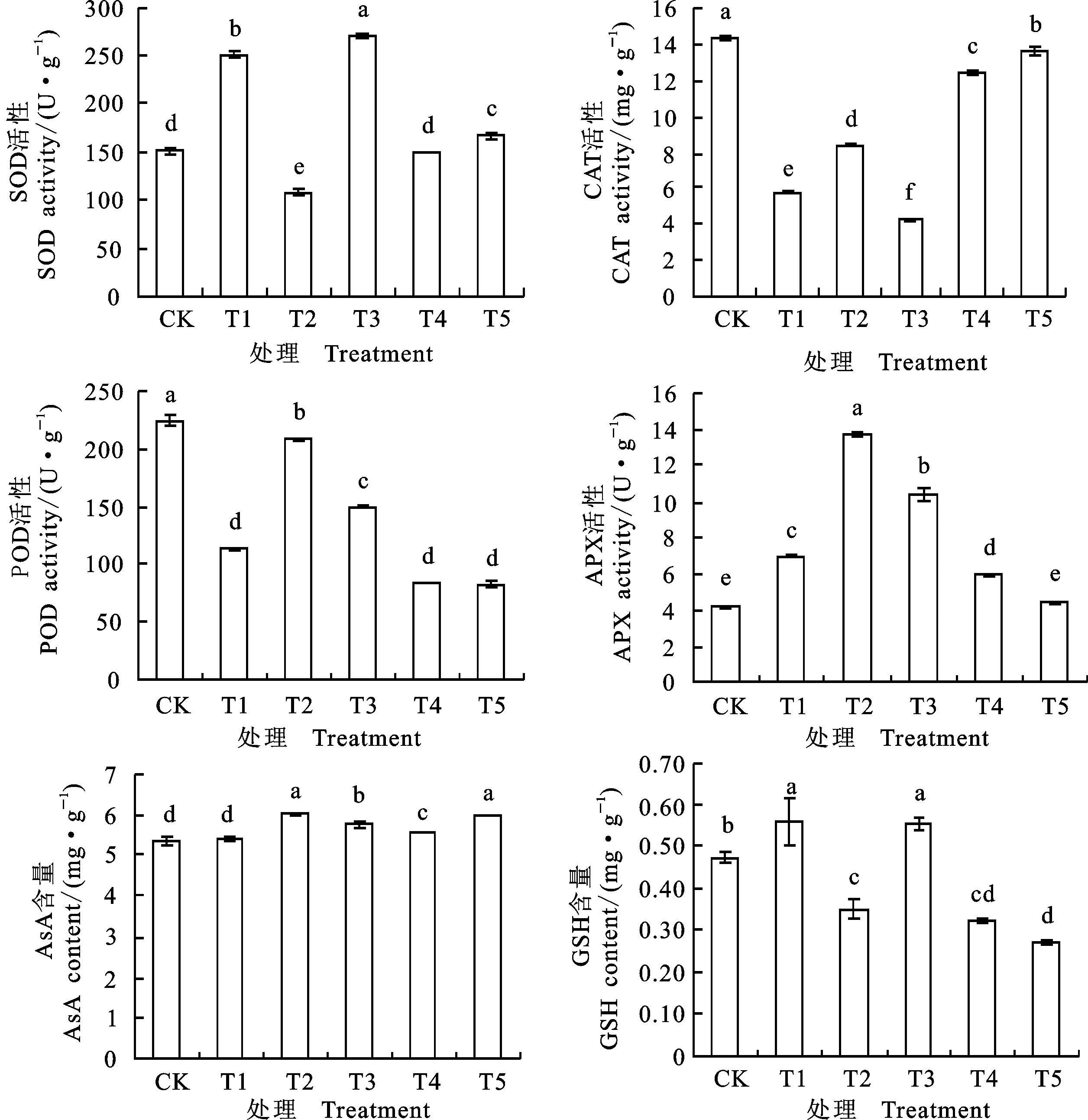
2.3碱胁迫下燕麦幼苗叶片渗透调节物质对外源H2O2的响应
与CK相比,NaHCO3胁迫(T1处理)显著提高了燕麦叶片可溶性糖和脯氨酸含量,降低可溶性蛋白质含量,对游离氨基酸含量影响不大(图2)。NaHCO3胁迫下,与T1处理相比,添加50 μmol·L-1H2O2(T2处理)显著降低叶片可溶性糖和游离氨基酸含量,提高了可溶性蛋白质和脯氨酸含量;添加150 μmol·L-1H2O2(T3处理)也使可溶性糖含量明显下降,但下降幅度明显小于T2处理,可溶性蛋白质、游离氨基酸和脯氨酸含量均显著提高,其中可溶性蛋白质和脯氨酸的增幅小于T2处理。单施H2O2处理(T4和T5)均不同程度提高了幼苗叶片可溶性糖、可溶性蛋白质和脯氨酸含量,游离氨基酸含量则明显下降,其中T4处理的下降幅度大于T5处理。由此可见,外源H2O2对NaHCO3胁迫下燕麦幼苗的渗透调节有一定的改善效应。
2.4碱胁迫下燕麦幼苗叶片光合色素含量对外源H2O2的响应
与CK相比,NaHCO3胁迫(T1处理)提高了燕麦叶片叶绿素a、叶绿素b、叶绿素(a+b)和类胡萝卜素含量,但对叶绿素a/b值无明显影响(表2)。NaHCO3胁迫下,与T1处理相比,添加50 μmol·L-1H2O2(T2处理)后叶绿素a、叶绿素(a+b)含量没有显著改变,叶绿素b含量却明显下降,叶绿素a/b值和类胡萝卜素含量明显提高;添加150 μmol·L-1H2O2(T3处理)后光合色素的变化与T2处理相似,但叶绿素b的降幅和叶绿素a/b的增幅更大,类胡萝卜素增幅则较小。单施H2O2(T4和T5处理)均提高了燕麦叶片叶绿素a、叶绿素b、叶绿素(a+b)、类胡萝卜素含量和叶绿素a/b值。这说明外源H2O2主要影响了叶绿素b和类胡萝卜素的合成。
2.5碱胁迫下燕麦幼苗生物量对外源H2O2的响应
与CK相比,NaHCO3胁迫(T1处理)显著降低了燕麦幼苗鲜重,下降幅度为30.7%(图3)。NaHCO3胁迫下,添加50 μmol·L-1H2O2(T2处理)后幼苗鲜重较T1处理显著升高,说明该处理在一定程度上缓解了NaHCO3胁迫对燕麦幼苗生长的抑制作用,但添加150 μmol·L-1H2O2处理(T3)的燕麦幼苗鲜重与T1处理无明显差异。单施50 μmol·L-1H2O2(T4处理)能显著提高燕麦幼苗鲜重,而单施用150 μmol·L-1H2O2(T5处理)的燕麦幼苗鲜重与CK无明显差异。
图柱上不同字母表示处理间在5%水平差异显著。图2同。
Different letters above the columns indicate significant difference among the different treatments at 5% level. The same in figure 2.
图1外源H2O2对碱胁迫下燕麦幼苗叶片活性氧清除系统的影响
Fig.1EffectofexogenousH2O2onactiveoxygenscavengingsysteminoatseedlingleavesunderalkalistress
图2 外源H2O2对碱胁迫下燕麦幼苗叶片渗透调节物质含量的影响

表2 外源H2O2对碱胁迫下燕麦幼苗叶片光合色素含量的影响Table 2 Effect of exogenous H2O2 on photosynthetic pigment content in oat seedling leaves under alkali stress
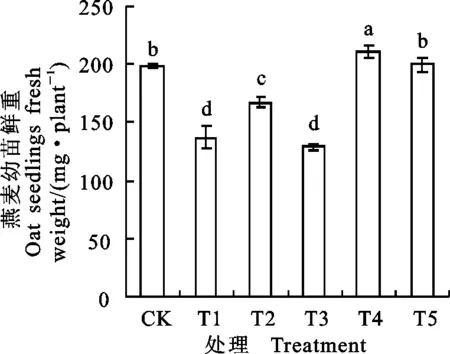
图3 不同处理下燕麦幼苗鲜重
3 讨 论
3.1外源H2O2对燕麦活性氧代谢响应碱胁迫的调节
郭慧娟等[18]研究表明,随着碱胁迫处理液pH的升高,多年生黑麦草(Loliumperenne)幼苗膜脂过氧化水平也显著升高。Gong等[3]指出,与NaCl胁迫相比,NaHCO3短期胁迫诱导了番茄(Lycopersiconesculentum)更高的活性氧积累和抗氧化系统活性及脂质过氧化,而较长时间胁迫则产生相反结果。本实验表明,外源H2O2能够有效抑制NaHCO3胁迫下燕麦幼苗超氧阴离子和H2O2含量的提高,减缓膜脂过氧化产物MDA的积累和质膜透性的增大。这与外源H2O2能够缓解干旱胁迫诱导的活性氧积累对黄瓜叶绿体膜伤害[10]的结果一致。这说明外源H2O2能够减轻碱胁迫诱导的燕麦膜脂过氧化伤害。
SOD催化超氧阴离子发生歧化反应生成H2O2和O2,CAT、POD、APX等协同作用进一步催化H2O2反应生成H2O,而AsA和GSH直接或通过AsA-GSH循环途径参与了H2O2的清除[19]。在NaHCO3胁迫下黄瓜幼苗SOD、CAT、POD和APX活性显著升高[2];黑麦草幼苗SOD活性随碱处理液pH升高而先升后降,CAT活性逐步递减,POD活性呈递增趋势[18]。本实验表明,燕麦幼苗通过提高叶片SOD、APX活性和GSH含量增强了NaHCO3胁迫下的抗氧化能力,但不同浓度的外源H2O2效应有所差异。施用50 μmol·L-1H2O2明显降低了NaHCO3胁迫下燕麦叶片的SOD活性和GSH含量,提高了CAT、POD、APX活性及AsA含量;施用150 μmol·L-1H2O2则提高了SOD、POD和APX活性及AsA含量,降低了CAT活性,GSH含量却没有变化。这与外源H2O2预处理可通过提高SOD、CAT、APX、GSH-PX活性来降低热胁迫下黄瓜叶片活性氧和MDA含量的结果类似[21]。
3.2外源H2O2对碱胁迫下燕麦幼苗渗透调节物质含量的影响
张波等[22]研究表明,外源H2O2能够通过提高可溶性糖和GSH含量来增强小麦(Triticumaestivuml)幼苗的耐盐性。本研究表明,外施50和150 μmol·L-1H2O2显著提高了NaHCO3胁迫下燕麦叶片的可溶性蛋白和脯氨酸含量,却降低了可溶性糖含量,50 μmol·L-1H2O2还降低了NaHCO3胁迫下燕麦的游离氨基酸含量,150 μmol·L-1H2O2作用则相反。这与Wahid等[19]的H2O2预处理能够促进盐胁迫下小麦幼苗胁迫蛋白表达的结果类似。说明外源H2O2对可溶性蛋白和脯氨酸积累的促进作用可能是其增强燕麦在碱胁迫下渗透调节能力的重要原因。H2O2提高碱胁迫下燕麦可溶性蛋白含量的原因可能是其抑制依赖光合电子传递链活性氧的产生,从而阻止了活性氧诱导的蛋白酶活性升高对可溶性蛋白的降解[23]或诱导新胁迫蛋白形成所致[19]。脯氨酸水平的提高可能与H2O2能够激活燕麦幼苗脯氨酸合成的谷氨酸途径和鸟氨酸途径、抑制脯氨酸的降解有关[24]。但外源H2O2对燕麦可溶性糖及游离氨基酸含量的影响机理还需进一步深入探讨。
3.3外源H2O2对碱胁迫下燕麦幼苗光合色素含量和生物量的影响
在筛选NaHCO3和H2O2浓度的预备实验中,用100 mmol·L-1NaHCO3处理2叶1心期燕麦幼苗7 d后,植株鲜重显著下降56.3%。用 0~1 000 μmol ·L-1H2O2处理2叶1心期燕麦幼苗12 d后,25~300 μmol ·L-1H2O2处理下鲜重显著提高,500和1 000 μmol ·L-1H2O2处理下鲜重变化不大或明显下降, 其中50和150 μmol ·L-1H2O2处理下鲜重最大。本实验中,外施50和150 μmol ·L-1H2O2对100 mmol·L-1NaHCO3胁迫下燕麦叶绿素含量没有影响,却提高了叶绿素a/b值和类胡萝卜素含量;50 μmol·L-1H2O2处理显著缓解了100 mmol·L-1NaHCO3胁迫对燕麦幼苗生长的抑制作用,但100 mmol·L-1NaHCO3胁迫下添加150 μmol·L-1H2O2后燕麦幼苗鲜重变化不明显。这说明适宜浓度H2O2才可以缓解碱胁迫对燕麦生长的抑制作用。叶绿素a/b值反映类囊体的垛叠程度,垛叠程度越高,植株抵御环境胁迫的能力越强,类胡萝卜素除辅助光能吸收外,还可吸收剩余光能和淬灭活性氧[20]。外源H2O2对碱胁迫下燕麦叶绿素a/b值和类胡萝卜素含量的提升作用,可能促进了过剩激发能的耗散和活性氧的淬灭,进而缓解了碱胁迫对燕麦生长的抑制。这与外源H2O2能够减轻受旱黄瓜叶绿体膜损伤[10]及通过提高叶绿素和可溶性糖含量增强小麦耐盐性[22]的结论一致。
综上所述,外源H2O2可缓解NaHCO3胁迫对燕麦幼苗的氧化伤害及生长抑制,增强燕麦植株的耐碱性。这种作用一方面可能归因于其诱导了高效的活性氧清除系统;另一方面,可能归因于其调节了渗透平衡,提高了叶绿素a/b值和类胡萝卜素含量。
[1] 卢培娜,刘景辉,李 倩,等.盐碱地不同燕麦品种的品质及产量比较[J].麦类作物学报,2016,36(11):1510.
LU P N,LIU J H,LI Q,etal.Comparison of quality and yield of different oat varieties in saline-alkali land [J].JournalofTriticeaeCrops,2016,36(11):1510.
[2] 颜 宏,赵 伟,盛艳敏,等.碱胁迫对羊草和向日葵的影响[J].应用生态学报,2005,16(8):1497.
YAN H,ZHAO W,SHENG Y M,etal.Effects of alkali-stress onAneurolepidiumchinenseandHelianthusannuus[J].ChineseJournalofAppliedEcology,2005,16(8):1497.
[3] GONG B,WEN D,VANDENLANGENBERG K,etal.Comparative effects of NaCl and NaHCO3stress on photosynthetic parameters,nutrient metabolism,and the antioxidant system in tomato leaves [J].ScientiaHorticulturae,2013,157(3):1.
[4] YANG C,SHI D,WANG D L.Comparative effects of salt stress and alkali stress on growth,osmotic adjustment and ionic balance of an alkali resistant halophyteSuaedaglauca(Bge.) [J].PlantGrowthRegulation,2008,56(2):179.
[5] LI C Y,FANG B,YANG C W,etal.Effects of various salt-alkaline mixed stresses on the state of mineral elements in nutrient solutions and the growth of alkali resistant halophyteChlorisvirgata[J].JournalofPlantNutrition,2009,174(32):1137.
[6] DRZIKOVA B,DONGOWSKI G,GEBHARDT E.Dietary fibre-rich oat-based products affect serum lipids,microbiota,formation of short-chain fatty acids and steroids in rats [J].BritishJournalofNutrition,2005,94(6):1012.
[7] 刘建新,王金成,王瑞娟,等.燕麦幼苗对盐胁迫的响应及过氧化氢对响应的调节[J].生态学杂志,2015,34(9):2506.
LIU J X,WANG J C,WANG R J,etal.Response ofAvenanudaL.seedlings to salt stress and the modulation of hydrogen peroxide [J].ChineseJournalofEcology,2015,34(9):2506.
[8] DAT J F,VANDENDEDE F,VRANOVA E,etal.Dual action of the active oxygen species during plant stress response [J].CellularandMolecularLifeSciences,2000,57(5):779.
[9] BRIGHT J,DESIKAN R,HANCOCK J T,etal.ABA-induced NO generation and stomatal closure inArabidopsisare dependent on H2O2synthesis [J].PlantJournal,2006,45(1):113.
[10] 刘忠静,郭延奎,林少航,等.外源过氧化氢对干旱胁迫下温室黄瓜叶绿体超微结构和抗氧化酶的影响[J].园艺学报,2009,36(8):1140.
LIU Z J,GUO Y K,LIN S H,etal.Effects of exogenous hydrogen peroxide on ultra structure of chloroplastsand activities of antioxidant enzymes in greenhouse-ecotype cucumber under drought stress [J].ActaHorticulturaeSinica,2009,36(8):1140.
[11] ANDRE-DIAS DE A N,JOSE-TARQUINIO P,JOAQUIM E F,etal.Hydrogen peroxide pre-treatment induces salt stress acclimation in maize plants [J].JournalofPlantPhysiology,2005,162(10):1114.
[12] 李希东,侯丽霞,刘 新,等.H2O2与葡萄 VvIPK2基因表达及其低温胁迫响应的关系[J].园艺学报,2011,38(6):1052.
LI X D,HOU L X,LIU X,etal.Involvement of H2O2in regulating the expression of VvIPK2 in response to low temperature stress in leaves of vitis [J].ActaHorticulturaeSinica,2011,38(6):1052.
[13] 白晓娟,刘丽娟,张春华,等.H2O2预处理对不同水稻品种Cd耐性的影响[J].中国水稻科学,2010,24(4):391.
BAI X J,LIU L J,ZHANG C H,etal.Effect of H2O2pretreatment on Cd tolerance of different rice cultivars [J].ChineseJournalofRiceScience,2010,24(4):391.
[14] 高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006:74-77,208-209,221-222.
GAO J F.Plant physiology experiment guide [M].Beijing:Higher Education Press,2006:74-77,208-209,221-222.
[15] SERGIEV I,ALEXIEVA V,KARANOV E.Effect of spermine,atrazine and combination between them on some endogenous protective systems and stress markers in plants [J].ComptesRendusdeI'AcademieBulgaredesSciences,1997,51(2):121.
[16] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:184-196,258-261.
LI H S.The Experiment Principle and Technique on Plant Physiology and Biochemistry [M].Beijing:Higher Education Press,2000:184-196,258-261.
[17] 陈建勋,王晓峰.植物生理学实验指导[M].广州:华南理工大学出版社,2002:68-77.
CHEN J X.Plant Physiology Experiment Instruction [M].Guangzhou:South China University of Technology Press,2006:68-77.
[18] 郭慧娟,胡 涛,傅金民.苏打碱胁迫对多年生黑麦草的生理影响[J].草业学报2012,21(1):118.
GUO H J,HU T,FU J M.Effects of saline sodic stress on growth and physiological responses ofLoliumperenne[J].ActaPrataculturaeSinica,2012,21(1):118.
[19] WAHID A,PERVEEN M,GELANI S,etal.Pretreatment of seed with H2O2improves salt tolerance of wheat seedlings by alleviation of oxidative damage and expression of stress protein [J].JournalofPlantPhysiology,2007,164(3):283.
[20] 苗 丽,巩 彪,聂文婧,等.外源IAA对NaHCO3胁迫下黄瓜幼苗光合特性和抗氧化系统的影响[J].植物生理学报,2014,50(6):765.
MIAO L,GONG B,NIE W J,etal.Effects of exogenous IAA on photosynthetic characteristics and antioxidative system inCucumissativusseedlings under NaHCO3stress [J].PlantPhysiologyJournal,2014,50(6):765.
[21] GAO Y,GUO Y K,LIN S H,etal.Hydrogen peroxide pretreatment alters the activity of antioxidant enzymes and protects chloroplast ultrastructure in heat-stressed cucumber leaves [J].ScientiaHorticulturae,2010,126(1):20.
[22] 张 波,张怀刚.外源H2O2对小麦幼苗耐盐性的调节作用[J].西北植物学报,2007,27(12):2491.
ZHANG B,ZHANG H G.Regulation of exogenous hydrogen peroxide on wheat seedling salinity tolerance [J].ActaBotanicaBoreali-OccidentaliaSinica,2007,27(12):2491.
[23] GARCA-FERRIS C,MORENO J.Redox regulation of enzymatic activity and proteolytic susceptibility of ribulose-1,5-bisphosphate carboxylase/oxygenase fromEuglenagracilis[J].PhotosynthesisResearch,1993,35(1):55.
[24] 刘建新,王金成,刘秀丽,等.外源H2O2对混合盐碱胁迫下燕麦幼苗叶片脯氨酸积累和代谢途径的影响[J].植物研究,2016,36(3):427.
LIU J X,WANG J C,LIU X L,etal.Effect of exogenous H2O2on proline accumulation and metabolic pathway in leaves of oat seedlings under complex saline-alkali stress [J].BulletinofBotanicalResearch,2016,36(3):427.
EffectofExogenousHydrogenPeroxideonPhysiologicalResponsesofAvenanudaL.SeedlingsunderAlkaliStress
LIUJianxin,WANGJincheng,LIUXiuli,LIBoping
(College of Life Science and Technology, Longdong University;Provincial Key Laboratory for Protection andUtilization of Longdong Bio-resources in Gansu Province,Qingyang,Gansu 741500,China)
To understand the regulative effect of H2O2, a signaling molecule on the physiological response of oat(AvenanudaL.) seedlings under alkali stress, in the present study, a sand culture experiment was conducted to investigate the effect of H2O2on leaves active oxygen metabolism, osmotica accumulation and growth of ‘Dingyou 6’ oat seedlings under 100 mmol·L-1NaHCO3stress. The results showed that under the NaHCO3stress, the application of 50 and 150 μmol·L-1H2O2both could effectively decrease the contents of superoxide anion, H2O2, malondialdehyde, soluble sugar and plasma membrane permeability in leaves, while increased the contents of soluble protein, proline, carotenoid and chlorophyll a/b values. The application of 50 μmol·L-1H2O2significantly alleviated the inhibitory effect of NaHCO3stress on the growth of oat seedlings, decreased superoxide dismutase(SOD) activity and glutathione(GSH) content, and increased activities of catalase(CAT), peroxidase(POD), ascorbate peroxidase(APX) and ascorbic acid(AsA) content in the leaves of the seedlings under NaHCO3stress. While 150 μmol·L-1H2O2had no significantly alleviate growth inhibitory of oat seedlings induced by NaHCO3stress, but it increased SOD, POD, APX activities and AsA content, and decreased CAT activity, but no significant difference was observed in the GSH content compared with NaHCO3stress.
H2O2; Oat; Alkali stress; Active oxygen metabolism; Osmotic adjustment substance
时间:2017-10-11
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20171011.1602.030.html
2017-03-11
2017-04-21
甘肃省庆阳市科技计划项目(kz2014-19)
E-mail:liujx1964@163.com
刘建新(E-mail: liujx1964@163.com)
S512.1;S311
A
1009-1041(2017)10-1383-07
猜你喜欢
现代农业科技(2022年5期)2022-12-14
中老年保健(2022年5期)2022-11-25
中老年保健(2022年3期)2022-11-21
科学导报(2022年11期)2022-03-03
文萃报·周五版(2021年37期)2021-10-09
今日农业(2021年7期)2021-07-28
智慧健康(2020年9期)2020-12-03
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
延河(2017年7期)2017-07-19
