
六倍体(AnAnCnCnCoCo)与白菜型油菜杂交可交配性及后代菌核病抗性
2017-10-13 10:31李勤菲陈致富刘瑶梅家琴钱伟
中国农业科学 2017年1期
李勤菲,陈致富,刘瑶,梅家琴,钱伟
六倍体(AnAnCnCnCoCo)与白菜型油菜杂交可交配性及后代菌核病抗性
李勤菲1,2,陈致富2,刘瑶2,梅家琴2,钱伟2
(1西南大学园艺园林学院,重庆 400715;2西南大学农学与生物科技学院,重庆 400715)
【目的】利用亲本种遗传资源是改良甘蓝型油菜的重要手段。以甘蓝型油菜与菌核病抗性甘蓝杂交合成的六倍体为桥梁,与大量的白菜型油菜杂交,合成杂种,探索改良甘蓝型油菜菌核病抗性的策略。【方法】采用菌核病抗病甘蓝(C01)与甘蓝型油菜(中双9号)杂交合成六倍体,通过分析六倍体的育性、菌病抗性和减数分裂行为来分析其作为桥梁材料转移菌核病抗性的可能性;将六倍体与110份白菜型油菜杂交,通过考察杂种发育和可交配性来分析六倍体与白菜型油菜杂交的可行性;通过鉴定杂种的苗期表型特征、自交结实率及离体茎秆的菌核病抗性来分析杂种在改良甘蓝型油菜遗传背景上的利用潜力。【结果】该六倍体的花粉育性为90.6%—92.7%,自交结实率为3—7粒/角果;菌核病抗性显著高于对照品种(中双9号);处于减数分裂后期I的花粉母细胞中,68.80%(86/125)的染色体分离比为28﹕28。110份春性、冬性和半冬性的白菜型油菜与六倍体杂交,授粉15 d后的胚珠发育正常,并且都能收获成熟种子,平均可交配性为(4.25±3.91)粒/角果。尽管不同基因型之间的可交配性存在显著差异,但是不同生态型的白菜型油菜与六倍体杂交的可交配性无显著差异(半冬性:(4.35±3.77)粒/角果,春性:(4.34±4.51)粒/角果,冬性:(4.01±3.43)粒/角果;=0.44)。六倍体作为母本或者父本与白菜型油菜杂交都能结籽,而且没有显著性差异(六倍体为母本:平均结实率为4.27粒/角果;六倍体为父本:平均结实率为3.95粒/角果;=0.69)。六倍体与白菜型油菜杂交创建的杂种,苗期形态似甘蓝型油菜,但是表型变异丰富;杂种都能自交结籽,平均自交结实率为(7.72±4.45)粒/角果;来自不同生态型的白菜型油菜与六倍体合成的杂种自交结实率无显著性差异(冬性白菜型油菜合成的杂种平均自交结实率:(8.07±3.43)粒/角果,半冬性:(7.88±4.64)粒/角果,春性:(6.41±3.00)粒/角果,=0.95)。经过两年的离体茎秆菌核病抗性鉴定,6份杂种的菌核病发病程度两年均显著低于中双9号(<0.05)。【结论】以六倍体为桥梁能有效地将甘蓝型油菜亲本种的优良性状导入到甘蓝型油菜中。
六倍体;甘蓝型油菜;白菜型油菜;可交配性;菌核病
0 引言
【研究意义】前期研究中,Li等[1]提出了以六倍体AnAnCnCnCoCo为桥梁,与白菜型油菜杂交,创建新型甘蓝型油菜,拓宽甘蓝型油菜遗传背景的策略。本研究将调查以甘蓝型油菜与菌核病抗性甘蓝杂交合成的六倍体为桥梁,与白菜型油菜杂交的可交配性,考察杂种的苗期表型和结实率,并以菌核病抗性为目标性状,评价该策略的应用潜力。这对利用甘蓝和白菜型油菜遗传资源,拓宽和改良甘蓝型油菜遗传背景具有重要意义。【前人研究进展】甘蓝型油菜(,AACC,2n=38)是世界上重要的油料作物之一,是由甘蓝和白菜杂交,染色体自然加倍而成[2-4]。与亲本种相比,甘蓝型油菜的遗传背景相对狭窄[5],利用亲本种遗传资源是拓宽现有甘蓝型油菜遗传背景的重要途径[6-10]。亲本种的利用途径主要有:(1)利用甘蓝与白菜型油菜杂交,人工合成甘蓝型油菜[11-12],该方法需结合胚挽救和染色体加倍等组织培养技术;(2)利用甘蓝型油菜与亲本种杂交,自交或回交,选育新型甘蓝型油菜[4,13-15],该方法从后代中筛选新型甘蓝型油菜所需的周期长,效率低。除此以外,钱伟等[16]发明了一种以甘蓝型油菜与甘蓝杂交创建的六倍体AnAnCnCnCoCo为桥梁,大量快速利用白菜型油菜遗传资源,创建新型甘蓝型油菜,拓宽甘蓝型油菜遗传背景的新策略。在此基础上,Mei等[17]利用六倍体作为桥梁,与甘蓝型油菜不断回交,获得了转移甘蓝菌核病抗性的甘蓝型油菜。【本研究切入点】本研究将采用改良菌核病抗性的六倍体为桥梁,与大量的白菜型油菜杂交,试图导入甘蓝型油菜亲本种优良遗传性状,并利用亲本种广泛的遗传资源,改良现有甘蓝型油菜遗传背景。【拟解决的关键问题】利用菌核病抗性甘蓝与甘蓝型油菜杂交获得的六倍体为桥梁,与白菜型油菜杂交,通过考察可交配性、杂种育性和菌核病抗性来分析该策略对拓宽和改良甘蓝型油菜背景的应用潜力。
1 材料与方法
试验于2013—2015年在西南大学进行。
1.1 试验材料
前期,利用甘蓝型油菜(中双9号)与菌核病抗性甘蓝(C01)杂交,合成了六倍体(AnAnCnCnCoCo)[17]。2013年将74份半冬性、20份春性和16份冬性白菜型油菜(电子附表1)与六倍体杂交,创建新型甘蓝型油菜。成熟期,统计各杂交组合的角果数和种子数,检测可交配性。2014和2015年,杂种F1种植于西南大学油菜工程技术研究中心试验地,用于杂种育性检测和菌核病抗性鉴定。菌核病抗性鉴定采用耐病品种中双9号作为对照。
1.2 试验方法
1.2.1 染色体行为分析 现蕾期,取幼嫩花蕾,于卡诺固定液(酒精﹕醋酸=3﹕1)固定24 h以上,取幼嫩子房用于体细胞染色体数目的观察;取幼嫩花药用于减数分裂时期染色体行为观察。具体方法参照LI等[18]。
1.2.2 菌核病抗性鉴定 本研究采用的菌核病接种和抗性鉴定方法,参照MEI等[19]的离体茎秆接种方法。在90 mm的无菌PDA培养基中,接种菌核病菌丝,于22℃,湿度85%的条件下培养菌丝2 d,待菌丝布满整个培养基时,用直径6 mm的打孔器沿培养皿边缘一圈打孔,获得的菌丝块用于茎秆接种。青荚期,截取植株的茎秆,每个株系取3—5个单株,每个单株取30 cm长的茎秆,两端包裹保鲜膜,每个茎秆接种2个菌丝块于22℃、湿度85%的密闭环境中培养。接种3 d后,统计菌斑长度,并以中双9号作为对照,计算材料的相对感病度=/对照,其中,为待测材料的菌斑长度,对照为‘中双9号’的病斑长度。值越小,表示抗性越强;反之,抗性越弱。
1.3 数据分析
可交配性和育性均采用每角果粒数来衡量,六倍体与白菜型油菜杂交的可交配性=饱满种子数/角果数;新型甘蓝型油菜的育性为自交结实率,结实率=饱满种子总数/取样角果数。数据采用SAS version 8.0(SAS Institute)软件进行方差分析(ANOVA)和相关性分析。
2 结果
2.1 六倍体的特点
本研究采用‘中双9号’与菌核病抗性甘蓝‘C01’()杂交,合成了六倍体AnAnCnCnCoCo(图1-A)。该六倍体的花粉育性是90.6%—92.7%,自交结实率为3—7粒/角果;菌核病抗性高于甘蓝型油菜‘中双9号’(六倍体:接种3 d后的病斑长度为4.6 cm,‘中双9号’接种3 d后的病斑长度是9.6 cm)[17]。六倍体的体细胞染色体数目为56条(图1-B),68.80%(86/125)处于减数分裂后期I的花粉母细胞是28﹕28分离(图1-C)。
2.2 六倍体与白菜型油菜的可交配性
利用六倍体为桥梁,与110份来自3种不同生态型的白菜型油菜杂交,合成杂种。六倍体与白菜型油菜授粉后15 d的胚珠发育正常(图1-D),平均可交配性为(4.25±3.91)粒/角果(最高结实率:17.83粒/角果;最低结实率:0.06粒/角果)。尽管不同基因型的白菜型油菜与六倍体杂交,可交配性存在明显差异,但是来自半冬性、春性和冬性3种生态型的白菜型油菜与六倍体杂交,可交配性没有显著性差异(半冬性:(4.35±3.77)粒/角果;春性:(4.34±4.51)粒/角果;冬性:(4.01±3.43)粒/角果,=0.44,图2)。六倍体与白菜型油菜杂交正、反交之间的可交配性没有显著性差异(六倍体为母本:平均结实率为4.27粒/角果;六倍体为父本:平均结实率为3.95粒/角果,=0.69)。本研究说明了六倍体与白菜型油菜正反交都能结籽。
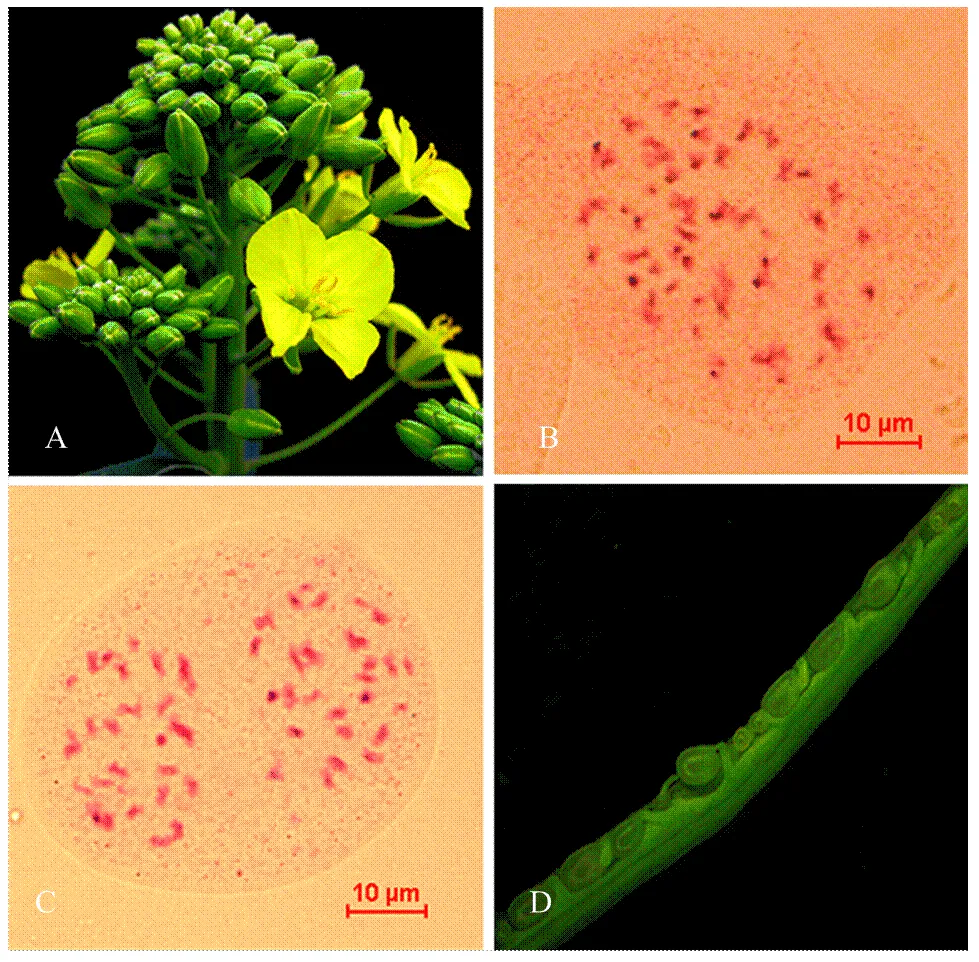
A:六倍体花期形态;B:六倍体体细胞染色体数目(2n=56条);C:六倍体减数分裂后期I染色体分离比(28﹕28);D:六倍体与白菜型油菜授粉15 d后的胚珠发育
2.3 六倍体与白菜型油菜杂种的形态学和育性
选取种子量多的67份(来自46份半冬性、12份春性和9份冬性白菜型油菜)杂种种植于田间,观察杂种的苗期表型和育性。杂种的苗期形态不同于白菜型油菜,与现有的甘蓝型油菜类似,合成的杂种形态学表型变异丰富(图3)。
收获期,统计杂种的自交结实率,结果发现所有杂种都能自交结籽,平均结实率为(7.72±4.45)粒/角果。尽管不同杂种之间育性存在显著性差异,但是根据白菜型油菜的生态型,不同生态型之间没有显著性差异(=0.95,冬性白菜型油菜合成的杂种平均自交结实率:(8.07±3.43)粒/角果;半冬性:(7.88±4.64)粒/角果;春性:(6.41±3.00)粒/角果)(图4)。这说明了不同生态型的白菜型油菜与该六倍体杂交,合成的杂种都能结籽,并且白菜型油菜亲本生态型对合成的杂种结实率没有显著性影响。六倍体与白菜型油菜杂种丰富的表型变异和育性说明了杂种具有拓宽甘蓝型油菜遗传背景的潜力。
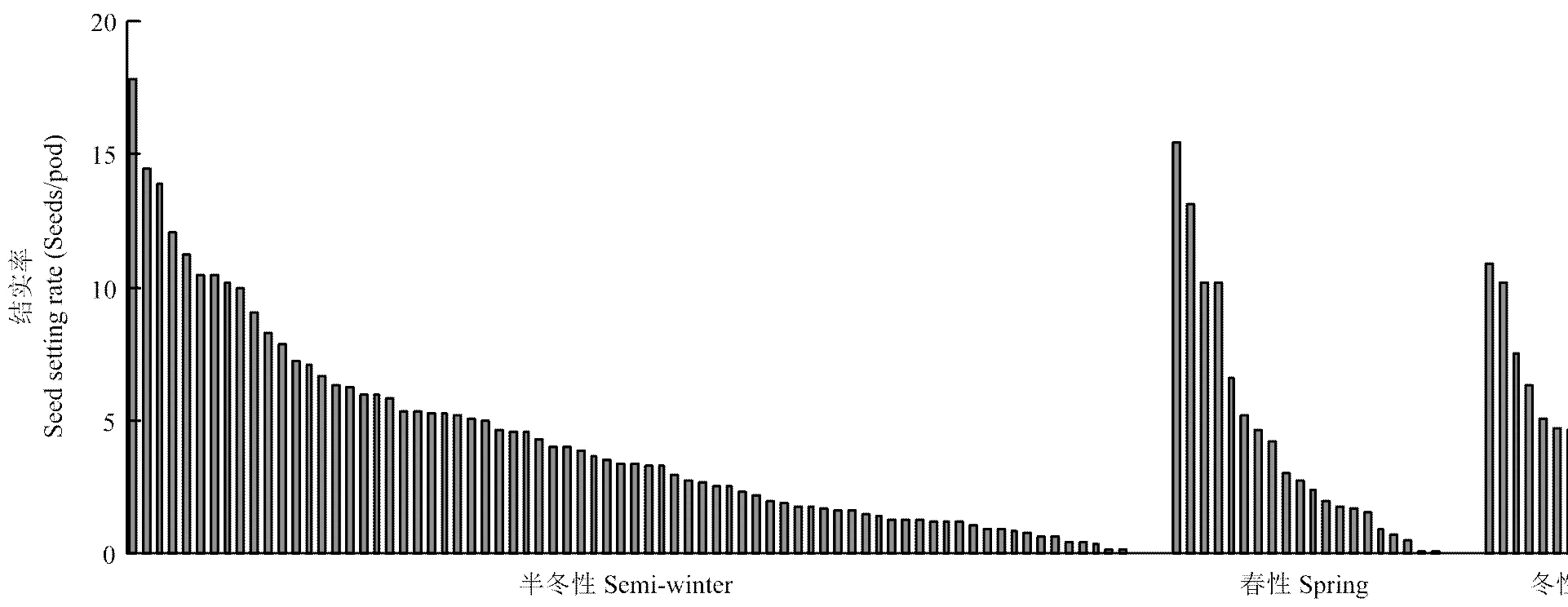
图2 六倍体与110份白菜型油菜杂交可交配性

A:甘蓝型油菜(中双9号),B:六倍体,C—F:六倍体与白菜型油菜杂种,G—I:分别为杂种D—F对应的白菜型油菜亲本

图4 67份白菜型油菜与六倍体杂种的结实率
2.4 六倍体与白菜型油菜杂种的菌核病抗性鉴定
随机选取23份杂种于青荚期截取茎秆进行菌核病抗性鉴定,连续重复两年。结果发现,2014年,这些杂种的平均菌斑长度为(5.77±2.39)cm,相对感病度为0.95±0.42,不同基因型之间存在显著差异(<0.0001)。7个杂种的菌核病抗性显著高于‘中双9号’(平均菌斑长度为:(6.01±1.41)cm),5份杂种的病斑显著大于‘中双9号’。2015年,该23份杂种的平均菌斑长度为:(4.33±1.69)cm,相对感病度为0.73±0.29,不同基因型之间存在显著性差异(<0.0012),11份杂种的病斑显著小于‘中双9号’。其中,6份杂种连续两年的菌核病相对感病度显著低于‘中双9号’(图5,表1)。
将两年的菌核病抗性进行比较,结果发现,尽管不同年度间,杂种的菌核病平均菌斑长度和平均相对感病度存在显著性差异(<0.0001,= 0.0004),但是两年的菌核病发病程度极显著正相关(=0.74,<0.00001)。说明六倍体与白菜型油菜杂种可转移甘蓝的菌核病抗性,表明该途径具有利用甘蓝亲本的遗传变异改良甘蓝型油菜遗传背景的潜力。

图5 六倍体与白菜型油菜茎秆菌核病抗性鉴定

表1 23份六倍体与白菜型油菜杂种的茎秆菌核病抗性鉴定
*代表杂种在=0.05水平上与对照中双9号存在显著性差异
* represents significant difference of hybrids comparing with control Zhongshuang 9 at=0.05
3 讨论
3.1 甘蓝型油菜亲本种的利用途径
甘蓝型油菜是由甘蓝和白菜型油菜杂交,染色体自然加倍而成[2]。甘蓝型油菜起源于7 500年前,但作为栽培种其栽培历史短,经过长期的驯化和进化,甘蓝型油菜的遗传背景比亲本种相对狭窄[3,5,7]。白菜型油菜是一个古老的栽培作物,其农艺特征和DNA结构上具有丰富的遗传变异[20-21]。白菜型油菜与甘蓝型油菜杂交和回交容易,利用该策略创建新型甘蓝型油菜已经在甘蓝型油菜育种中广泛应用[4,6,22]。甘蓝具有丰富的栽培类型和野生资源,以及优良的抗病、抗虫资源[23]。甘蓝型油菜与甘蓝杂交,是利用甘蓝遗传资源的重要手段。将甘蓝型油菜与甘蓝杂交,不断自交,后代也能利用甘蓝遗传资源,合成甘蓝型油菜[14-15]。但是该策略的使用需结合胚挽救获得杂种,杂种F1育性低,需大量的无性系隔离授粉方能获得后代。目前,同时利用白菜型油菜和甘蓝资源的主要途径是人工合成甘蓝型油菜。然而,该途径仍需借助胚挽救等组织培养技术获得杂种,并且后代染色体分离异常,基因组稳定性差[7,24]。钱伟等[16]发明了一种以甘蓝型油菜与甘蓝杂交,染色体加倍获得的六倍体为桥梁,与白菜型油菜杂交,创建新型甘蓝型油菜的方法。该方法不仅利用了甘蓝资源,还导入大量白菜型油菜遗传成分,创建的新型甘蓝型油菜能拓宽现有甘蓝型油菜遗传背景。该策略的提出为本研究的顺利开展提供了思路。本研究提出了以菌核病抗性甘蓝为亲本,与甘蓝型油菜杂交,创建了菌核病抗性改良的六倍体;以该六倍体为桥梁,与大量的白菜型油菜杂交的策略,该策略不仅能利用白菜型油菜丰富资源,还能将甘蓝菌核病抗性导入到油菜中。
3.2 菌核病抗性的改良
菌核病是甘蓝型油菜的主要病害之一,严重危害其生产。尽管育种家们已培育出一些相对耐病的油菜品种(‘中油821’和‘中双’系列品种[25];‘先锋45S51’和‘先锋45S52’[26]),然而,甘蓝型油菜中缺乏菌核病抗性资源。笔者在甘蓝资源中发现了一个野生甘蓝(),对菌核病有突出抗性,其茎秆抗性是甘蓝型油菜耐病品种‘中油821’的20倍[19]。目前,MEI[16]、刘瑶[27]利用该野生甘蓝与甘蓝型油菜杂交,合成了具有菌核病抗性的六倍体,该六倍体与甘蓝型油菜不断回交,选育出的甘蓝型油菜具有改良的菌核病抗性。DING等[28]通过人工合成甘蓝型油菜的途径,将甘蓝亲本的菌核病抗性导入到油菜中。
本研究将菌核病抗性甘蓝‘C01’与甘蓝型油菜‘中双9号’杂交,合成的具有菌核病抗性的六倍体为桥梁,与大量白菜型油菜杂交,合成杂种的菌核病抗性存在显著性差异。相比对照品种‘中双9号’,鉴定的23份杂种中,只有6份杂种菌核病抗性高于对照。产生这种现象的主要原因可能是:一方面,白菜型油菜中广泛缺乏菌核病抗性,造成六倍体与白菜型油菜杂种的菌核病抗性降低;另一方面,甘蓝型油菜与甘蓝杂交合成的六倍体(AnAnCnCnCoCo),由于A、C基因组的高度同源性[29-31],六倍体的染色体减数分裂中产生大量的染色体重排,可能导致配子中菌核病抗性位点的丢失。综上所述,本研究实现了一种利用甘蓝型油菜与菌核病抗性甘蓝杂交,创建抗菌核病的六倍体为桥梁,与白菜型油菜杂交,利用亲本种拓宽和改良甘蓝型油菜遗传背景的新途径。
4 结论
本研究将菌核病抗性甘蓝与甘蓝型油菜杂交合成的六倍体,与110份春性、冬性和半冬性的白菜型油菜正反交均能结籽。杂种苗期形态似甘蓝型油菜,但是表型变异丰富;杂种能自交结籽。经过两年的离体茎秆菌核病抗性鉴定,6份杂种的菌核病发病程度两年均显著低于中双9号。本研究提出的利用菌核病抗性甘蓝与甘蓝型油菜杂交合成的六倍体为桥梁,与大量的白菜型油菜杂交合成的杂种策略,具有利用亲本种遗传资源,拓宽和改良甘蓝型油菜遗传背景的潜力。
References
[1] Li Q, Mei J, Zhang Y, Li J, Ge X, Li Z, Qian W. A large-scale introgression of genomic components ofintoby the bridge of hexaploid derived from hybridization betweenand., 2013, 126(8): 2073-2080.
[2] NAGAHARU U. Genomic analysis in Brassica with special reference to the experimental formation ofand peculiar mode of fertility., 1935, 7: 389-452.
[3] Chalhoub B, Denoeud F, Liu S, Parkin I A P, Tang H, Wang X, Chiquet J, Belcram H, Tong C, Samans B,. Early allopolyploid evolution in the post-Neolithicto oilseed genome., 2014, 345: 950-953.
[4] 刘后利. 油菜遗传育种学. 北京: 中国农业大学出版社, 2000.
Liu H L.. Beijing: Chinese Agriculral Universitatis Press, 2000. (in Chinese)
[5] BECKER H C, ENGQVIST G M, KARLSSON B. Comparison of rapeseed cultivars and resynthesized lines based on allozyme and RFLP markers., 1995, 91: 62-67.
[6] QIAN W, MENG J, LI M, FRAUEN M, SASS O, NOACK J, JUNG C. Introgression of genomic components from Chinesecontributes to widening the genetic diversity in rapeseed (L.), with emphasis on the evolution of Chinese rapeseed., 2006, 113: 49-54.
[7] GIRKE A, SCHIERHOLT A, BECKER H C. Extending the rapeseed gene pool with resynthesizedI: Genetic diversity., 2012, 59: 1441-1447.
[8] GIRKE A, SCHIERHOLT A, BECKER H C. Extending the rapeseed gene pool with resynthesizedII: Heterosis., 2012, 124: 1017-1026.
[9] EICKERMANN M, ULBER B, VIDAL S. Resynthesized lines and cultivars ofL. provide sources of resistance to the cabbage stem weevil ((Mrsh.))., 2011, 101: 287-294.
[10] FU D, QIAN W, ZOU J, MENG J. Genetic dissection of intersubgenomic heterosis incarrying genomic components of., 2012, 184:151-164.
[11] FUJII K, OHMIDO N. Stable progeny production of the amphidiploid resynthesizedcv. Hanakkori, a newly bred vegetable., 2011, 123: 1433-1443.
[12] MALEK M A, ISMAIL M R, RAFII M Y, RAHMAN M. SyntheticL.: development and studies on morphological characters, yield attributes, and yield., 2012, 2012: 416901.
[13] QIAN W, CHEN X, FU D, ZOU J, MENG J. Intersubgenomic heterosis in seed yield potential observed in a new type ofintrogressed with partialgenome., 2005, 110: 1187-1194.
[14] LI Q, ZHOU Q, MEI J, ZHANG Y, LI J, LI Z, GE X, XIONG Z, HUANG Y, QIAN W. Improvement ofvia interspecific hybridization betweenand., 2014, 34: 1955-1963.
[15] RAHMAN M H, BENNETT R A, YANG R C, KEBEDE B, THIAGARAJAH M R. Exploitation of the later flowering speciesL. for the improvement of earliness inL.: an untraditional approach., 2011, 177: 365-374.
[16] 钱伟, 李勤菲, 梅家琴, 付东辉, 李加纳. 一种利用白菜型油菜拓宽甘蓝型油菜遗传变异的方法. 中国, ZL. 201010607185.4. 2013.
Qian W, Li Q F, Mei J Q, Fu D H, Li J N. A strategy of usingto widen genetic variance of: China, Zl. 201010607185.4. 2013. (in Chinese)
[17] MEI J, LIU Y, WEI D, WITTKOP B, DING Y, LI Q, LI J, WAN H, LI Z, GE X, FRAUEN M, SNOWDON R J, QIAN W, FRIEDT W. Transfer of sclerotinia resistance from wild relative ofintousing a hexaploidy step., 2015, 128: 639-644.
[18] LI Z, LIU H L, LUO P. Production and cytogenetics of intergeneric hybrids betweenand.1995, 91:131-136.
[19] MEI J, QIAN L, DISI J O, YANG X, LI Q, LI J, FRAUEN M, CAI D, QIAN W. Identification of resistant sources againstinspecies with emphasis on., 2015, 177: 393-400.
[20] TALEBI R, HAGHNAZARI A, TABATABAEI I. Assessment of genetic variation within international collection ofgenotypes using inter simple sequence repeat DNA markers., 2010, 4: 145-151.
[21] ZHAO J, WANG X, DENG B, LOU P, WU J, SUN R, XU Z, VROMANS J, KOORNNEEF M, BONNEMA G. Genetic relationships withinas inferred from AFLP fingerprints., 2005, 110: 1301-1314.
[22] XIAO Y, CHEN L, ZOU J, TIAN E, XIA W, MENG J. Development of a population for substantial new typediversified at both A/C genomes., 2010, 121: 1141-1150.
[23] MEI J, LI Q, YANG X, QIAN L, LIU L, YIN J, FRAUEN M, LI J, QIAN W. Genomic relationships between wild and cultivatedL. with emphasis on the origination of cultivated crops., 2010, 57: 687-692.
[24] XIONG Z, GAETA R T, PIRES J C. Homoeologous shuffling and chromosome compensation maintain genome balance in resynthesized allopolyploid., 2011, 108: 7908-7913.
[25] 王汉中, 刘贵华, 郑元本, 王新发, 杨庆. 抗菌核病双低油菜新品种中双9号选育及其重要防御酶活性变化规律的研究. 中国农业科学, 2004, 37(1): 23-28.
WANG H Z, LIU G H, ZHENG Y B, WANG X F, YANG Q. Breeding of the Brassica napus cultivar Zhongshuang 9 with high-resistance toand dynamics of its important defense enzyme activity., 2004, 37(1): 23-28. (in Chinese)
[26] FALAK I, MCNABB W, HACAULT K, PATEL J. Field performance ofL. spring canola hybrids with improved resistance to Sclerotinia stem rot. 13th International Rapeseed Congress, Prague, Czech, 2011: 622-626.
[27] 刘瑶, 丁一娟, 汪雷, 万华方, 梅家琴, 钱伟. 甘蓝型油菜与AnAnCnCnCoCo六倍体的可交配性及杂种菌核病抗性. 中国农业科学, 2015, 48(24): 4885-4891.
LIU Y, DING Y J, WANG L, WAN H F, MEI J Q, QIAN W. Crossability betweenwith hexaploid AnAnCnCnCoCo and Sclerotinia resistance in the hybrids., 2015, 48(24): 4885-4891. (in Chinese)
[28] DING Y, MEI J, LI Q, LIU Y, WAN H, WANG L, BECKER H C, QIAN W. Improvement ofresistance inby using., 2013, 60: 1615-1619.
[29] CHEUNG F, TRICK M, DROU N, LIM Y P, PARK J Y, KWON S J, KIM J A, SCOTT R, PIRES J C, PATERSON A H, TOWN C, BANCROFT I. Comparative analysis between homoeologous genome segments ofand its progenitor species reveals extensive sequence-level divergence., 2009, 21: 1912-1928.
[30] LIU S, LIU Y, YANG X, TONG C, EDWARDS D, PARKIN I A, ZHAO M, MA J, YU J, HUANG S,. Thegenome reveals the asymmetrical evolution of polyploid genomes., 2014, 5: 3930.
[31] WANG X, WANG H, WANG J, SUN R, WU J, LIU S, BAI Y, MUN J H, BANCROFT I, CHENG F,. The genome of the mesopolyploid crop species., 2011, 43: 1035-1039.
(责任编辑 赵伶俐)
附表1 110份白菜型油菜类型
Table S1 110 types of

编号Code类型Type来源Origin生态型Ecotype 1PI223501阿富汗 Afghanistan半冬性 Semi-winter 2PI432372孟加拉 Bangladesh半冬性 Semi-winter 3PI392024加拿大 Canada半冬性 Semi-winter 4PI226505伊朗Iran半冬性 Semi-winter 5PI227296伊朗Iran半冬性 Semi-winter 6PI138894伊朗Iran半冬性 Semi-winter 7PI458618新西兰 New Zealand半冬性 Semi-winter 8PI443024挪威 Norway半冬性 Semi-winter 9PI269431巴基斯坦 Pakistan半冬性 Semi-winter 10PI209020波多黎各 Puerto Rico半冬性 Semi-winter 11PI263055俄罗斯 Russian Federation半冬性 Semi-winter 12PI360878瑞典 Sweden半冬性 Semi-winter 131075中国西南大学 Southwest University, China半冬性 Semi-winter 140002-1中国西南大学 Southwest University, China半冬性 Semi-winter 150005-1中国西南大学 Southwest University, China半冬性 Semi-winter 160007-1中国西南大学 Southwest University, China半冬性 Semi-winter 170010-1中国西南大学 Southwest University, China半冬性 Semi-winter 180011-1中国西南大学 Southwest University, China半冬性 Semi-winter 190014-1中国西南大学 Southwest University, China半冬性 Semi-winter 200015-1中国西南大学 Southwest University, China半冬性 Semi-winter 210016-1中国西南大学 Southwest University, China半冬性 Semi-winter 220020-1中国西南大学 Southwest University, China半冬性 Semi-winter 230021-1中国西南大学 Southwest University, China半冬性 Semi-winter 240023-1中国西南大学 Southwest University, China半冬性 Semi-winter 250027-1中国西南大学 Southwest University, China半冬性 Semi-winter 260028-1中国西南大学 Southwest University, China半冬性 Semi-winter 270237-1中国西南大学 Southwest University, China半冬性 Semi-winter 280243-1中国西南大学 Southwest University, China半冬性 Semi-winter 290245-1中国西南大学 Southwest University, China半冬性 Semi-winter 300249-1中国西南大学 Southwest University, China半冬性 Semi-winter 310260-1中国西南大学 Southwest University, China半冬性 Semi-winter 320267-1中国西南大学 Southwest University, China半冬性 Semi-winter 330281-1中国西南大学 Southwest University, China半冬性 Semi-winter 340340-1中国西南大学 Southwest University, China半冬性 Semi-winter 350426-1中国西南大学 Southwest University, China半冬性 Semi-winter 360427-1中国西南大学 Southwest University, China半冬性 Semi-winter 370434-1中国西南大学 Southwest University, China半冬性 Semi-winter 380465-1中国西南大学 Southwest University, China半冬性 Semi-winter 390475-1中国西南大学 Southwest University, China半冬性 Semi-winter 400477-1中国西南大学 Southwest University, China半冬性 Semi-winter 续附表1 Continued table S1 编号Code类型Type来源Origin生态型Ecotype 410489-1中国西南大学 Southwest University, China半冬性 Semi-winter 420595-1中国西南大学 Southwest University, China半冬性 Semi-winter 430768-1中国西南大学 Southwest University, China半冬性 Semi-winter 440770-1中国西南大学 Southwest University, China半冬性 Semi-winter 451773-1中国西南大学 Southwest University, China半冬性 Semi-winter 46毕节油菜 Bijieyoucai中国西南大学 Southwest University, China半冬性 Semi-winter 475K1002中国西南大学 Southwest University, China半冬性 Semi-winter 48黄油1号 Huangyou 1中国西南大学 Southwest University, China半冬性 Semi-winter 496R095中国西南大学 Southwest University, China半冬性 Semi-winter 506Y733中国西南大学 Southwest University, China半冬性 Semi-winter 516Y812中国西南大学 Southwest University, China半冬性 Semi-winter 527B12-1中国西南大学 Southwest University, China半冬性 Semi-winter 537C-126中国西南大学 Southwest University, China半冬性 Semi-winter 549B05-3中国西南大学 Southwest University, China半冬性 Semi-winter 55高棋种 Gaoqizhong中国西南大学 Southwest University, China半冬性 Semi-winter 56黄岩宁波种 Huangyanningbozhong中国西南大学 Southwest University, China半冬性 Semi-winter 57R8021中国西南大学 Southwest University, China半冬性 Semi-winter 58大冶矮脚白油菜 Daye aijiaobaiyoucai中国西南大学 Southwest University, China半冬性 Semi-winter 59云梦半直立油菜 Yunmeng banzhiliyoucai中国西南大学 Southwest University, China半冬性 Semi-winter 60沅江桂林子 Yuanjiang guilinzi中国西南大学 Southwest University, China半冬性 Semi-winter 61治城小花油菜 Zhicheng xiaohuayoucai中国西南大学 Southwest University, China半冬性 Semi-winter 62Ames 30081美国 United States半冬性 Semi-winter 63巢湖油菜 Caohu youcai中国安徽 Anhui, China 半冬性 Semi-winter 64苏州青 Suzhouqing中国江苏 Jiangsu, China半冬性 Semi-winter 65丰城孙渡油菜 Fengcheng sunduyoucai中国江西 Jiangxi, China 半冬性 Semi-winter 66大悟洋白菜 Dawu yangbaicai中国湖北 Hubei, China 半冬性 Semi-winter 67宜昌三区油菜 Yichang sanquyoucai中国湖北 Hubei, China半冬性 Semi-winter 68湘乡油菜 Xiangxiang youcai中国湖南 Hunan, China半冬性 Semi-winter 69武冈油菜白 Wugang youcaibai中国湖南 Hunan, China半冬性 Semi-winter 70长沙姑香 Changsha guxiang中国湖南 Hunan, China半冬性 Semi-winter 71永结油菜 Yongjie youcai中国甘肃 Gansu, China半冬性 Semi-winter 72秦油4号 Qinyou 4中国陕西 Shanxi, China 半冬性 Semi-winter 73宁强仓社矮油菜 Ningqiang cangsheaiyoucai中国陕西 Shanxi, China 半冬性 Semi-winter 74青油13号 Qingyou 13中国青海Qinghai, China半冬性 Semi-winter 75PI254543阿富汗Afghanistan春性 Spring 76PI254542阿富汗Afghanistan春性 Spring 77PI268371阿富汗 Afghanistan春性 Spring 78PI162778阿根廷Argentina春性 Spring 79PI365643加拿大 Canada春性 Spring 80CGN06842德国科隆 Cologne, Germany春性 Spring 81CGN06831德国科隆 Cologne, Germany春性 Spring 续附表1 Continued table S1 编号Code类型Type来源Origin生态型Ecotype 82CGN06832德国科隆 Cologne, Germany春性 Spring 83CGN06838德国科隆 Cologne, Germany春性 Spring 84CGN06839德国科隆 Cologne, Germany春性 Spring 85CGN06840德国科隆 Cologne, Germany春性 Spring 86CGN06841德国科隆 Cologne, Germany春性 Spring 87PI347605印度 India春性 Spring 88PI370740印度 India春性 Spring 89PI267726日本 Japan春性 Spring 90PI343183荷兰 Netherlands春性 Spring 91PI458977美国 United States春性 Spring 92藏油10号 Zangyou 10中国西藏 Xizang, China 春性 Spring 93隆孜县当日镇俗坡村小油菜Supocun xiaoyoucai, Dangri, Longzi中国西藏 Xizang, China春性 Spring 94隆孜县日当镇日当村小油菜Ridangcun xiaoyoucai, Ridang, Longzi中国西藏Xizang, China春性 Spring 95PI254541阿富汗 Afghanistan冬性 Winter 96PI268369阿富汗 Afghanistan冬性 Winter 97PI432376孟加拉 Bangladesh冬性 Winter 98CGN06819德国科隆 Cologne, Germany冬性 Winter 99CGN15216德国科隆 Cologne, Germany冬性 Winter 100ECD04德国 Germany冬性 Winter 101Neib03-2德国 Germany冬性 Winter 102PI458616新西兰 New Zealand冬性 Winter 103PI458931瑞典 Sweden冬性 Winter 104PI458932瑞典 Sweden冬性 Winter 105PI177286土耳其 Turkey冬性 Winter 106PI205283土耳其 Turkey冬性 Winter 107PI176885土耳其 Turkey冬性 Winter 108Ames 30084美国 United States冬性 Winter 109PI633153法国 France冬性 Winter 110PI603024巴基斯坦 Pakistan冬性 Winter
Crossability and Sclerotinia Resistance among Hybrids between Hexaploid (AnAnCnCnCoCo) and
LI Qinfei1,2, CHEN Zhifu2, LIU Yao2, MEI Jiaqin2, QIAN Wei2
(1College of Horticulture and Landscape Architecture, Southwest University, Chongqing 400715;2College of Agronomy and Biotechnology, Southwest University, Chongqing 400715)
【Objective】 Utilization of parental species is helpful to broaden and improve genetic basis of. A strategy of using hexaploid derived from hybrid betweenandas bridge was evaluated to improveby crossing with. 【Method】 Hexaploid derived from interspecific hybrid between(Zhongshuang 9) and(C01, high resistant to) was developed to cross withby evaluating its fertility, meiotic chromosome segregation and resistance to. Hybrids between hexaploid and 110were developed to evaluate crossability, seed setting percentage and resistance to. 【Result】Hexaploid had higher resistance tothan Zhongshuang 9. Its pollen fertility was 90.6%-92.7%, seed setting was 3-7 seeds/pod. The 68.80% pollen mother cells at anaphase I was 28﹕28. The hybrids between 110and hexaploid could be obtained when hexaploid was used as either female or male parent, with an average seed setting rate of (4.25±3.91) seeds/pod (hexaploid used as female parent: 4.27 seeds/pod, male: 3.95 seeds/pod on average,=0.69). Embryos 15 d after pollination grew well when crossing hexaploid with. Although crossability among genotypes ofwith hexaploid was different, no significant difference was detected in crossability among ecotypes of(semi-winter: (4.35±3.77) seeds/pod, spring: (4.34±4.51) seeds/pod, winter: (4.01±3.43) seeds/pod,=0.44). Hybrids owned kinds of morphology at the seedling stage, but were similar with. Seed setting rate of hybrids was (7.72± 4.45) seeds/pod on an average, without significant difference among hybrids derived from spring, winter and semi-winter ecotypes of(seed setting rate of hybrids derived from winter: (8.07±3.43) seeds/pod, semi-winter: (7.88±4.64) seeds/pod, spring: (6.41±3.00) seeds/pod,=0.95). Compared with Zhongshuang 9, 6 hybrids had higher resistance tovia two years identification (<0.05). 【Conclusion】It was concluded that it is an efficient way to improveby transferring elite traits from parental species via hexaploid strategy.
hexaploid;;; crossability;
2016-04-28;接受日期:2016-08-19
中国博士后科学基金(2015M582500)、中央高校基本科研业务费专项(XDJK2016C080)
李勤菲,E-mail:feifei1984998@126.com。通信作者钱伟,E-mail:qianwei666@hotmail.com
猜你喜欢
中国饲料(2022年5期)2022-04-26
麦类作物学报(2020年7期)2020-04-01
西北农业学报(2019年10期)2019-11-20
农村百事通(2019年17期)2019-10-08
农村百事通(2018年16期)2018-09-29
农业科技与装备(2017年9期)2018-03-02
安徽农业科学(2017年2期)2017-07-13
农家科技中旬版(2016年7期)2016-10-20
植物营养与肥料学报(2012年1期)2012-10-26
吉林农业·下半月(2009年6期)2009-07-31
