
根域限制对牡丹花器官碳代谢的影响
2017-09-25 02:54刘晓娟
河南农业大学学报 2017年2期
刘晓娟
(郑州师范学院生命科学学院,河南 郑州 450044)
根域限制对牡丹花器官碳代谢的影响
刘晓娟
(郑州师范学院生命科学学院,河南 郑州 450044)
研究了根域限制(盆栽)对牡丹花器官(花瓣、雄蕊、雌蕊)碳代谢的影响,探索根域限制下牡丹花器官中光合同化物分配和转化酶的变化,为提高盆栽牡丹开花质量、延长花期提供理论和技术支持。结果表明,在根域限制条件下,牡丹花器官出现如下变化:(1)花朵的直径和厚度都显著降低,花器官干质量增加缓慢;(2)花器官之间干质量的分配发生变化,分配到雄蕊的比例增加;(3)花器官总体的蔗糖、己糖含量降低,其中在花瓣和雌蕊中降低程度大,雄蕊中降低程度小;花器官中淀粉含量降低,其中雌蕊中降低最显著。(4)所有花器官中酸性转化酶(Acid Invertase,AI,EC 3.2.1.26)活性显著降低,AI活性的降低使牡丹花瓣、花粉和子房的生长和发育活力都受到影响。因此,根域限制条件下,牡丹花器官库强小,拉动蔗糖输送到花器官的动力不足,蔗糖输送少,蔗糖被利用后,能形成淀粉被存储起来作为储备资源也少,碳水化合物储备资源受限,调动碳水化合物利用的动力不足,以牡丹花径小,开花不良,败育率较高等形式表现出来。
根域限制;牡丹;花器官;碳代谢
随着20世纪60年代容器育苗业的发展,盆栽对植物所产生的影响就越来越受到学者们的研究和关注。盆栽是一种根域限制,根域限制栽培是改善农业资源利用的一种有效的方法,可有效地控制植物地上部分和地下部分以及营养器官和生殖器官同化物的分配[1]。同时,根域限制栽培又是对植物的一种物理胁迫,根域限制下根的生长和发育首先受到抑制,并进行根域信号传递,使整个植株对根域限制胁迫发生反应。根域限制对植物生殖生长的研究表明,根域限制缩短了从播种到开花的时间,使植物开花提前,且根域限制越严重,开花越早[2]。SHI等[3]指出,根域限制越严重,番茄开花期也就越短,但番茄果实成熟延迟。YEH等[4]发现,根域限制使八仙花开花数量减少。SAITO等[1]指出,根域限制使番茄果实减少,产量降低。刘晓娟等[5]研究发现,根域限制显著地降低了牡丹叶片的净光合速率,蔗糖磷酸合成酶和酸性转化酶的活性,“库”限制了“源”,从而影响了碳代谢的中间产物。目前,根域限制的研究主要集中在番茄、葡萄、棉花和西瓜等这几种农作物上,在观赏植物上的研究,国内鲜有报道。盆栽是花卉商品化的重要途径,中国的牡丹资源丰富,栽培面积广,产量大,但是牡丹产业还是以大田种植为主,因为牡丹为落叶灌木,根系为深根系肉质根,在盆栽的条件下,根系生长空间受限,常常造成花蕾小或者不开花。碳水化合物是影响植物生长发育的重要因素,碳代谢决定了植物的源库关系,并控制碳在各种器官中的分区和分布,是植物生长和产量等各方面的基础[6]。因此,本文主要研究根域限制对牡丹花器官(花瓣、雄蕊、雌蕊)碳代谢的影响,探索根域限制下牡丹花器官中光合同化物分配和转化酶的变化,为提高盆栽牡丹开花质量、延长花期提供理论和技术支持。
1 材料与方法
1.1试验材料与处理
以8 a生‘洛阳红’牡丹为研究材料,选择株高、长势一致的分株苗80株,其中花盆栽培40株作为处理,大田栽培40株作为对照。盆栽基质选择与大田栽培土壤一致。花盆高40 cm,直径35 cm。共分10行8列,对照和处理相间排列,株行距为80 cm×100 cm,花盆的1/3置于地面以下防止倒伏。根据牡丹喜干忌湿的特性,在遇干旱时适当浇水。2010—2012年,在每年牡丹开花前(04-18前),开花后(05-10后)和入冬前(12-15前后)3次施肥和松土。分别在牡丹开花的圆桃期(04-05前后),初开期(04-12前后),盛开期(04-18前后)和衰老期(04-25前后)采取牡丹花朵(注:圆桃期为花蕾),盆栽和大田牡丹各取 5 株,每株各取花朵2个。选取花朵分离出花瓣、雄蕊、雌蕊,经液氮处理后,放置于-70 ℃冰箱中,供测试用。
1.2测定项目及方法
花朵直径为花朵正射投影时圆的直径,用数显游标卡尺测量。花朵厚度是从花托到花朵顶部的距离,用数显游标卡尺测量。
选取已经分离出的萼片、花托、花瓣、雄蕊、雌蕊,放入100 ℃烘箱内烘干24 h后,用电子分析天平测量干质量。
取经液氮处理的花瓣、雄蕊、雌蕊样品,分别测定样品中蔗糖、葡萄糖、果糖和淀粉的含量,按照张志良等[7]的方法进行测定。酸性转化酶(Acid Invertase,AI,EC 3.2.1.26)活性的测定参照米国全等[8]的方法。
1.3数据处理与分析
大田试验和室内试验重复3次,所有数据保留小数点后两位数。数据统计分析采用SPSS统计软件,用独立样本t-检验进行显著性分析。
2 结果与分析
2.1根域限制对牡丹花朵发育和器官干质量的影响
从圆桃期到盛开期,是牡丹花器官盛开的过程,从表1可以看出,在盆栽条件下,花朵(花蕾)的直径和厚度都低于对照,且差异显著。
根域限制对牡丹花器官干质量的影响如表2所示。无论根域限制还是对照,花朵总质量的增加主要来自于花瓣。从圆桃期到衰老期,根域限制条件下,牡丹花瓣干质量/花朵总干质量的值分别是51.30%、54.48%、72.09%、58.46%,对照牡丹花瓣干质量/花朵总干质量的值分别是52.81%、63.89%、71.95%、60.30%,所占比例都随着花瓣的开放而增加,在盛开期时达到最大值,花瓣干质量/花朵总干质量的比值,盆栽牡丹与大田牡丹在初开期差异显著。花瓣干质量盆栽牡丹与大田牡丹在圆桃期P<0.05 水平上差异显著,在初开期、盛开期和衰老期在P<0.01 水平上差异显著。
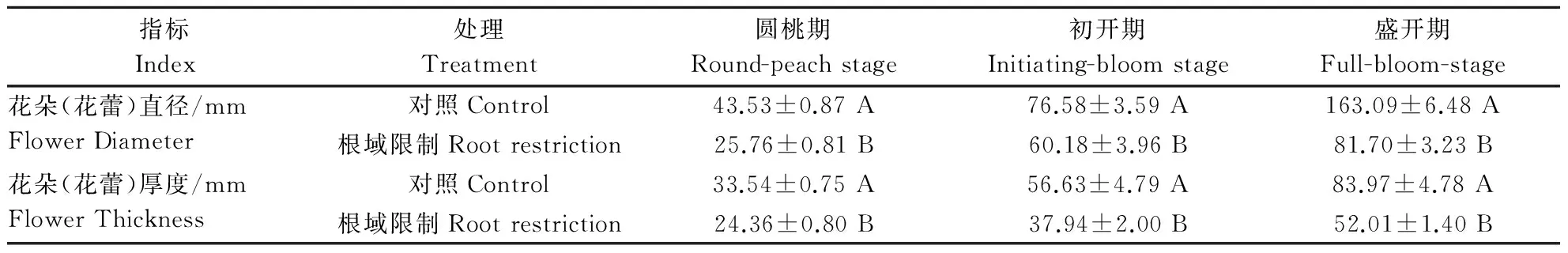
表1 根域限制对牡丹花朵(花蕾)发育的影响 Table 1 Effect of root restriction on flower development in tree peony
注:aa、ab、AB 分别表示差异不显著、在P<0.05,P<0.01水平上差异显著。下同。
Note: aa, ab, AB indicate that difference is not significant, difference is significant atP<0.05,P<0.01levels respectively.The same as below.
由表2可以看出,无论根域限制还是大田种植,雄蕊的干质量都是在初开期达到最大值。根域限制条件下牡丹的雄蕊干质量/花朵总干质量的值是23.05%、20.68%、7.71%、4.29%;对照牡丹的雄蕊干质量/花朵总干质量的值是21.19%、14.67%、7.62%、4.44%,雄蕊干质量/花朵总干质量的值都是在圆桃期达到最大,之后随花朵开放而下降。与圆桃期相比,在初开期,根域限制下牡丹雄蕊干质量增加了50.00%,对照牡丹雄蕊干质量增加了43.08%。在初开期,无论是雄蕊干质量/花朵总干质量的值还是干质量相对上一时期的增加量,根域限制牡丹都显著高于对照。可以看出,在根域限制这种逆境条件下,牡丹植株通过自身调节,优先将营养物质分配给雄蕊生长发育。
雌蕊的干质量随着花的发育逐渐增加,雌蕊干质量/花朵总干质量的比例,根域限制分别是:8.61%、7.57%、6.89%、19.85%;对照分别是:11.38%、8.03%、7.86%、20.61%。在4个时期,根域限制牡丹的雌蕊干质量显著低于对照。
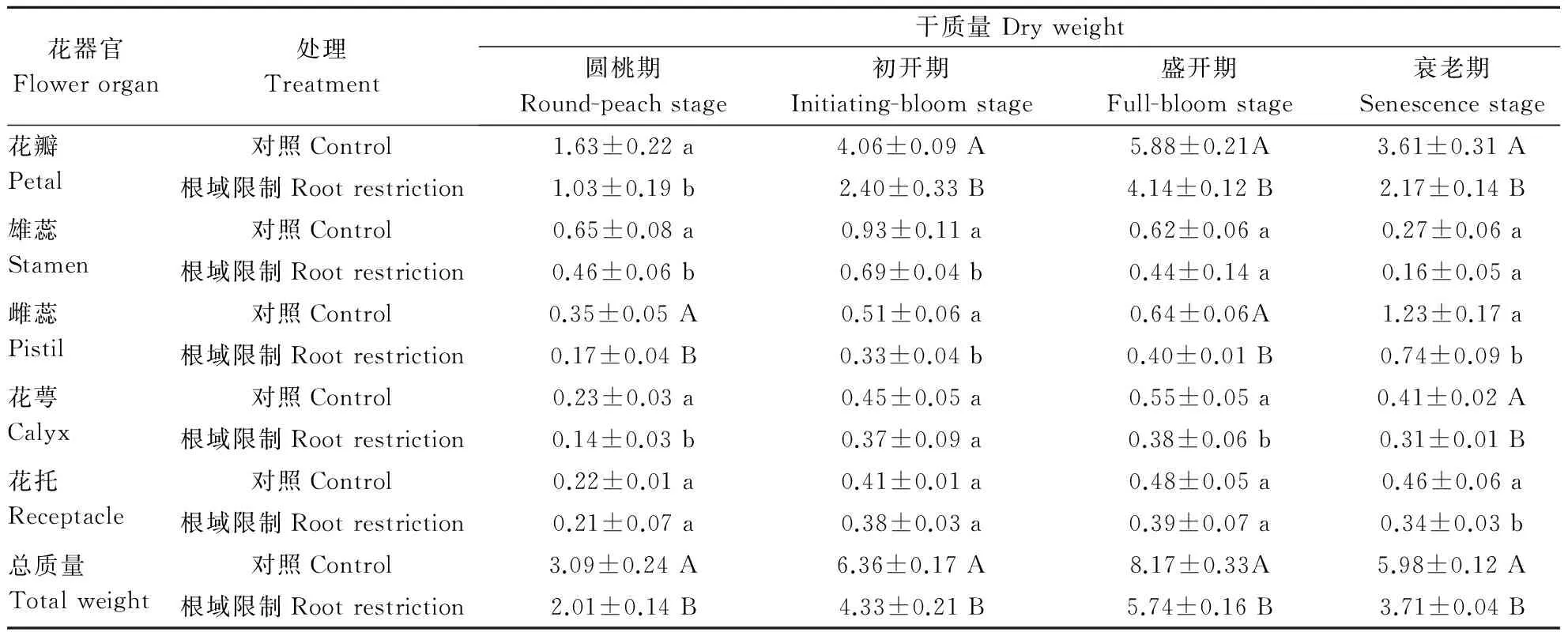
表2 根域限制对牡丹花器官干质量的影响Table 2 Effect of root restriction on dry matter distribution in tree peony flower g
花萼和花托是花器官的支持和辅助部分,二者的生长和发育与整个花朵组织相适应,干质量/花朵总干质量的值基本保持不变。根域限制条件下,牡丹花萼干质量/花朵总干质量的值分别是6.75%、8.48%、6.58%、8.23%;对照牡丹花萼干质量/花朵总干质量的值分别是7.51%、7.04%、6.71%、6.90%,根域限制条件下,牡丹花托干质量/花朵总干质量的值分别是10.29%、8.81%、6.72%、9.17%;对照牡丹花托干质量/花朵总干质量的值分别是7.11%、6.37%、5.85%、7.76%。根域限制下牡丹花朵的总干质量在这4个时期都显著低于对照。
2.2根域限制对牡丹花瓣碳水化合物的影响
从图1中可以看出,在根域限制条件下,在圆桃期、初开期和盛开期,牡丹花瓣中蔗糖含量显著低于对照,分别是对照的68.97%、58.62%、74.11%。在盛开期和衰老期,牡丹花瓣中葡萄糖含量显著低于对照,分别是对照的53.50%和60.00%。在初开期和盛开期,牡丹花瓣中果糖含量显著低于对照,分别是对照的69.23%和74.44%。在圆桃期和初开期,牡丹花瓣中淀粉含量显著低于对照,分别是对照的66.42%和64.38%。
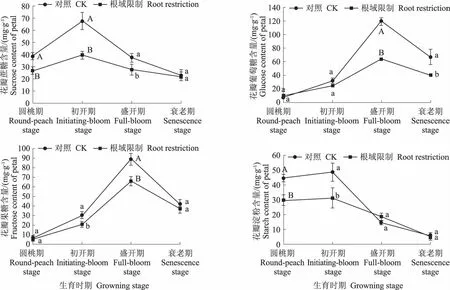
图1 根域限制对牡丹花瓣碳水化合物的影响Fig.1 Effects of root restriction on carbohydrate content in tree peony petal
2.3根域限制对牡丹雄蕊碳水化合物的影响
从图 2 中可以看出,无论处理与对照,牡丹的雄蕊中淀粉的含量最高,蔗糖和淀粉含量在圆桃期最高,之后急剧下降。葡萄糖和果糖含量从圆桃期上升至初开期,之后开始下降,且下降幅度较大。根域限制下,牡丹雄蕊中蔗糖含量在圆桃期显著低于对照,是对照的57.22%,其余时间则与对照无显著差异,葡萄糖含量在圆桃期和初开期显著低于对照,分别是对照的71.56%和52.52%,在盛开期和衰老期则与对照无显著差异。果糖含量在圆桃期和初开期显著低于对照,分别是对照的61.81%和81.15%,在盛开期和衰老期则与对照无显著差异。淀粉含量在圆桃期、初开期和盛开期显著低于对照,分别是对照的47.67%、58.20%和44.44%。
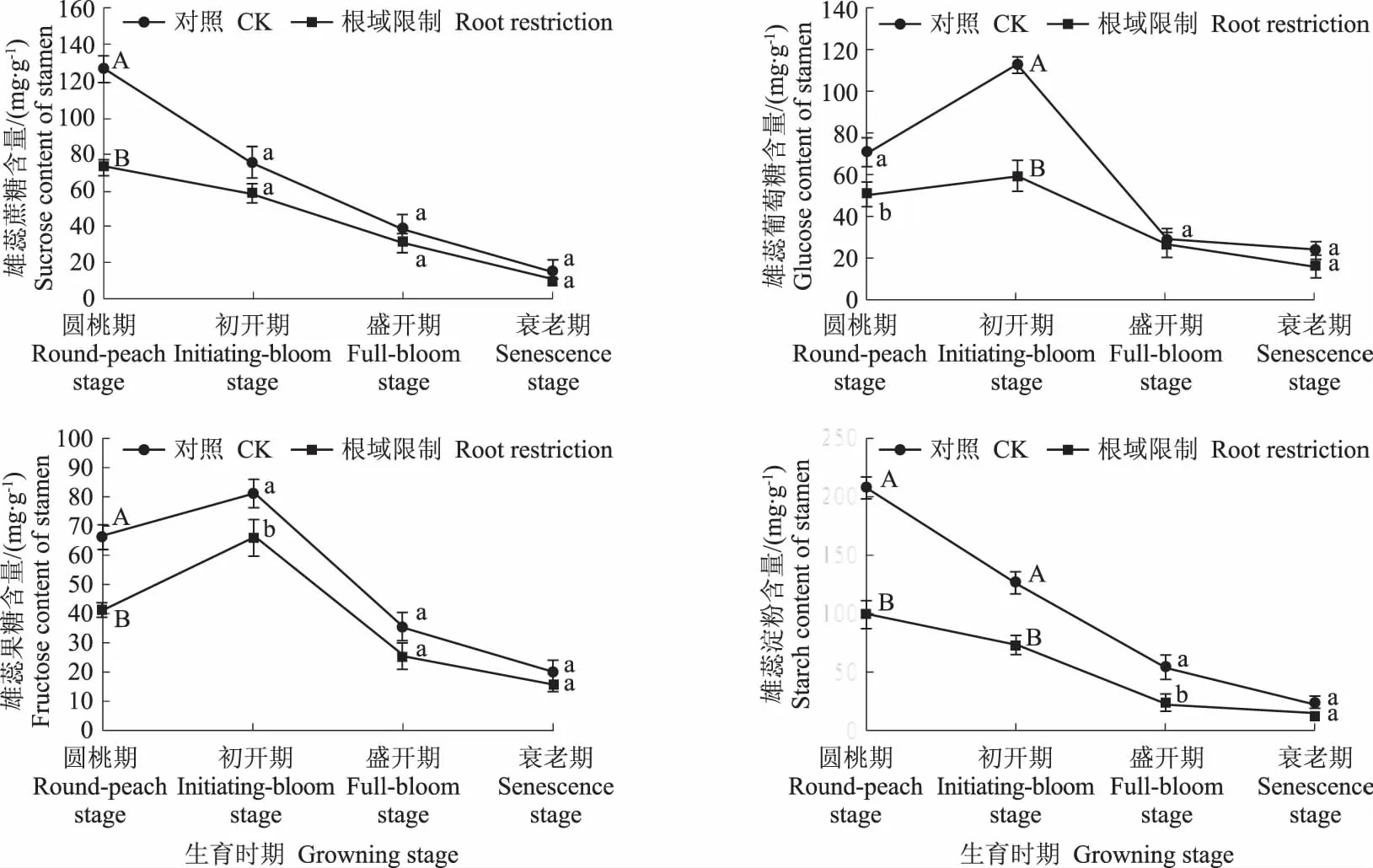
图2 根域限制对牡丹雄蕊碳水化合物的影响Fig.2 Effects of root restriction on carbohydrate content in tree peony stamen
2.4根域限制对牡丹雌蕊碳水化合物的影响
从图3中可以看出,牡丹雌蕊中淀粉含量较高,蔗糖含量变化平稳,从圆桃期缓慢增长到衰老期;牡丹雌蕊中葡萄糖和果糖含量从圆桃期到盛开期持续增加,衰老期有所下降;雌蕊中淀粉含量由圆桃期到初开期呈增加趋势,初开期过后开始降低,但降低幅度不大。根域限制条件下,牡丹雌蕊中在蔗糖含量4个时期显著低于对照,分别是对照的57.88%、63.91%、66.01%、66.82%;雌蕊中葡萄糖含量在4个时期显著低于对照,分别是对照的53.03%、66.10%、44.66%、38.72%;果糖含量在圆桃期和初开期与对照无显著差异,盛开期和衰老期与对照差异显著,分别是对照的79.35%和68.53%;雌蕊中淀粉含量与对照差异最显著,牡丹开花的所有时期,淀粉含量远低于对照,分别是对照的35.06%、32.12%、34.78%、38.97%。
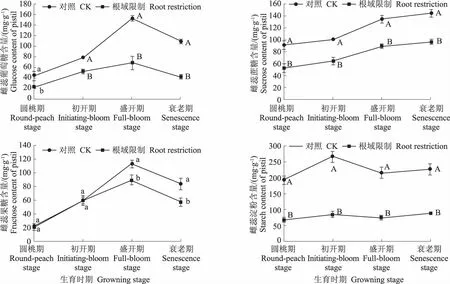
图3 根域限制对牡丹雌蕊碳水化合物的影响Fig.3 Effects of root restriction on carbohydrate content in tree peony pistil
2.5根域限制对牡丹花器官酸性转化酶活性的影响
从图4中可以看出,无论是处理还是对照,牡丹花瓣中AI的变化趋势一致,圆桃期上升到初开期,初开期达到最高之后开始下降,一直持续到衰老期。牡丹雄蕊中AI的变化趋势是,在初开期上升到最高值后开始下降,且下降幅度大;牡丹雌蕊中AI的变化趋势是,从圆桃期开始上升,在盛开期达到最高值后开始下降,表明盛开期牡丹雌蕊已完成受精。在根域限制条件下,在4个时期,牡丹花瓣中AI活性都显著低于对照,分别是对照的53.12%、58.03%、56.39%、62.16%;牡丹雄蕊中AI活性显著低于对照,分别是对照的56.67%、56.06%、62.49%、63.95%。牡丹雌蕊中AI活性低于对照,分别是对照的67.63%、72.89%、69.54%、64.12%,与对照差异显著。
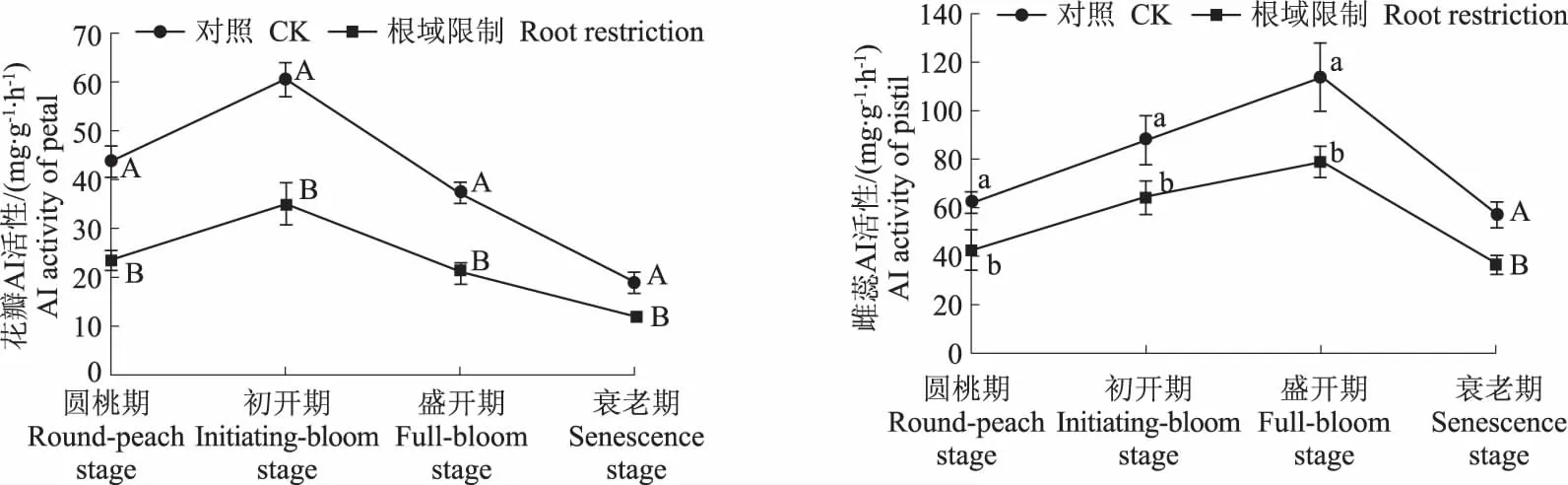
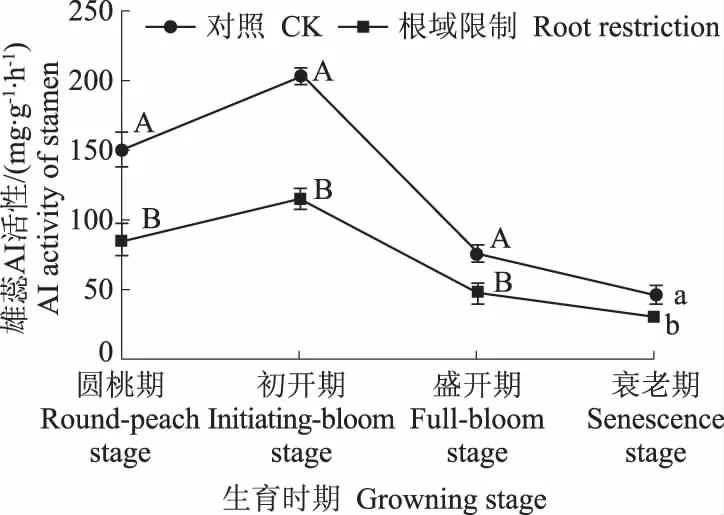
图4 根域限制对牡丹花器官AI活性的影响Fig.4 Effects of root restriction on AI activities in tree peony flower
3 结论与讨论
根域限制条件下,牡丹花朵(花蕾)的直径和厚度都显著降低,牡丹根部为肉质根,是光合产物最主要的存储器官,牡丹开花期间主要的能量来自于根部[9],根域限制条件下,牡丹花朵(花蕾)的直径和厚度都显著降低,这与牡丹叶片光合速率降低[5],根部存储功能下降有关。牡丹单个花器官干质量的增加主要来自于花瓣[10]。干质量是从源器官传送到库器官的同化物,干物质在不同器官之间分配的多少主要取决于库强的大小,而分配的快慢则为器官潜在的生长速率,即生长速率越高,干物质分配就越多。根域限制条件下,牡丹的花瓣、雌蕊、雄蕊和整个花器官的干质量都低于对照,说明根域限制使牡丹花器官的库强降低,生长速率也降低。根域限制牡丹在初开期花瓣所占干质量比例显著低于对照,从圆桃期到初开期花瓣干质量的增加速度也低于对照。这说明根域限制抑制了牡丹花瓣干质量的增加和干质量增加的速度。
逆境条件下,植物形成抗逆机制保护生殖发育,保证最大产量的形成[6]。从圆桃期到盛开期,雄蕊干质量/总干质量的比值高于对照,雌蕊干质量/总干质量的比值低于对照,表明到初开期,根域限制下牡丹植株优先供应雄蕊的生长,营养物质分配到雄蕊的比例增加。这说明牡丹在根域限制的条件下,优先供应生殖器官的生长,尤其是花药的生长,这样能为后期的花粉生长提供保证。
一般情况下,蔗糖、葡萄糖和果糖是植物花瓣中主要的可溶性糖[11]。蔗糖是从叶片转移到花器官的糖的主要形式。从圆桃期到盛开期,根域限制牡丹花瓣中蔗糖含量显著低于对照,雌蕊中蔗糖含量在4个时期显著低于对照,雄蕊中在圆桃期,蔗糖含量低于对照,在初开期、盛开期和衰老期则与对照无显著差异。这说明根域限制使牡丹花器官总体的蔗糖含量降低。但是降低的程度不一样,在雄蕊中降低的程度要低于在花瓣和雌蕊中降低的程度。从而证实根域限制下,不同花器官对碳水化合物的吸收存在着竞争,牡丹花器官优先将碳水化合物供应给雄蕊。
植物的花器官是一个强大的库器官,尤其是开花前,库强最大,需要吸收大量的光合同化物,蔗糖进入花瓣后,必须分解成葡萄糖和果糖才能被花瓣利用[12]。在盛开期之前,牡丹花瓣中己糖含量显著低于对照,这一方面与花瓣中蔗糖的含量较低有关,另一方面也与AI活性较低,分解蔗糖的速率较低有关。雌蕊中葡萄糖含量在4个时期显著低于对照。较之花瓣和雌蕊,雄蕊中己糖含量与对照的差异就相对小些,这可能与雄蕊对碳水化合物的竞争能力较强,蔗糖含量相对较多有关。
淀粉是植物碳水化合物存储的主要形式,当库强需求增加时,淀粉水解为植物提供能量[13]。在植物开花中,糖含量的增加是淀粉含量降低的7—8倍,表明花器官对糖的吸收促进了淀粉的分解。在开花过程中,尤其是花瓣展开的过程中,淀粉分解是可溶性糖增加的一个主要来源。根域限制下,牡丹花瓣中淀粉含量在圆桃期和初开期,显著低于对照,雄蕊中淀粉含量在圆桃期、初开期和盛开期显著低于对照,雌蕊中淀粉含量在4个时期都显著低于对照。雌蕊中的淀粉含量,尤其是子房中的淀粉含量,对后期种子的繁殖和发育有重要的作用,子房对能源的不足非常敏感,非还原性糖和淀粉的减少影响了子房的发育[14]。MCLAUGHLIN等[15]研究指出,在干旱情况下,子房中的葡萄糖被耗尽,当另外使用蔗糖时,可以阻止不育。KUDO等[16]研究发现海拔较高的山地生长的黄芪(Astragalus-alpinus)胚珠受精后,种子败育率提高,认为是营养物质限制引起的。由此可以看出,雌蕊中可溶性糖和淀粉的不足,是根域限制牡丹败育率较高的原因。
任何植物的生长和发育都是碳水化合物的代谢和积累过程[13]。库强对光合产物向库器官的分配具有极其重要的作用,源中蔗糖的输出速率和输出方向由库强控制。库强较大时,进入库细胞的蔗糖或被合成贮藏物质(主要是淀粉),或分解用于库细胞的生长,从而使库细胞处于低浓度的蔗糖状态,保持了源库两端压力势差,从而使源端制造的光合产物源源不断地运入库,也维持了源强[17]。作为一种非光合器官,植物花器官发育中源库之间碳水化合物的转移与转化酶活性密切相关[18]。转化酶在很大程度上决定了花器官库强的大小,从圆桃期到衰老期,根域限制牡丹花瓣、雄蕊、雌蕊中AI活性都显著低于对照,AI活性的降低使牡丹花瓣、花粉和子房的生长和发育活力都受到影响。综合根域限制下牡丹花器官中蔗糖、己糖、淀粉和AI的变化,可以看出根域限制牡丹花器官生长和发育较对照不良的根本原因是库强小,AI活性较低,拉动蔗糖输送到花器官的动力不足,蔗糖输送的少,蔗糖被利用后,能形成淀粉被存储起来作为储备资源也少,碳水化合物储备资源受限,调动碳水化合物利用的动力(AI)不足,以牡丹花径小、开花不良、败育率较高等形式表现出来。
[1] SAITO T, FUKUDA N, LIKUBO T, et al. Effects of root-restriction and salinity on the fruit yield and quality of processing tomato[J] . The Japanese Society for Horticultural Science, 2008, 77(2): 165-172.
[2] RUBEN R, GUILLAUME L, JOSE R D. Environmental control of root system biology[J]. Plant Biology, 2016, 67(67):619-642.
[3] SHI K, DING X, DONG D, et al. Root restriction-induced limitation to photosynthesis in tomato (LycopersiconesculentumMill.) leaves [J]. Scientia Horticulturae, 2008,117(3): 197-202.
[4] YEH D M, CHIANG H H. Growth and flower initiation in hydrangea as affected by root restriction and defoliation[J]. Scientia Horticulturae, 2001,91(1/2):123-132
[5] 刘晓娟,朱丽娟,杨秋生,等. 根域限制对牡丹叶片碳代谢的影响[J]. 林业科学,2012,48(12): 53-57.
[6] LEBON G, WOJNAROWIEZ G, HOLZAPFEL B, et al. Sugars and flowering in the grapevine (VitisviniferaL.)[J]. Journal of Experimental Botany,2008, 59(10): 2565-2578.
[7] 张志良,瞿伟菁,李小方.植物生理学实验指导[M].郑州:高等教育出版社. 2009,
[8] 米国全,刘丽英,金宝燕,等. 弱光对不同生态型黄瓜幼苗光合速率及蔗糖代谢相关酶活性的影响[J]. 华北农学报,2011,26(1): 146-150.
[9] 高志民,王雁,李振坚,等. 牡丹开花前后营养变化分析研究[J]. 林业科学研究,2007,20(3): 390-393.
[10] 史国安,郭香凤,张国海,等. 牡丹开花和衰老期间花瓣糖代谢的研究[J]. 园艺学报,2009,36(8): 1184-1190.
[11] YAP Y, LOH C, ONG B. Regulation of flower development in Dendrobium crumenatum by changes in carbohydrate contents, water status and cell wall metabolism[J]. Scientia Horticulturae, 2008, 119(1): 59-66.
[12] LEBON G, WOJNAROWIEZ G, HOLZAPFEL B, et al. Sugars and flowering in the grapevine (VitisviniferaL.) [J]. Journal of Experimental Botany, 2008, 59(10): 2565-2578.
[13] FERNIE A R, WILLMITZER L, TRETHEWEY R N. Sucrose to starch: a transition in molecular plant physiology[J] . Trends in Plant Science, 2002, 7(1): 35-41.
[14] RODRIGO J, HORMAZA J I, HERRERO M. Ovary starch reserves and flower development in apricot (Prunusarmeniaca) [J]. Physiologia Plantarum, 2000, 108(1): 35-41.
[15] MCLAUGHLIN J E, BOYER J S. Sugar-responsive gene expression, invertase activity, and senescence in aborting maize ovaries at low water potentials[J]. Annals of Botany, 2004, 94(5): 675-689.
[16] KUDO G, MOLAU U. Variations in reproductive traits at inflorescence and flower levels of anarcticlegume,AstragalusalpinusL. Comparisons between a subalpine and an alpine population[J]. Plant Species Biology, 1999, 14(3): 181-191.
[17] LIM J D, CHO J, PARK Y, et al. Sucrose transport from source to sink seeds in rice[J]. Physiologia Plantarum, 2006, 126(4): 572-584.
[18] ESHGHI S, TAFAZOLI E, DOKHANI S, et al. Changes in carbohydrate contents in shoot tips, leaves and roots of strawberry (Fragaria×ananassaDuch.) during flower-bud differentiation[J] . Scientia Horticulturae, 2007, 113(3): 255-260.
(责任编辑:李 莹)
Effectsofrootrestrictiononcarbohydratemetabolismintreepeonyfloralorgans
LIU Xiaojuan
(College of Life Sciences,Zhengzhou Normal College, Zhengzhou 450044 , China)
This paper studied the carbon metabolism of tree peony’s flower organ (petal, stamen, pistil) under root restriction,to explore the photoassimilate partitioning and invertase activity of tree peony under root restriction, aiming to improve flower quality and to extend flowering period. The results showed that, under root restriction, tree peony’s flower organ showed the following changes:1.The diameter and thickness of tree peony flower were significantly reduced and dry matter increase rate was much lower;2.Dry matter distribution of flower organ changed, which distributed relatively more to stamens; 3.Sucrose content and hexose content of tree peony flower organ were decreased, which reduced much more in petals and pistils than in stamens. The starch content in flower organs were lower than control generally, which reduced much more in stamens; 4. Acid invertase (AI) activities of tree peony flower organ were significantly lower than that of the control, which influenced growth and development of petal, pollen and ovary. It can be seen that the sink strength of flower organ was not strong enough and have insufficient power to transport sucrose. Sucrose was transported little. When sucrose was used, there was less sucrose to form starch as a reserve resource. There were not enough carbohydrate and lack of motivation to mobilize carbohydrate to utilize, so the flower diameter was small, with bad flowering and higher abortion rate in tree peony when it was growing under root restriction.
root restriction; tree peony; floral organ; carbohydrate metabolism
S685.11
:A
2016-03-24
国家自然科学基金项目(31140057);河南省高等学校重点科研项目计划(17A220002);郑州市科技攻关项目(153PKJGG145)
刘晓娟(1982-),女,河南郑州人,讲师,博士,主要从事风景园林方面的研究。
1000-2340(2017)02-0170-07
猜你喜欢
经济林研究(2022年2期)2022-07-20
课外生活·趣知识(2022年6期)2022-07-14
现代农业科技(2022年4期)2022-03-08
中国农学通报(2022年4期)2022-03-02
中国甜菜糖业(2020年3期)2020-12-08
花生学报(2019年2期)2019-10-22
浙江工业大学学报(2017年5期)2018-01-22
生物学教学(2017年6期)2017-02-18
中国糖料(2016年1期)2016-12-01
红蜻蜓·低年级(2016年9期)2016-05-14
