
不同密度下羊草种群的无性繁殖对策
2017-09-13 05:23:50金晓明官庆新马梁硕张忠哲
草地学报 2017年4期
金晓明, 兰 雪, 官庆新, 马梁硕, 张 骁, 张 浩, 张忠哲
(呼伦贝尔学院生命与环境科学学院, 内蒙古 呼伦贝尔021008)
种群密度在一定程度上反映了竞争强度以及环境条件,因而种群密度被作为研究种群生长和繁殖的主要生态因子[1]。植物的繁殖对策是指植物在其生活史中,通过最佳的资源分配格局,以其特有的繁殖属性去适应环境,提高植物适合度的自组织过程。其适应的对象是生存环境,最终目标是物种的持续生存和繁衍[2]。无性系植物指具有无性系性的植物,即生物在自然状况下自发地产生遗传结构相同的后代个体的生物学习性[3],包括无性生长和无性繁殖两大类[4]。呼伦贝尔草原处于我国北部,是畜牧业生产的重要基地,羊草(Leymuschinensis)是呼伦贝尔天然草原的主要优势种,为无性系植物,是既能进行营养繁殖, 又能进行有性生殖的多年生禾本科牧草[5]。目前,国内外许多学者对种群的密度调节[6-9]及羊草种群生态学方面开展了广泛的研究[5,10-18],如羊草觅养生长格局、年龄结构及无性繁殖对策、生物量动态、种群数量特征等方面的研究。但有关栽培密度对羊草种群繁殖适应的研究还很缺乏[19-20]。本文以不同栽培密度的羊草种群为研究对象,旨在探讨羊草种群在密度制约条件下的生长及繁殖动态,以期增进对羊草在空间环境变化下的构件调节适应机制,为植物繁殖生态学的研究提供理论依据。
1 材料与方法
1.1 研究区概况
试验区位于内蒙古呼伦贝尔市鄂温克族自治旗的呼伦贝尔学院牧草种质资源研发基地内。E119°43′87″,N49°12′84″,海拔577 m。气候属中温带大陆性季风气候,冬季漫长寒冷,夏季温和短促。年平均降水量为350 mm左右, 其中70%集中于夏秋季节;蒸发量1 300 mm,为降水量的4~6倍,年平均日照时数约为2 800~3 100 h,全年无霜期100~120 d[21]。试验地土壤为暗栗钙土。
1.2 试验材料与方法
于2013年7月初,通过种子播种方式,建立羊草(Leymuschinensis)人工草地。播种时设置10个羊草播种量梯度(即10个播种密度),播种量(密度梯度)如表1所示。每个梯度3个重复,分别播种3个小区,共30个小区,随机排列,每个小区面积15 m2,小区之间间隔0.5 m。样地播种小区布局如图1所示。播种后进行灌溉和除杂草等田间管理。

表1 羊草的播种量Table 1 Seeding rates of Leymus chinensis

图1 样地布局Fig.1 The layout of sample area
于2015年8月10日对人工草地的羊草按不同密度梯度小区进行随机取样,每个梯度3次重复,样方面积为50 cm×50 cm,地下深度为30 cm,将羊草的地上和地下部分完整挖掘出来,编号,装入塑料袋带回实验室。
年龄结构的划分方法采用杨允菲[14]对羊草分株的年龄结构鉴定方法。分别统计1龄分株、2龄分株和3龄分株的数量,然后在80℃下烘干至衡重后称量其干重(生物量)。
按羊草无性系的构件,即茎、叶、穗、根茎和须根进行分置,分别测定其干重,同时测定根茎累计长度。
由根茎顶端形成的芽称为根茎顶端芽,由根茎节间形成的芽称为根茎节间芽,由分株分蘖节形成的芽称为分蘖节芽。对根茎顶端芽、根茎节间芽、分蘖节芽的数量进行计数,即芽数量。
1.3 数据分析
地上生物量=茎生物量+叶生物量+穗生物量
地下生物量=根茎生物量+须根生物量
茎/叶=茎生物量/叶生物量
根茎/须根=根茎生物量/须根生物量
地上/地下=地上生物量/地下生物量
根茎累计长度为样方中所有根茎的长度之和。
运用SPSS 19.0软件进行数据处理,对不同处理之间进行单因子方差分析(One—way ANOVA),对于具有显著性差异的不同处理之间进一步做Duncan多重比较。
2 结果与分析
2.1 不同密度下羊草种群分株的年龄结构
不同密度下羊草种群分株数量和分株生物量的年龄结构如图1所示。由图1可知,各龄级分株数量的年龄谱中,1龄级分株占绝对优势,占总分株数量的85.1%(数据范围66.7%~93.9%),2龄级分株数量占总分株数量的13.5%(数据范围6.1%~24.2%),3龄级分株数量占总分株数量的1.4%(数据范围0~9.1%)。由此表明,不同密度下,羊草种群分株数量均为增长型年龄结构。对不同密度下1龄级分株数量进行比较,结果表明,T1,T2和T3显著高于其他处理,而T9最低。

图2 不同密度下羊草分株数量的年龄结构Fig.2 Age structures of the number of ramets of L.chinensis under different densities
在年龄级组成上,CK,T1,T2包含3个年龄级(1龄、1龄和3龄),T3,T4,T5,T6,T7,T8,T9包含2个年龄级(1龄和2龄)。表明,随着密度的增加,羊草分株的年龄级组成数量呈减少趋势。
由图2可知,各龄级分株生物量的年龄谱中,1龄级分株占绝对优势,占总分株生物量的95.8%(数据范围89.3%~98.1%),2龄级分株生物量占总分株生物量的3.9%(数据范围1.9%~9.6%),3龄级分株生物量占总分株生物量的0.3%(数据范围0~2.1%)。由此表明,在不同密度羊草分株生物量上,1龄级分株占绝对优势,而2龄级和3龄级的比例则很小。因此,不同密度下,羊草种群分株生物量呈增长型年龄结构。对不同密度下1龄级分株生物量进行比较,结果表明CK、T1和T2显著高于其他处理,而T6、T8和T9最低。
随着密度的增加,羊草总分株生物量呈逐渐降低趋势。其中,CK、T1和T2之间均无显著差异,但显著高于T3~T9。羊草总分株生物量和1龄级分株生物量,在T3~T9呈现逐渐降低趋势,而羊草2龄级分株生物量,在T3~T9之间基本一致。3龄级分株生物量上,CK、T1和T2的分株生物量之间均无显著差异。

图3 不同密度下羊草分株生物量的年龄结构Fig.3 Age structures of the biomass of ramets of L.chinensis under different densities
由羊草分株数量和生物量的年龄结构可知,随着密度的增加,羊草分株的年龄级组成由CK~T2的3个年龄级变成T3~T9的2个年龄级,即密度越高,种群的年龄结构越不稳定。
2.2 不同密度下羊草种群的构件分配特征
不同密度下羊草种群的构件分配特征如表2所示。随着密度的增加,羊草的根茎累计长度、地下生物量和分株数量均呈先增加后降低的趋势。根茎累计长度在T2和T3水平上显著高于其他密度水平。地下生物量和分株生物量在T1和T2水平上显著高于其他密度水平。而地上生物量则随着密度的增大呈逐渐降低趋势。由此表明,随着栽培密度的增加,羊草在种间竞争中采取优先延长根茎长度,然后再增加分株数量和地下生物量的对策。当密度达到T4水平时,根茎累计长度、分株数量和地下生物量开始降低。当密度提高到T9水平时,羊草地上和地下构件的生长受到严重制约,尤其是根系的生长变得非常缓慢。而密度越低越有利于地上生物量的积累。

表2 不同密度下羊草种群的繁殖分配特征Table 2 Characteristics of reproductive allocation of L.chinensis under different densities
不同栽培密度下羊草种群的构件生物量比值如表3所示。在任何密度水平下羊草的根茎/须根都是稳定不变的。在CK~T3时,茎/叶呈先增加后降低的趋势;在T4~T8时,随着密度的增加,茎/叶呈降低趋势;在T9时,则呈升高趋势。
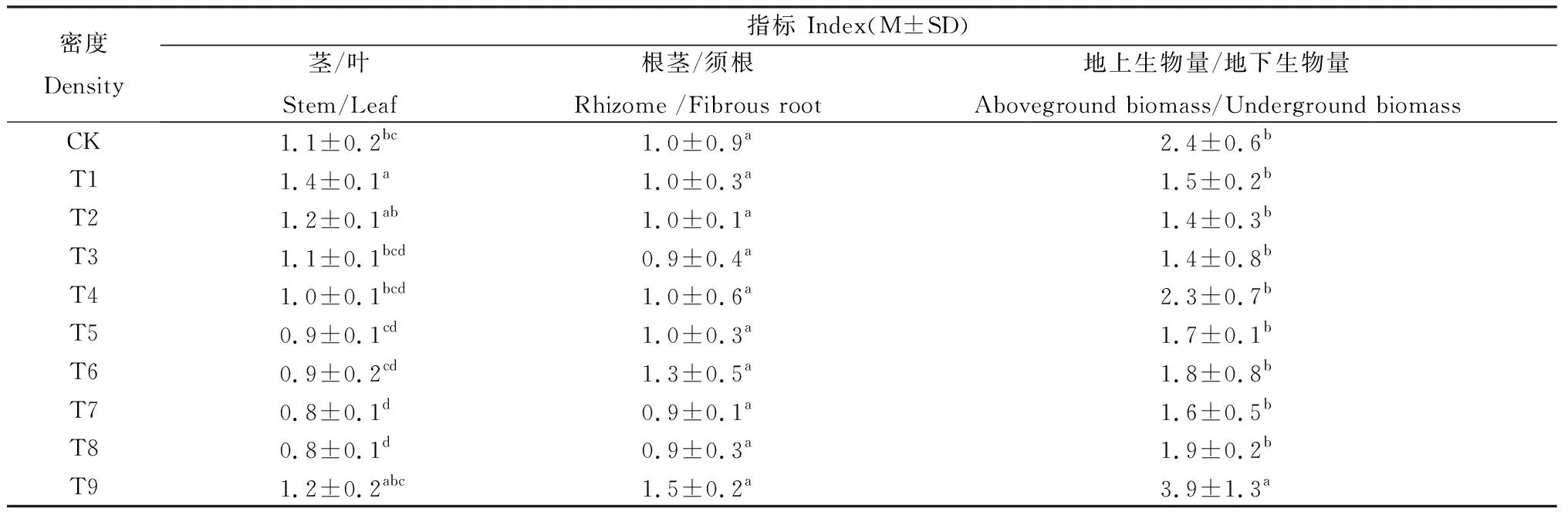
表3 不同密度下羊草种群的构件生物量比值Table 3 The ratio of modules’ biomass of L.chinensis under different densities
T9地上生物量/地下生物量显著高于T1~T8,而T1~T8之间均无显著差异。表明栽培密度足够大时,羊草通过提高地上生物量/地下生物量的构件分配调节策略来适应空间环境的变化。另外,随着密度的增加,地上/地下呈波动变化趋势,即在T1~T3水平下呈现下降现象。由此表明,当羊草根茎快速生长时,其地上/地下呈下降趋势,而茎/叶呈增加趋势。
2.3 羊草潜在种群与现实种群的相关性
羊草潜在种群与现实种群相关指标的相关性分析如表4所示。

表4 羊草潜在种群与现实种群之间的相关系数Table 4 Correlation coefficient between potential population and real population of L.chinensis
总芽数代表潜在种群的数量。由表4可知,总芽数与地上生物量、地下生物量和根茎累计长度之间的相关系数分别为0.654,0.658和0.436,即总芽数与地上生物量和地下生物量的相关程度高于根茎累计长度。
潜在种群的3个指标中,根茎顶端芽和根茎节间芽数量与现实种群的相关系数较高,分别为0.573和0.550,而分蘖节芽则较低,为0.217。表明,羊草将自身积累的能量主要用于具有空间侵占能力的根茎顶端芽和根茎节间芽上,即人工种群建立初期,羊草优先采取空间扩展的策略。
现实种群的3个指标中,地上生物量与潜在种群的相关系数最高,平均达0.525,其次是地下生物量与潜在种群的相关系数,平均为0.500,而根茎累计长度与潜在种群的相关系数最低,为0.315。表明人工种群建立初期,羊草地上生物量、地下生物量和根茎累计长度的物质积累均有利于潜在种群的发育,按其对潜在种群数量增长的贡献率进行排序为:地上生物量>地下生物量>根茎累计长度。其中地上生物量与根茎顶端芽的相关系数最高,为0.717,即羊草地上生物量是判断其潜在扩展空间能力的主要因素。
3 讨论与结论
在植物整个生活史进化过程中,无性系的分株密度起着重要作用[22]。密度增加导致无性系植物的种内竞争,从而使无性生长的资源分配发生改变。无性系植物的种内竞争包括两个水平,即基株之间的竞争和分株之间的竞争[23]。其中,基株水平上的竞争对进化是有意义的[24]。本文的研究对象是不同栽培密度下生长了3年的羊草种群,因此既包括基株水平上的竞争,又包括分株之间的竞争。随着密度的增加,总分株数量、地上和地下生物量及根茎累计长度均呈降低趋势,即随着播种密度的增加,种内竞争加剧,羊草为了保持种群的生存、生长和繁殖,在播种后的前2年内就淘汰了一部分单株。而较低的播种密度则有利于羊草种群的繁殖,无论在种群数量还是生物量上均高于高播种密度。
无性系植物的营养生长使得个体在空间和资源利用、逃避环境风险等方面具有明显的优势[25],植物在生活史各个阶段都要承受由环境、竞争、捕食等引起的各种压力,植物为了使种群得到延续,必须对繁殖过程进行调节[26]。Rautiainen 等对人工环境下鹅绒委陵菜(Potentillaanserine) 的一个亚种(P.anserine‘egedii’)的研究发现,无性系植物对资源分配的最适转换策略,当无性系植物在新的未被占领的斑块内时,能依靠无性生长迅速拓展空间,从而获得更多支持,而当无性系密度达到一定程度,其种内竞争限制了无性系向外传播时,无性系基株在维持其生存的前提下,有效发挥种子长距离传播能力[27]。本研究中,栽培密度对羊草种群造成一定空间环境上的竞争压力。在低密度(CK~T3)时,随着密度的增加,羊草在种间竞争中采取优先延长根茎长度,然后再增加分株数量和地下生物量的繁殖适应对策。在中等密度(T4~T8)时,随着密度的增加,羊草采取降低根茎累计长度、分株数量和地下生物量的繁殖对策。在高密度(T9)时,羊草的地上和地下构件的生长受到严重制约,羊草采取优先生长地上茎和叶,而减缓地下根系生长的策略。但在本研究中,羊草的有性繁殖极少,即使高密度下,也没有增加羊草种群的有性繁殖。这是因为羊草的有性繁殖特征明显存在“三低”现象,即抽穗率低,大约7.9%;结实率低,自然条件下一般为25%;种子发芽率低,为10%~20%[28-30]。另外,羊草种群有性繁殖能力的差异主要与气候因子[29]及土壤养分条件[31]有关。杨允菲应用正交试验观测氮肥、磷肥、灌水和处理时间对羊草种子产生的效果,结果发现磷肥、氮肥和灌水均使每穗籽粒数、结实率、千粒重显著下降[31]。而本研究中各个密度处理的羊草均种植在相同的土壤环境下,且不存在气候差异,因此,本研究的有性繁殖能力也无明显差异。
潜在种群中,根茎顶端芽和根茎节间芽的数量代表种群向外扩展的潜在能量,而分蘖节芽数量则代表种群占据已有空间的潜在能量。本研究中,羊草潜在种群随着现实种群的增加而呈增加趋势。其中,根茎顶端芽与地上生物量之间的相关系数较高,为0.717,根茎节间芽与地下生物量之间的相关系数较高,为0.648。说明,随着密度的增加,羊草种群地上和地下生物量降低的同时,其潜在种群向外扩展的能力也降低,使未来种群的数量趋于减少。
综合以上研究表明,对于不同播种密度,羊草首先调节种群的数量、生物量及年龄结构,继而调节潜在种群的数量,使未来种群数量维持在较适宜的水平,即较低的播种密度不但有利于羊草种群的年龄结构呈现相对较快趋于稳定的趋势,而且还有利于种群数量和生物量的增长及空间扩展。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
数学物理学报(2021年6期)2021-12-21 06:24:52
现代畜牧科技(2021年7期)2021-07-28 06:40:30
现代畜牧科技(2021年4期)2021-07-21 06:13:04
河南农业大学学报(2020年2期)2020-05-22 02:56:30
数学物理学报(2019年5期)2019-11-29 07:47:04
山西林业科技(2019年1期)2019-05-27 12:24:52
数学年刊A辑(中文版)(2016年1期)2016-10-30 01:46:20
海洋渔业(2016年6期)2016-04-16 03:00:35
现代畜牧科技(2016年9期)2016-02-21 12:25:04
