
HPV16阳性和阴性患者宫颈癌组织中miR-27a-3p和HOXB8蛋白的表达差异及其机制研究
2017-09-07 02:14李会影王慧智张子旸牡丹江医学院附属红旗医院妇产科黑龙江牡丹江157011
中国药房 2017年23期
李会影,王慧智,张子旸(牡丹江医学院附属红旗医院妇产科,黑龙江牡丹江 157011)
·精准医疗·
HPV16阳性和阴性患者宫颈癌组织中miR-27a-3p和HOXB8蛋白的表达差异及其机制研究
李会影*,王慧智,张子旸#(牡丹江医学院附属红旗医院妇产科,黑龙江牡丹江 157011)
目的:探讨miR-27a-3p和同源盒B8(HOXB8)蛋白在人乳头瘤病毒16型阳性[HPV16(+)]和阴性[HPV16(-)]患者宫颈癌组织中表达的差异及其机制。方法:选取2012年1月-2016年1月于我院就诊的宫颈癌患者120例,根据其是否感染HPV16分为HPV16(+)组(60例)和HPV16(-)组(60例)。采用实时荧光定量聚合酶链反应法检测两组患者宫颈癌组织中miR-27a-3p和HOXB8 mRNA的表达水平,采用蛋白印迹法检测HOXB8蛋白的表达水平,采用巢式降落式甲基化特异性聚合酶链反应法检测miR-27a-3p启动子区DNA甲基化水平,采用染色质免疫共沉淀-定量聚合酶链反应法检测miR-27a-3p启动子区组蛋白甲基化水平;培养人宫颈癌细胞系SiHa细胞株,考察转染miR-27a-3p模拟物(mimic)和抑制物(inhibitor)后miR-27a-3p和HOXB8 mRNA的表达水平。结果:HPV16(+)组患者宫颈癌组织中miR-27a-3p的表达水平显著低于HPV16(-)组患者,HOXB8 mRNA和蛋白表达水平、miR-27a-3p启动子区DNA甲基化水平和组蛋白甲基化水平均显著高于HPV16(-)组患者,差异均有统计学意义(P<0.05或P<0.01)。在SiHa细胞转染miR-27a-3p mimic后,miR-27a-3p的表达水平显著升高,而HOXB8 mRNA的表达水平显著降低;转染miR-27a-3p inhibitor后,miR-27a-3p的表达水平显著降低,而HOXB8 mRNA的表达水平则显著升高,差异均有统计学意义(P<0.01)。结论:HPV16可能通过启动子区DNA甲基化和组蛋白甲基化下调miR-27a-3p的表达,从而影响宫颈癌的发生与发展,HOXB8蛋白可能是其作用靶点。
人乳头瘤病毒16型;宫颈癌;miR-27a-3p;同源盒B8蛋白;DNA甲基化;组蛋白甲基化
宫颈癌是女性生殖系统恶性肿瘤,在世界范围内的发病率和病死率均较高,严重威胁女性生命健康[1-2]。人乳头瘤病毒16型(Human papilloma virus type 16,HPV16)是最普遍、最致命的HPV类型之一,也是诱发宫颈癌的主要原因[3]。同源盒B8(Homeobox B8,HOXB8)是HOX蛋白家族成员,其表达水平在HPV16阳性[HPV16(+)]宫颈癌组织中显著升高,但具体机制尚未阐明[4]。微RNA(MicroRNAs,miRNAs)是一类内源性小分子非编码RNA,可在转录后调控基因的表达[5]。研究表明,多种miRNAs通过调控靶基因的表达参与宫颈癌的发生与发展,其中miR-27a-3p可调控HOXB8基因的表达[6-8]。现有研究多集中于miRNAs的调控作用,而忽视了miRNAs表达变化的机制。DNA甲基化和组蛋白甲基化是表观遗传学调控的重要方式[9-10],其是否参与miRNAs的表达调控仍不清楚。因此,本研究以HPV16(+)和HPV16阴性[HPV16(-)]宫颈癌患者为研究对象,探讨miR-27a-3p的表达变化及其机制,并考察其对HOXB8蛋白是否具有调控作用。
1 资料与方法
1.1 材料
1.1.1 仪器 QuantStudio Q3型实时荧光定量聚合酶链反应(PCR)仪(美国Applied Biosystems公司);Mastercycler Nexus Eco型普通PCR仪、547R型台式高速冷冻离心机(德国Eppendorf公司);Powerpac HV Power Supply型电泳仪(美国BioRad公司);FCM型凝胶成像系统(美国Protein Simple公司);DLHR-D1603型电动匀浆器(华邻科技有限公司);HR900-ⅡA2型生物安全柜(海尔生物医疗)。
1.1.2 试剂 胎牛血清(FBS)、opti-MEM无血清培养基、青链霉素和高糖培养基(DMEM)(美国Gibco公司,批号分别为12483-012、51985042、15240-062、12491-015);磷酸盐缓冲液(PBS)(美国Hyclone公司,批号:SH30256.01B);聚山梨酯20(美国BioRad公司,批号:170-6531);DNA提取试剂盒、总RNA提取试剂盒、荧光定量PCR试剂盒(含SYBR Green、ddH2O)(美国Qiagen公司,批号分别为69506、74104、204054);逆转录试剂盒及发光液(美国ThermoFisher公司,批号分别为K1691、32106);DNA甲基化修饰试剂盒(北京天漠科技开发有限公司,批号:D5030);miRNA提取试剂盒、miRNA逆转录试剂盒、miRNA荧光定量PCR试剂盒、miR-27a-3p及U6引物(天根生化科技有限公司,批号分别为DP501、KR201-02、FP401-02、CD201-0025、CD201-0145);蛋白质提取及定量试剂盒、蛋白上样缓冲液(江苏凯基生物技术股份有限公司,批号分别为KGP250、KGPBCA、KGP101);anti-HOXB8、anti-H3K9me2(美国Abcam公司,批号分别为ab125727、ab120);anti-β-actin及辣根过氧化物酶(HRP)标记二抗(北京中杉金桥生物技术有限公司,批号分别为TA-09、SC-2048);人工合成miR-27a-3p模拟物(mimic)、抑制物(inhibitor)及阴性对照(上海吉玛制药技术有限公司);Lipo2000(美国Invitrogen公司,批号:11668027);染色质免疫共沉淀试剂盒[含蛋白A/G琼脂糖(Protein G Agarose),美国Millipore公司,批号:17-371];普通引物由华大基因提供,HOXB8、甘油醛-3-磷酸脱氢酶(GAPDH)引物及甲基化特异性引物由上海生工生物工程股份有限公司合成;人宫颈癌细胞系Si-Ha细胞株购自美国模式培养物集存库。
1.2 研究对象
纳入标准:(1)于我院首次就诊或接受治疗的宫颈癌患者,经组织活检及术后病理学检查确诊为宫颈癌;(2)既往无其他恶性肿瘤病史;(3)不合并其他恶性肿瘤;(4)术前未接受放疗、化疗或其他治疗。排除标准:(1)年龄≤18岁者;(2)妊娠期妇女;(3)严重的肝肾功能障碍者;(4)合并不能有效控制的高血压、糖尿病及心脏疾病等。本研究方案经医院医学伦理委员会审核通过,患者均知情同意并签署知情同意书。
本研究共纳入2012年1月—2016年1月于我院妇产科就诊的宫颈癌患者120例,根据其是否感染HPV16分为HPV16(+)组和HPV16(-)组。其中,HPV16(+)组患者60例,年龄28~63岁,平均年龄(47.2±9.5)岁;HPV16(-)组患者60例,年龄25~59岁,平均年龄(45.7±8.3)岁。所有患者均留取手术切除的癌组织标本,于-80℃冻存,备用。
1.3 研究方法
1.3.1 实时荧光定量PCR法检测miR-27a-3p和HOXB8 mRNA的表达水平 按照试剂盒说明书提取宫颈癌组织中的总miRNA和总mRNA,并将其逆转录为互补脱氧核糖核酸(cDNA)。其中,总miRNA的逆转录分为加多聚腺苷酸(PolyA)尾和逆转录两部分。以cDNA为模板,进行实时荧光定量PCR分析,反应体系包括cDNA 2 μL,上、下游引物各0.8 μL,SYBR Green 12.5 μL,ddH2O 8.9 μL,共25 μL。反应条件:95℃预变性5 min,1个循环;95℃变性10 s,58℃退火30 s,72℃延伸30 s,40个循环;产物经熔解曲线分析。HOXB8基因上游引物:5′-GTCCCTGCGCCCCAATTATTA-3′,下游引物:5′-GCCCGTGGTAGAACTCCTG-3′。HOXB8 mRNA相对表达水平以GAPDH的表达水平为参照,根据目的基因的相对表达量=2-ΔΔCt计算结果,ΔΔCt=[CtHOXB8-CtGAPDH],计算HOXB8 mRNA的相对表达变化。miR-27a-3p相对表达水平以U6的表达水平为参照,根据目的基因的相对表达量=2-ΔΔCt计算结果,ΔΔCt=[CtmiR-27a-3p-CtU6],计算miR-27a-3p的相对表达变化。式中,Ct(Cycle threshold)表示每个反应管内的荧光信号到达设定的阈值时所经历的循环数。
1.3.2 蛋白质印迹(Western blotting)法检测HOXB8蛋白的表达水平 将宫颈癌组织剪碎,冰上匀浆,以离心半径6 cm、转速5 000 r/min离心5 min,弃去上清,收集沉淀。按照蛋白提取试剂盒说明书提取宫颈癌组织的总蛋白,并进行蛋白定量。按体积比1∶4加入蛋白上样缓冲液混匀,于100℃下反应5 min,使蛋白变性。取变性后的蛋白样品5 μL,进行十二烷基硫酸钠-聚丙烯酰胺(SDS-PAGE)凝胶电泳,切取大小约为32 kDa(HOXB8)和43 kDa(β-actin)的片段,分别进行半干转膜操作。转膜结束后,采用5%脱脂奶粉对聚偏二氟乙烯膜(PVDF)封闭2 h。分别与anti-HOXB8和anti-β-actin混合后,于4℃下孵育过夜。用磷酸缓冲盐溶液-聚山梨酯(PBST,由PBS和聚山梨酯20配制)洗膜后,与HRP标记的二抗于室温下孵育2 h,PBST洗膜后加发光液,于凝胶成像分析仪上成像并分析条带灰度值。以β-actin的表达量为参照,计算HOXB8蛋白的相对表达水平。HOXB8蛋白的相对表达水平=HOXB8灰度值/βactin灰度值。
1.3.3 巢式降落式甲基化特异性PCR(nMS-PCR)法检测miR-27a-3p启动子区DNA甲基化水平 按照试剂盒说明书提取宫颈癌组织的基因组DNA,并采用亚硫酸氢盐法对基因组DNA进行甲基化修饰。在Ensembl Genome Browser(http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi)中查找miR-27a-3p启动子区序列并线上设计1对外引物和2对内引物(即甲基化引物和非甲基化引物)。外引物上游:5′-GGTTTTGTGTAGGGGAAATTTTATAG-3′,外引物下游:5′-CAACCCTATCCCCTTAAAATCTAAA-3′;甲基化引物上游:5′-TTGGGGTGAGATTATTTTTTTATTC-3′,甲基化引物下游:5′-CTAAAACCTAACTATACCCTCCGTT-3′;非甲基化引物上游:5′-GGGGTGAGATTATTTTTTTATTTGT-3′,非甲基化引物下游:5′-CTAAAACCTAACTATACCC-TCCATT-3′。外引物扩增的反应条件:95℃预变性5 min;95℃变性30 s,61℃退火30 s,72℃延伸30 s,20个循环,每个循环降0.5℃至52℃,72℃再延伸7 min。以外引物的PCR产物为模板,继续进行内引物的扩增,其扩增反应条件同外引物。取PCR产物5 μL于2%琼脂糖凝胶上进行电泳分析,并于凝胶成像分析系统成像,计算甲基化条带及非甲基化条带的光密度(OD),miR-27a-3p启动子区DNA甲基化水平=甲基化条带的OD值/(甲基化条带的OD值+非甲基化条带的OD值)。
1.3.4 染色质免疫共沉淀-定量PCR(CHIP-qPCR)法检测miR-27a-3p启动子区组蛋白甲基化水平 将宫颈癌组织冰上匀浆后,以终体积分数为1%的甲醛室温固定10 min。采用终浓度为0.125 mol/L甘氨酸作用5 min以终止交联。以离心半径6 cm、转速5 000 r/min离心5 min后,弃去上清,PBS冲洗2遍。SDS裂解液重悬并裂解组织沉淀,冰上进行超声破碎操作,使DNA破碎为100~1 000 bp大小的片段。以离心半径6 cm、转速12 000 r/min离心15 min后取上清液100 μL,加入anti-H3K9me2抗体,于4℃下孵育过夜,形成anti-H3K9me2-H3K9me2复合物;经蛋白A/G琼脂糖结合沉淀后,形成Protein G Agarose-anti-H3K9me2-H3K9me2复合物,于65℃水浴中解除交联,并采用DNA提取试剂盒、按说明书操作纯化DNA,以纯化后的DNA为模板,对miR-27a-3p启动子区进行实时荧光定量PCR扩增反应。扩增反应体系及反应条件同“1.3.1”项,退火温度为59℃。
1.3.5 细胞培养及转染 用含10%FBS及1%抗生素的DMEM培养基,于37℃、5%CO2的培养箱中静置培养SiHa细胞株。按每孔1×106个细胞铺板:①取100 μL opti-MEM与适量Lipo2000混匀静置5 min;②另取100 μL opti-MEM与适量miR-27a-3p mimic、inhibitor或阴性对照混匀静置5 min;③将上述①②混匀后静置20 min,加至细胞培养液中,于48 h后收集细胞,提取RNA进行后续试验,步骤同“1.3.1”项。
1.4 统计学方法
应用SPSS 17.0软件对数据进行统计分析。计量资料以±s表示,两组间比较采用Students’t检验,多组间比较采用单因素方差分析,组间两两比较采用SNK检验。P<0.05为差异有统计学意义。
2 结果
2.1 两组患者宫颈癌组织中miR-27a-3p表达水平比较
HPV16(+)组患者宫颈癌组织中miR-27a-3p的表达水平显著低于HPV16(-)组,差异有统计学意义(P<0.01),详见图1。
2.2 两组患者宫颈癌组织中HOXB8 mRNA和蛋白表达水平比较

图1 两组患者宫颈癌组织中miR-27a-3p表达水平比较Fig 1 Comparison of the expression of miR-27a-3p in cervical cancer tissue between 2 groups
HPV16(+)组患者宫颈癌组织中HOXB8 mRNA和蛋白的表达水平均显著高于HPV16(-)组患者,差异均有统计学意义(P<0.05),详见图2。
2.3 两组患者宫颈癌组织中miR-27a-3p启动子区DNA甲基化水平比较
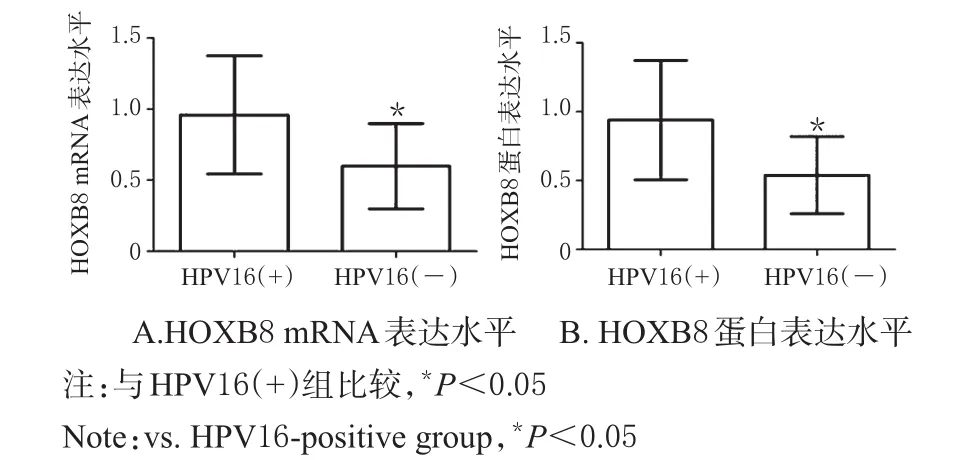
图2 两组患者宫颈癌组织中HOXB8 mRNA和蛋白表达水平比较Fig 2 Comparison of the expression of HOXB8 mRNA and protein in cervical cancer tissues between 2 groups
在线(http://www.urogene.org/methprimer/)查看miR-27a-3p启动子区CpG位点和CpG岛分布情况,结果显示,miR-27a-3p启动子区富含CpG岛,详见图3。
HPV16(+)组患者宫颈癌组织中miR-27a-3p启动子区DNA甲基化水平显著高于HPV16(-)组患者,差异有统计学意义(P<0.01),详见图4、图5。

图3 miR-27a-3p启动子区CpG位点和CpG岛分布情况Fig 3 Distribution of CpG islands and CpG sites in miR-27a-3p promoter region
2.4 两组患者宫颈癌组织中miR-27a-3p启动子区组蛋白甲基化水平比较

图4 nMS-PCR产物电泳图Fig 4 Electrophoretogram of nMS-PCR product
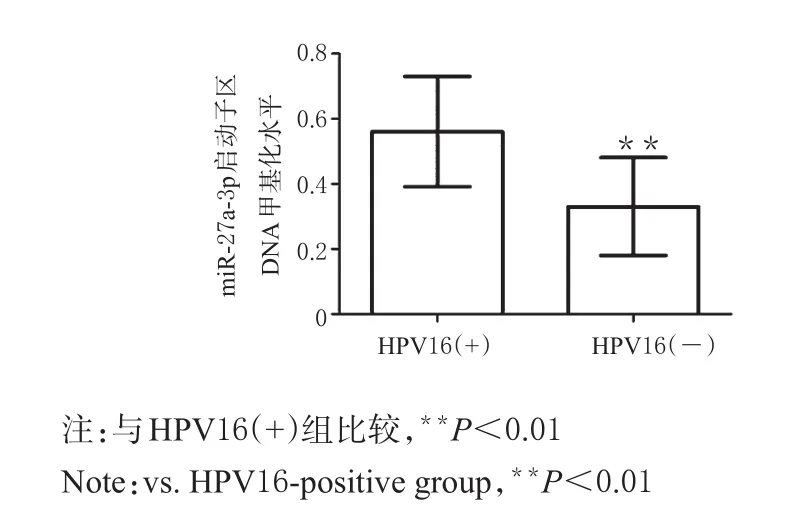
图5 两组患者宫颈癌组织中miR-27a-3p启动子区DNA甲基化水平比较Fig 5 Comparison of DNA methylation levels of miR-27a-3p promoter region in cervical cancer tissues between 2 groups
HPV16(+)组患者宫颈癌组织中miR-27a-3p启动子区组蛋白甲基化水平显著高于HPV16(-)组患者,差异有统计学意义(P<0.05),详见图6。
2.5 miR-27a-3p对HOXB8表达的影响
在人宫颈癌细胞系SiHa细胞株中分别转染miR-27a-3p mimic和inhibitor后,检测miR-27a-3p和HOXB8 mRNA表达的变化。结果显示,在SiHa细胞转染miR-27a-3p mimic后,miR-27a-3p的表达水平显著升高,而HOXB8 mRNA的表达水平显著降低,差异均有统计学意义(P<0.01);转染miR-27a-3p inhibitor后,miR-27a-3p的表达水平显著降低,而HOXB8 mRNA的表达水平则显著升高,差异均有统计学意义(P<0.01),详见图7。
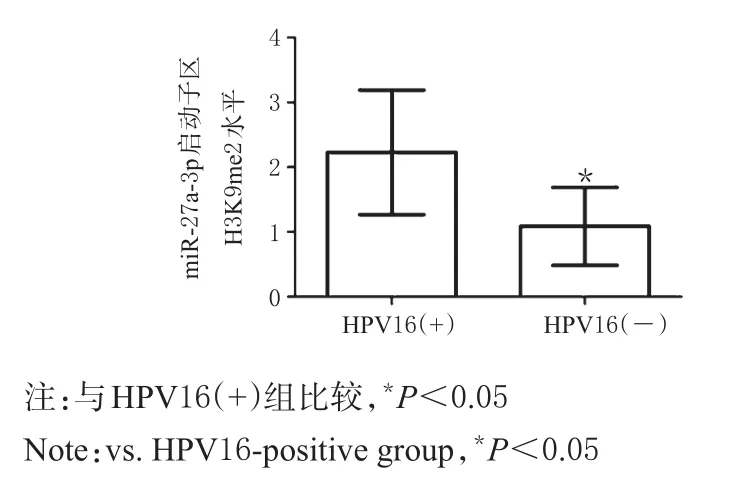
图6 两组患者宫颈癌组织中miR-27a-3p启动子区组蛋白甲基化水平比较Fig 6 Comparison of histone methylation levels of miR-27a-3p promoter region in cervical cancer tissues between 2 groups
3 讨论
宫颈癌是女性生殖系统恶性肿瘤,HPV感染是引起宫颈癌的主要原因之一。HPV是一种双链DNA病毒,低危险性HPV可引起良性的生殖器疣,而高危险性HPV与宫颈癌的发生与发展密切相关。其中,HPV16是HPV最常见的类型,是导致宫颈癌的重要因素[3,11-12]。本研究以HPV16(+)和HPV16(-)宫颈癌患者为研究对象,初步探讨HPV16的致病机制。
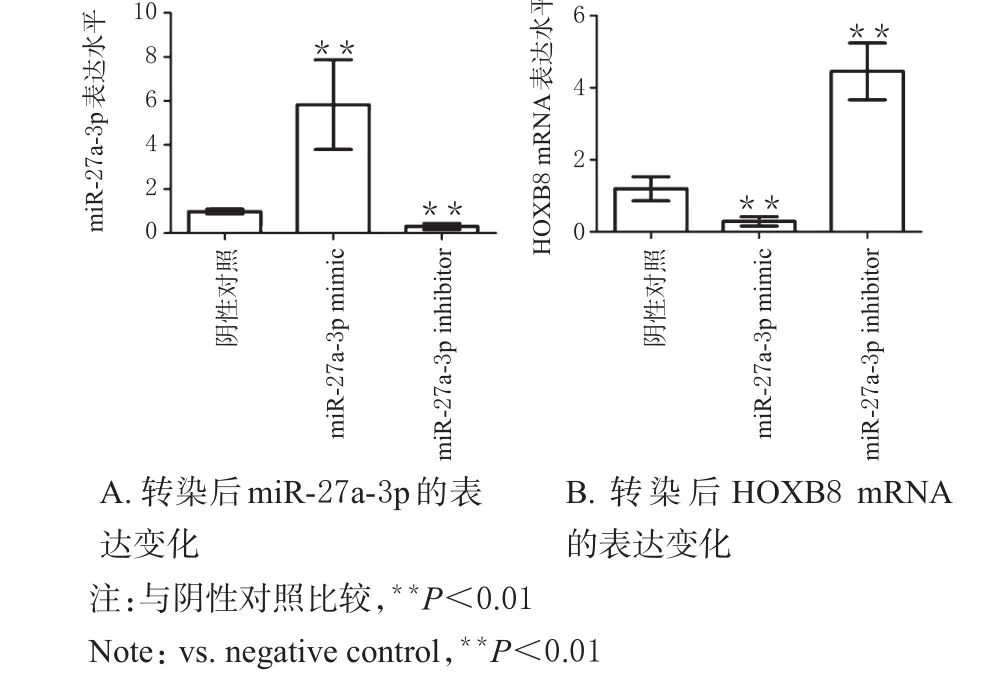
图7 miR-27a-3p对HOXB8表达的影响Fig 7 Effect of miR-27a-3p on the expression of HOXB8
3.1 HOXB8与miRNAs
HOXB8是同源盒家族成员,能够编码具有HOX DNA结合结构域的核蛋白。研究表明,HOXB8与结直肠癌、胃癌和宫颈癌的发生、发展密切相关[4,13-14]。本研究结果显示,在HPV16(+)宫颈癌患者组织中HOXB8的表达水平显著升高,与文献[4]报道一致,但其具体机制尚不清楚。miRNAs是一类大小约为18~22个核苷酸的内源性小分子非编码RNA,可通过结合靶mRNA 3′-非翻译(UTR)区使其降解或翻译一致,从而实现在转录后调控基因表达的目的。Yang H等[15]研究发现,miR-497可靶向转酮酶调控宫颈癌对铂化疗的敏感性;Zhou N等[16]研究发现,miR-138可通过调控人端粒逆转录酶(hTERT)的表达水平抑制宫颈癌细胞的增殖、迁移和侵袭;Lai XJ等[17]研究表明,miR-421可通过下调Bcl-xL从而介导c-33a宫颈癌细胞的凋亡,这提示多种miRNAs可通过调控靶基因的表达参与宫颈癌的发生与发展。本研究结果显示,在人宫颈癌细胞系SiHa细胞株中,转染miR-27a-3p mimic(即过表达miR-27a-3p)后,HOXB8 mRNA的表达水平显著降低;而转染miR-27a-3p inhibitor(即抑制miR-27a-3p的表达)后,HOXB8 mRNA的表达水平显著升高,提示miR-27a-3p可能直接或间接靶向调控HOXB8的表达,但具体机制尚有待进一步研究。
3.2 miRNAs与表观遗传学修饰
以往研究多集中于miRNAs对其靶基因的调控作用,而忽视了miRNAs表达变化的机制。表观遗传学修饰是指在基因核苷酸序列不发生改变的情况下,基因表达发生可逆、可遗传的改变。表观遗传的现象很多,已知的有DNA甲基化(DNA methylation)、组蛋白甲基化(Histone methylation)、基因组印记(Genomic imprinting)、母体效应(Maternal effects)、基因沉默(Gene silencing)、核仁显性、休眠转座子启动和RNA编辑(RNAediting)等[18]。其中,DNA甲基化和组蛋白甲基化是表观遗传学调控的重要方式,文献报道DNA甲基化和H3K9me2均为基因沉默的标志,且二者可能具有协同调控基因表达的作用[9,19]。赵丽等[20]研究表明,miR-124启动子区DNA甲基化在动脉粥样硬化的发生与发展过程中发挥了重要的作用。然而,DNA甲基化和组蛋白甲基化是否参与HPV16(+)宫颈癌组织miR-27a-3p的表达调控仍不清楚。本研究结果显示,HPV16(+)组患者宫颈癌组织中miR-27a-3p启动子区DNA和组蛋白甲基化水平均显著高于HPV16(-)组患者,提示DNA甲基化和组蛋白甲基化参与了HPV16(+)宫颈癌组织中miR-27a-3p的表达调控。DNA甲基化和组蛋白甲基化的发生需要相应酶的催化,如DNA甲基转移酶1(DNMT1)、组蛋白甲基化酶G9a等,但其在HPV16(+)宫颈癌中是否发生改变尚需进一步研究。
综上所述,HPV16可能通过启动子区DNA甲基化和组蛋白甲基化下调miR-27a-3p的表达,从而影响宫颈癌的发生与发展;HOXB8蛋白可能是其作用靶点,HOXB8基因可能是miR-27a-3p的靶基因。本研究为HPV16(+)宫颈癌患者的临床治疗提供了理论基础和治疗靶点。但miR-27a-3p启动子区发生DNA甲基化和组蛋白甲基化的机制仍不明确,还需后续深入研究。
[1] Yamagami W,Nagase S,Takahashi F,et al.Clinical statistics of gynecologic cancers in Japan[J].J Gynecol Oncol,2017,28(2):e32.
[2] Hasahya OT,Berggren V,Sematimba D,et al.Beliefs,perceptions and health-seeking behaviours in relation to cervical cancer:a qualitative study among women in Uganda following completion of an HPV vaccination campaign[J].Glob Health Action,2016,doi:10.3402/gha. v9.29336.
[3] Torres Lobatón A,Rojo Herrera G,Torres Rojo A,et al. Cervical cancer:current view of its epidemiology and risk factors[J].Ginecol Obstet Mex,2004,72:466-474.
[4] Liu C,Lin J,Li L,et al.HPV16 early gene E5 specifically reduces miRNA-196a in cervical cancer cells[J].Sci Rep,2015,doi:10.1038/srep07653.
[5] Barwari T,Joshi A,Mayr M.MicroRNAs in cardiovascular disease[J].J Am Coll Cardiol,2016,68(23):2577-2584.
[6] Xu D,Liu S,Zhang L,et al.MiR-211 inhibits invasion and epithelial-to-mesenchymal transition(EMT)of cervical cancer cells via targeting MUC4[J].Biochem Biophys Res Commun,2017,485(2):556-562.
[7] Fan JY,Fan YJ,Wang XL,et al.MiR-429 is involved in regulation of NF-κB activity by targeting IKKβ and suppresses oncogenic activity in cervical cancer cells[J]. FEBS Lett,2017,591(1):118-128.
[8] Shi C,Zhang Z.MicroRNA-362 is downregulated in cervical cancer and inhibits cell proliferation,migration and invasion by directly targeting SIX1[J].Oncol Rep,2017,37(1):501-509.
[9] Estève PO,Chin HG,Smallwood A,et al.Direct interaction between DNMT1 and G9a coordinates DNA and histone methylation during replication[J].Genes Dev,2006,20(22):3089-3103.
[10] Hashimoto H,Vertino PM,Cheng X.Molecular coupling of DNA methylation and histone methylation[J].Epigenomics,2010,2(5):657-669.
[11] Schiffman M,Castle PE,Jeronimo J,et al.Human papillomavirus and cervical cancer[J].Lancet,2007,370(9590):890-907.
[12] Zhao JW,Fang F,Guo Y,et al.HPV16 integration probably contributes to cervical oncogenesis through interrupting tumor suppressor genes and inducing chromosome instability[J].J Exp Clin Cancer Res,2016,35(1):180.
[13] Shen S,Pan J,Lu X,et al.Role of miR-196 and its target gene HOXB8 in the development and proliferation of human colorectal cancer and the impact of neoadjuvant chemotherapy with FOLFOX4 on their expression[J].Oncol Lett,2016,12(5):4041-4047.
[14] Ding WJ,Zhou M,Chen MM,et al.HOXB8 promotes tumor metastasis and the epithelial-mesenchymal transition via ZEB2 targets in gastric cancer[J].J Cancer Res Clin Oncol,2017,143(3):385-397.
[15] Yang H,Wu XL,Wu KH,et al.MicroRNA-497 regu-lates cisplatin chemosensitivity of cervical cancer by targeting transketolase[J].Am J Cancer Res,2016,6(11):2690-2699.
[16] Zhou N,Fei D,Zong S,et al.MicroRNA-138 inhibits proliferation,migration and invasion through targeting hTERT in cervical cancer[J].Oncol Lett,2016,12(5):3633-3639.
[17]Lai XJ,Cheng XY,Hu LD.MicroRNA 421 induces apoptosis of c-33a cervical cancer cells via down-regulation of Bcl-xL[J].GenetMolRes,2016,doi:10.4238/gmr15048853.
[18] Russo V,Bernabò N,Di Giacinto O,et al.H3K9 trimethylation precedes DNA methylation during sheep oogenesis:HDAC1,SUV39H1,G9a,HP1,and Dnmts are involved in these epigenetic events[J].J Histochem Cytochem,2013,61(1):75-89.
[19] Leung DC,Dong KB,Maksakova IA,et al.Lysine methyltransferase G9a is required for de novo DNA methylation and the establishment,but not the maintenance,of proviral silencing[J].Proc Natl Acad Sci U S A,2011,108(14):5718-5723.
[20]赵丽,焦运,杨安宁,等.MiR-124及其启动子区DNA甲基化在同型半胱氨酸致动脉粥样硬化中的作用[J].生理学报,2015,67(2):207-213.
(编辑:张元媛)
Expression Difference and Its Mechanism Study of miR-27a-3p and HOXB8 Protein in Cervical Cancer Tissues of HPV16-positive and HPV16-negative Patients
LI Huiying,WANG Huizhi,ZHANG Ziyang(Dept.of Obstetrics and Gynecology,Hongqi Hospital Affiliated to Mudanjiang Medical College,Heilongjiang Mudanjiang 157011,China)
OBJECTIVE:To investigate the expression difference and its mechanism of miR-27a-3p and HOXB8 protein in cervical cancer tissues of HPV16-positive and HPV16-negative patients.METHODS:A total of 120 patients with cervical cancer in our hospital during Jan.2012-Jan.2016 were divided into HPV16-positive group(60 cases)and HPV16-negative group(60 cases)according to HPV16 infection situation.The expression of miR-27a-3p mRNA and HOXB8 mRNA in cervical cancer tissue were detected by RT-qPCR.The expression of HOXB8 protein was detected by Western blotting assay.DNA methylation level of miR-27a-3p promoter region was detected by nested-down methylation specific PCR(nMS-PCR).Histone methylation level of miR-27a-3p promoter region was detected by chromatin immunoprecipitation PCR(CHIP-PCR).The expression of miR-27a-3p mRNA and HOXB8 mRNA were detected after transfecting miR-27a-3p mimic and inhibitor into Human cervical cancer cell line SiHa,respectively.RESULTS:The expression of miR-27a-3p in HPV16-positive group was significantly lower than HPV16-negative group,while HOXB8 mRNA and protein expression,DNA and histone methylation levels of miR-27a-3p promoter region were significantly higher than HPV16-negative group,with statistical significance(P<0.05 or P<0.01).After transfecting miR-27a-3p mimic into SiHa cells,the expression of miR-27a-3p was increased significantly,while that of HOXB8 mRNA was decreased significantly;after miR-27a-3p inhibitor transfection,the expression of miR-27a-3p was decreased significantly,while that of HOXB8 mRNA was increased significantly,with statistical significance(P<0.01).CONCLUSIONS:HPV16 may down-regulate the expression of miR-27a-3p through DNA methylation and histone methylation of promoter region,so as to influence the generation and development of cervical cancer.HOXB8 may be the target protein of miR-27a-3p.
Human papilloma virus type 16;Cervical cancer;miR-27a-3p;Homeobox B8 protein;DNA methylation;Histone methylation
R711.74
A
1001-0408(2017)23-3169-06
2016-12-22
2017-06-15)
*副主任医师,硕士。研究方向:妇产科学。电话:0453-6602036。E-mail:huiying26@163.com
#通信作者:副主任医师。研究方向:妇产科学。电话:0453-6602036。E-mail:1403355759@qq.com
DOI10.6039/j.issn.1001-0408.2017.23.01
猜你喜欢
航天电子对抗(2022年4期)2022-10-24
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年1期)2022-02-28
中老年保健(2021年12期)2021-08-24
中华养生保健(2020年10期)2021-01-18
中国生殖健康(2020年7期)2021-01-18
中学生物学(2020年10期)2020-12-25
中国科技纵横(2018年2期)2018-11-29
医药前沿(2018年8期)2018-03-15
医学研究杂志(2015年12期)2015-06-10
