
大型海藻碳汇能力的种间差异
2017-08-30 12:48欧官用王鑫杰杨安强柯爱英关万春
浙江农业科学 2017年8期
欧官用,王鑫杰,杨安强,柯爱英,关万春
(1.浙江省海洋水产养殖研究所,浙江 温州 325005; 2.温州医科大学 生命科学学院海洋生物技术系,浙江 温州 325035)
大型海藻碳汇能力的种间差异
欧官用2,王鑫杰2,杨安强2,柯爱英1*,关万春2
(1.浙江省海洋水产养殖研究所,浙江 温州 325005; 2.温州医科大学 生命科学学院海洋生物技术系,浙江 温州 325035)
对浙南沿岸常见的11种大型海藻(2种绿藻,2种红藻和7种褐藻)进行研究,探讨其碳汇能力对光的响应差异,并初步了解该11种大型海藻在渔业碳汇中的作用。结果表明,这11种大型海藻的固碳能力对光响应的关系均符合方程y=Pm×tanh(α×x÷Pm)+Rd,且光合固碳速率存在种间差异,表现为海带>萱藻>鼠尾藻>珊瑚藻>铜藻>浒苔>瓦氏马尾藻>蜈蚣藻>石莼>铁钉菜>羊栖菜。其中,海带的固碳能力最强,固碳量可达0.89×10-3t·t-1·h-1,相当于可减排的最大CO2量达3.28×10-3t·t-1·h-1。羊栖菜的固碳能力最低,固碳量为0.28×10-3t·t-1·h-1,可减排的最大CO2量为1.04×10-3t·t-1·h-1,固碳能力仅为海带减排CO2能力的31.5%。通过对比发现,大型经济海藻海带对渔业碳汇的贡献最为明显,碳汇能力所能创造的经济价值为0.49~1.97美元·t-1·h-1。虽然其他海藻的固碳能力均低于海带,但由于其生物量大,分布较广,对于渔业碳汇的贡献也极其重要。因此,合理养殖大型海藻、保护大型海藻场及维护大型海藻的生物多样性,对提高渔业碳汇能力、增加CO2减排量具有重要作用。
大型海藻; 渔业碳汇; CO2; 光合固碳
人类活动使用大量的化石燃料,使大气中CO2的浓度持续升高,预计2100年大气中CO2的浓度达1.96 μg·L-1[1-4],该数值是目前大气中CO2浓度的2.5倍。同时,伴随着大气中CO2浓度的持续升高,全球变暖的速度也将进一步加剧。因此,如何减缓全球变暖的速度以及降低大气中的CO2浓度,已经成为全球关注的热点。大型海藻,作为海洋初级生产力中的重要组成部分,一方面可以通过光合作用调控和吸收大气中的CO2,直接影响全球碳循环;另一方面通过其自身具有的碳汇功能,吸收了人类排放CO2总量的20%~35%,从而有效降低了大气中的CO2浓度以及延缓了全球气候变化的速度[5-7]。大型海藻主要具有气候调节、固碳制氧、缓解水体富营养化、净化环境等生态功能[8]。据统计, 沿海生态系统是所有生态系统中经济价值最高的生态系统, 每年的生态服务价值已超过20万亿美元, 而这一值仅是一个偏低的估计值[9]。因此,以浙南地区采集的11种大型海藻为研究对象,探讨其固碳能力对光的响应机制,拟掌握11种大型海藻的碳汇能力和对渔业碳汇的贡献,并分析碳汇能力的种间差异。
1 材料与方法
1.1 材料
供试11种大型海藻均采自浙南地区。绿藻有石莼(Ulvalactuca)和浒苔(Enteromorphaprolifera),红藻有蜈蚣藻(Grateloupiafilicina)和珊瑚藻(Corallinaofficinalis),褐藻有羊栖菜(Sargassumfusiforme)、铁钉菜(Ishigeokamurai)、铜藻(S.horneri)、瓦氏马尾藻(S.vachellianum)、海带(Laminariajaponica)、萱藻(Scytosiphonlomentarius)和鼠尾藻(S.thunbergii)。选择健康的藻体,除去表面附着物,放置于培养箱中适应培养。适应期水温(20±0.5)℃,盐度24%,光照强度100 μmol·m-2·s-1,光暗周期为12 L∶12D。
1.2 固碳能力测定
采用氧电极法(Model 782 with 1302 oxygen electrode, Strathkelvin Instruments, 苏格兰)测定大型海藻固碳能力。由于藻体的光合固碳速率和光合放氧速率为1∶1,因此,藻体每释放1mol氧气,就相当于固定1mol碳,从而可以通过测得的光合放氧速率获得光合固碳速率。取海藻样品0.1 g,放于盛有3 mL无菌海水的氧电极反应杯中,通过水浴(Model:F12, Gesellschaft mit beschränkter Haftung,德国)控温20 ℃。光源为卤灯(QVF137,Philips,中国),通过调节光源与氧电极反应杯间的距离来获得0(关闭光源,并用黑布遮光)、20、50、100、200和300 μmol·m-2·s-1的6个光强梯度,并测定不同光照强度条件下藻体的光合放氧速率(固碳速率)及呼吸速率。光合固碳速率(P)与光的关系参考P公式进行拟合[10],每吨鲜重每小时可达到的最大固碳(碳汇)能力(Cm)采用Cm公式计算。
PCarbon=POxygen=PmαI/Pm+Rd;
Cm=(Pm+|Rd|)×12/1 000。
式中,I表示光照强度,PCarbon是光照强度为I时所对应的光合固碳速率,POxygen是光照强度为I时所对应的光合放氧速率,Pm是最大光合放氧速率,即最大光合固碳速率,Rd为呼吸速率,α为光能利用率,Cm是大型海藻每吨鲜重每小时可达到的最大碳汇能力。
α=Pm/Ik。
式中Ik为光饱和参数。
1.3 大型海藻每吨鲜重每小时可减排的最大CO2量
Cm×44/12。
1.4 大型海藻碳汇能力所能创造的经济价值
依据《联合国气候变化框架公约的京都议定书》确定的工业化国家减排CO2的开支标准:150~600美元·t-1,即可获得大型海藻每小时每吨鲜重藻体可创造的经济价值,而最终得出的数据有利于为当地渔业部门核算大型海藻减排CO2的潜在总经济价值提供依据。
1.5 数据处理
采用Origin8.0和SPSS软件对数据进行处理分析。
2 结果与分析
2.1 光合固碳能力与光的关系
11种大型海藻的固碳能力(y)对光照强度(x)的响应规律均符合方程y=Pm×tanh(α×x÷Pm)+Rd(图1),但光合固碳速率存在较大差异,表现为海带>萱藻>鼠尾藻>珊瑚藻>铜藻>浒苔>瓦氏马尾藻>蜈蚣藻>石莼>铁钉菜>羊栖菜。其中,海带的固碳速率最大,固碳量可达0.89×10-3t·t-1·h-1,羊栖菜的固碳速率最小,固碳量为0.28×10-3t·t-1·h-1,约是前者的31.5%。其他海藻固碳能力居两者之间。

图1 大型海藻光合固碳能力与光的关系
2.2 大型海藻间光合固碳能力的比较
2种绿藻对比,浒苔的固碳能力较高,为29.96 μmol·g-1·h-1,石莼的固碳能力较低,为28.93 μmol·g-1·h-1。2种红藻对比,珊瑚藻的固碳能力较高,为34.71 μmol·g-1·h-1,蜈蚣藻固碳能力较低,为23.76 μmol·g-1·h-1。褐藻之间相比较,海带的固碳能力最高,达70.58 μmol·g-1·h-1,萱藻、鼠尾藻、铜藻、瓦氏马尾藻、铁钉菜、羊栖菜6种藻依次降低,羊栖菜的固碳能力最低,为20.03 μmol·g-1·h-1,仅是海带的28.4%。比较分析可知,11种大型海藻的固碳能力存在一定差异。大型海藻无机碳的利用能力与其所在生态位相关, 一般认为,3 种基本大型海藻无机碳利用能力由高到低依次为绿藻、褐藻、红藻[11]。这与海藻的地理分布区域也有一定的关系,分布于潮间带的海藻对强光的适应能力略高于潮下带海藻。在本研究中发现,在潮间带的3种大型海藻石莼、浒苔和铁钉菜中,浒苔的饱和光强最大,为67.23 μmol·m-2·s-1,明显高于潮下带的蜈蚣藻和铜藻。从光能利用率α的角度分析,潮间带中石莼的光能利用率α最大,并且高于潮下带的大型海藻。这说明长期的生态环境适应促使潮间带的大型海藻对低光的耐受性和利用能力普遍高于潮下带的大型海藻(表1)。

表1 大型海藻Pm、α及Ik的比较
绿藻、红藻中固碳能力最高的是浒苔和珊瑚藻,但也仅是褐藻中海带固碳能力的52%和56%。由此可见,大规模开展养殖经济海藻海带,对于增加渔业碳汇能力是至关重要的。从生态系统多样性的角度来说,其他藻类固碳能力虽低于海带,但也要对其进行合理的保护,以保证大型海藻的物种多样性,维持生态系统平衡。
2.3 大型海藻可减排的最大CO2量和可创造的经济价值
大型海藻单位鲜重(t)、单位时间内(h)减排的最大CO2量如表2所示。其中,大型海藻可创造的经济价值依据《联合国气候变化框架公约的京都议定书》确定的工业化国家减排CO2的开支标准150~600美元·t-1·h-1进行估算。
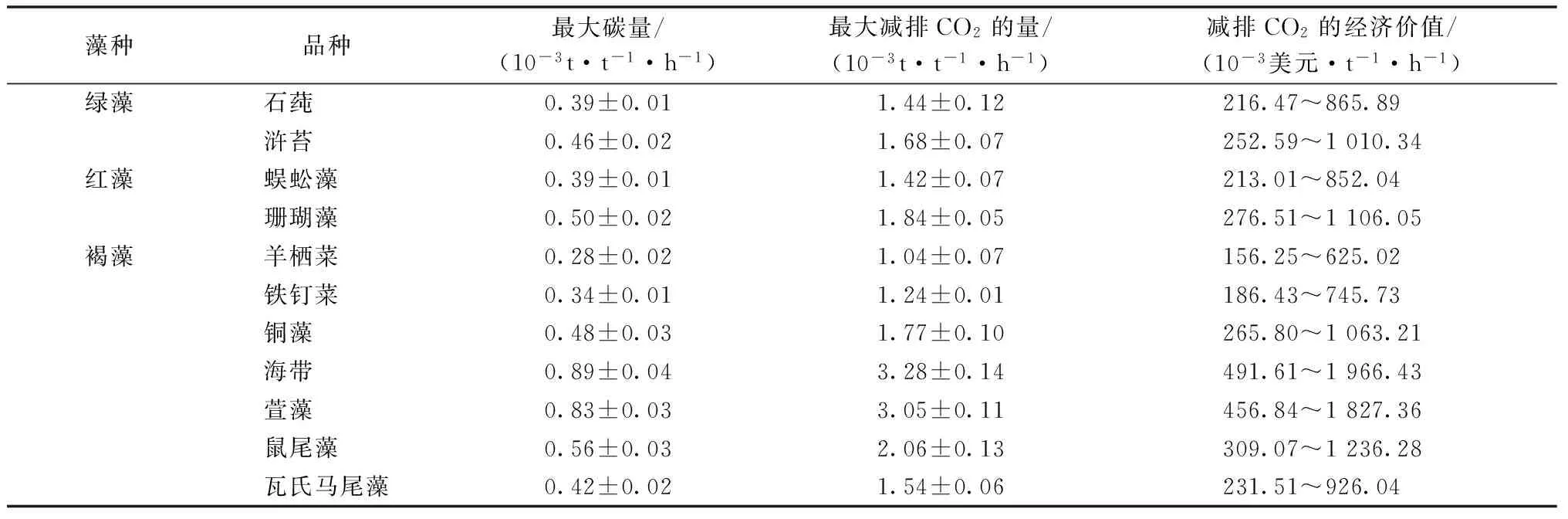
表2 大型海藻可达到的最大固碳能力、减排的最大CO2量和减排CO2的潜在经济价值
在本研究中,发现供试11种海藻的固碳能力存在显著的种间差异,固碳能力最高的海带可减排的最大CO2为3.28×10-3t·t-1·h-1,该减排量可创造的经济价值为0.49~1.97美元·t-1·h-1。固碳能力最小的羊栖菜减排的最大CO2仅为1.04×10-3t·t-1·h-1,其可创造的经济价值仅为0.16~0.64美元·t-1·h-1,其他9种大型海藻的固碳能力居两者之中。
同时可依据当地大型海藻的海产养殖面积和产量计算出大型海藻减排CO2的潜在总经济价值。为当地渔业估算大型海藻养殖在减排CO2所能创造的经济价值方面提供理论支撑。
3 讨论
光合作用作为大型海藻基础而复杂的生理代谢过程, 光强是影响光合固碳效率的直接因素。大型海藻作为海洋碳汇重要的组成部分,其养殖对于扩增海洋碳汇效应具有重要意义, 其固碳量和碳汇潜力都非常惊人。已有研究发现, 长石莼(U.linza)在25 ℃时的饱和光强为72 μmol·m-2·s-1, 在光饱和点以下,增加光强可促进光合作用, 在光抑制之后, 光合固碳能力随之下降[12]。海洋植物的总量虽然只有陆生植物总量的0.05%,但是其稳定碳总量却与陆生植物不相上下, 主要是因为海洋特殊的环境使海洋植物捕集转化并储存碳的能力和效率增加[13]。Duarte 等[14]的研究表明,海洋大型植物每年为全球埋藏的碳量高达120~329 Mt。而Chung 等[15]对于多年生褐藻Ecklonia试点海藻场研究结果表明, 该试点藻场利用中层水缆绳养殖模式每年可沉降CO2近10 t。由此可见, 培养和增殖大型藻类对扩增海洋碳汇具有重要的意义和价值。
本研究采用氧电极法针对大型海藻的碳汇能力进行检测。经测定,大型海藻的碳汇能力受到光照强度的显著影响,其固碳能力在一定范围内随着光照强度的增加而增加,而当光照强度增加到其光饱点时,固碳能力不再增加。大型海藻生活在潮间带和潮下带,随着潮水涨退,其接受的光也存在高低的波动,因此,大型海藻的固碳能力受到阳光日辐射强度变化和潮水涨退的影响。研究检测的11种大型海藻都具有显著的固碳能力,对于减排CO2都具有重要作用,其中海带碳汇能力最强,可创造的经济价值最大。因此,海带的大规模养殖对于渔业碳汇和生态保护具有十分重要的作用,不仅仅可以直接提供产品,还增加了碳汇,创造了巨大的社会和经济效益。造成碳汇能力差异的主要原因与藻体本身的差异以及其生长的环境有一定的关系,虽然其他大型海藻的碳汇能力小于海带,但由于其生物量巨大,分布较广,它们对渔业碳汇的贡献也十分重要。
生物固碳具有操作成本低、易施行的特点,且可以达到间接减排的效果,开展碳汇渔业的研究,关注大型海藻对渔业碳汇的贡献,在满足应对全球气候变暖、大力发展低碳经济和节能减排的同时,还有利于保护生态系统、保护大型海藻物种多样性和净化近岸水体污染。这有利于改善和修复在经济快速发展过程中给近岸水域环境造成的污染和破坏。
4 小结
随着光照强度的增加,11种大型海藻光合固碳能力都逐渐增大,且当光照强度增加到一定值时趋向饱和,达到最大固碳能力。大型海藻的碳汇能力存在一定的种间差异,但由于大型海藻生物量巨大,分布较广,不同的大型海藻对渔业碳汇的贡献都是不可忽视的。因此,意识到大型海藻对于渔业碳汇能力的贡献,能够合理地开发和利用大型海藻资源,对于生态修复和生态文明建设是至关重要的。
[1] SABINE C L, FEELY R A, GRUDER N, et al. The oceanic sink for anthropogenic CO2[J]. Science, 2004, 305(5682):367-371.
[2] IPCC. Climate change2007:Synthesis report[R].Geneva: IPCC, 2007.
[3] SOLOMON S, QIND, MANNING M, et al. IPCC, 2007: Climate change 2007: The physical science basis. Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change[M]. Cambridge, U K: Cambridge University Press, 2007: 996.
[4] STOCKER T F, QIN D, PLANTTNER G K, et al. IPCC, 2013: Climate change 2013: The physical science basis. Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change[M]. Cambridge, U K and New York: Cambridge University Press, 2013: 525-527.
[5] KHATIWALA S, PRIMEAU F, HALL T. Reconstruction of the history of anthropogenic CO2concentrations in the ocean [J]. Nature, 2009, 462: 346-349.
[6] LAFFOLEY D, GRIMSDITCH G. The management of natural coastal carbon sinks [M]. IUCN: Gland,2009:53.
[7] HOOD M, BROADGATE W, URBAN E, et al. Ocean acidification-a summary for policymakers from the second symposium on the ocean in high-CO2world [C]. IOC, 2009.
[8] COSTANZA R, ARGE R, GROOT R d, et al.The value of the world’s ecosystem services and natural capital[J].Nature, 1977, 387:253-260.
[9] 陈仲新, 张新时. 中国生态系统效益的价值[J]. 科学通报, 2000, 45(1): 17-23.
[10] HENLEY W J.Measurement and interpretation of photosynthetic light-response curves in algae in the context of photo inhibition and diel changes[J]. Journal of Phycology, 1993, 29(6): 729-739.
[11] 韩博平, 韩志国, 付翔. 藻类光合作用机理与模型[M]. 北京: 科学出版社, 2003: 48-53.
[12] 汤文仲, 李信书, 黄海燕, 等.不同光强和温度对长石莼(缘管浒苔)光合作用和叶绿素荧光参数的影响[J]. 水产学报, 2009, 33: 762-769.
[13] 刘慧, 唐启升. 国际海洋生物碳汇研究进展[J]. 中国水产科学, 2011, 18(3): 695-702.
[14] DUARTE C M, CHISCANO C L. Seagrass biomass and production: A reassessment[J]. Aquat Bot, 1999, 65(1): 159-174.
[15] CHUNG I K, JUNG H O. Installing kelp forests/seaweed beds for mitigation and adaptation against global warming: Korean project overview[J]. Ices Journal of Marine Science, 2013, 70(6):1038-1044.
(责任编辑:吴益伟)
2017-05-12
浙江省科技计划项目(2016F50032)
欧官用(1996—),男,贵州遵义人,本科生,E-mail: ouguanyong1214@163.com。
柯爱英(1965—),女,浙江建德人,高级工程师,从事海水养殖研究工作,E-mail: keaiying8899@126.com。
10.16178/j.issn.0528-9017.20170843
Q949.2
A
0528-9017(2017)08-1436-04
文献著录格式:欧官用,王鑫杰,杨安强,等. 大型海藻碳汇能力的种间差异[J].浙江农业科学,2017,58(8):1436-1439,1443.
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
中国农业科学(2022年10期)2022-06-28
广东蚕业(2021年12期)2022-01-18
四川蚕业(2021年4期)2021-12-01
山西林业(2021年2期)2021-07-21
第一财经(2020年10期)2020-10-15
第一财经(2020年3期)2020-02-29
第一财经(2019年11期)2019-12-02
第一财经(2019年7期)2019-07-25
蔬菜(2018年5期)2018-05-17
