
粉棒束孢对柑橘全爪螨的毒杀活性及其生物学特性
2017-07-18 11:41龙艳玲胡军华王帆陈若霓陈仕江卿玉玲秦绍蓉
湖南农业大学学报(自然科学版) 2017年4期
龙艳玲,胡军华*,王帆,陈若霓,陈仕江,卿玉玲,秦绍蓉
(1.西南大学柑橘研究所,重庆 400712;2.太极集团有限公司,重庆 400712;3.重庆市中药研究院甘孜州康定贡嘎中华虫草产业有限责任公司,重庆 400712)
粉棒束孢对柑橘全爪螨的毒杀活性及其生物学特性
龙艳玲1,胡军华1*,王帆1,陈若霓2,陈仕江3,卿玉玲2,秦绍蓉2
(1.西南大学柑橘研究所,重庆 400712;2.太极集团有限公司,重庆 400712;3.重庆市中药研究院甘孜州康定贡嘎中华虫草产业有限责任公司,重庆 400712)
室内测定了粉棒束孢(Paecilomyces farinosus)菌株nq1-1对柑橘全爪螨雌成螨的毒力,并通过单因素试验,分析该菌的生物学特性。结果表明:nq1-1菌株对柑橘全爪螨雌成螨的LC50和LT50分别为1.59×107/mL和7.9 d;菌株最适生长和产孢培养基分别为萨氏培养基和PDA培养基;蔗糖和有机氮源(赖氨酸)有利于该菌生长和产孢;菌丝生长和产孢的最适温度均为20 ℃,最适孢子萌发温度为25~28 ℃;菌丝生长、产孢以及孢子萌发的最适pH均为6.0;nq1-1菌株具有较强的抗紫外线能力;82.6%氧化亚铜(水分散粒剂)、75%百菌清(可湿性粉剂)、50%喹啉铜(可湿性粉剂)等杀菌剂与菌株相容性较好。nq1-1菌株对柑橘全爪螨有较好的毒杀活性,喜好偏低温、中性以及富含有机营养的环境。
粉棒束孢;柑橘全爪螨;杀螨活性;生物学特性
以柑橘全爪螨(Panonychus citri(McGregor))为代表的果园害虫,个体小,发育历期短,种群密度大,繁殖能力和适应力强,对这类害虫的防治大多依赖化学农药,抗药性问题十分突出[1-2]。玫烟色拟青霉(Paecilomyces fumosorosea)曾被报道对朱砂叶螨雌成螨及其螨卵有较强的侵染致病力,在39~1 554 /mm2下杀螨19.4%~77.7%[3];高毒力环链拟青霉(Paecilomyces cateniannulata)芽孢对二斑叶螨雌成螨LT50仅为1.472 d[4];淡紫拟青霉(Paecilomyces lilacinus)、宛氏拟青霉(Paecilomycesvariotii)也曾被报道对蚜虫、粉虱、木虱等小型刺吸式口器害虫有很强的致病力[5-8]。粉棒束孢(Isaria farinose(Homskiold)),又名粉拟青霉(Paecilomyces farinosus),杀虫毒力强,寄主范围广,对油松毛虫、松梢螟、苹果蠹蛾等害虫有良好的杀虫效果[9-14],被认为是极具应用潜力的生防菌种,但其对柑橘全爪螨是否具有毒杀效果尚未见相关报道。
笔者从贡嘎蝙蝠蛾发病幼虫上分离得到1株粉棒束孢菌株 nq1-1,采用室内毒力法测定了其对柑橘全爪螨雌成螨的毒力,并分析该菌的生物学特性,以期为生物防控叶螨提供参考。
1 材料与方法
1.1 材料
供试菌株贡嘎蝙蝠蛾幼虫粉棒束孢 nq1-1菌株,分离自四川康定患病贡嘎蝙蝠蛾幼虫,经纯化、鉴定后,保藏于西南大学柑橘研究所。在 PDA培养基上(25±1) ℃、相对湿度95%培养10 d(每天12 h光照,12 h黑暗)。
4龄家蚕由西南大学蚕学与系统生物研究所提供。用新鲜桑叶饲养,温度25 ℃、相对湿度95%,每天8 h光照,16 h黑暗。
于西南大学柑橘研究所采集带柑橘全爪螨的枝条和叶片,温室培养,温度 25 ℃、相对湿度60%~80%,每天8 h光照、16 h黑暗。
1.2 方法
1.2.1 nq1-1菌株对4龄家蚕和柑橘全爪螨的毒力测定
用含0.02% Tween-80灭菌水洗下培养10 d的nq1-1菌株分生孢子,过滤后充分振荡混匀,分别配制成 1×108、1×107、1×106、1×105、1×104、1×103/mL的悬浮液。
分别取4龄家蚕20头,将其浸入孢子悬浮液中20 s后取出,用滤纸吸干,放入烧杯中,加入新鲜桑叶饲养,以含 0.02%吐温-80的灭菌水处理家蚕和桑叶作对照,第2天开始,换用新鲜桑叶常规饲养。每个处理 3次重复,以镊子接触虫体,5 s内不动判定为死亡。连续11 d逐日观察,统计幼虫死亡率。
参照李一玉[15]方法,喷雾接种柑橘全爪螨雌成螨:采集新鲜健康的温州蜜柑叶片,75%乙醇清洗后,无菌水清洗3遍,自然晾干,置于铺有湿海绵和滤纸的托盘内,叶缘用湿棉条围绕保湿。每片温州密柑叶片上接35雌成螨,均匀喷洒nq1-1菌株分生孢子悬液2.5 mL,3次重复,对照组喷洒2.5 mL含 0.02%吐温-80的灭菌水,自然晾干后,用保鲜膜封口培养,连续7 d逐日统计柑橘全爪螨雌成螨死亡率(对照组统计时间直至螨全部死亡)。挑取死螨于干净培养皿中保湿培养,螨尸上长出nq1-1菌丝的为有效处理。
1.2.2 nq1-1菌株生物学特性的测定
1) 培养基种类及碳源和氮源的筛选。配制马铃薯葡萄糖琼脂培养基(PDA)、萨氏培养基(SDAY)、察氏培养基(Czapek)、马铃薯蔗糖琼脂培养基(PSA)、燕麦培养基(OA)、玉米粉培养基(CMA)、综合马铃薯培养基(SPA)、米饭蛋白胨培养基(RPA)。
分别以 3%蔗糖、甘油、乳糖、麦芽糖、甘露醇、D-果糖、可溶性淀粉代替察氏培养基(Czapek)中的葡萄糖,配制成7种不同碳源培养基。分别以0.3%的硝酸铵、硝酸钾、蛋白胨、甘氨酸、赖氨酸、氯化铵、尿素代替察氏培养基(Czapek)中的硝酸钠,配制成7种不同氮源培养基。
接种2 µL nq1-1孢子悬浮液,每处理5次重复。第 10天时采用十字交叉法测量菌落直径,计算菌株平均日增长量,并每皿加5 mL含0.02%吐温-80的无菌水洗脱分生孢子,用血球计数板在显微镜下检查分生孢子数量。
2) 培养温度、酸碱度和紫外线照射时间的筛选。接种2µL nq1-1孢子悬浮液(1×107/mL)于PDA平板中央,分别置于4、10、15、20、25、28、30、35、40 ℃培养;将PDA培养基的pH用1 mol/L HCl和1 mol/L NaOH调节为5.0、6.0、7.0、8.0、9.0、10.0,接种 2 µL nq1-1 孢子悬浮液(1×107/mL)后培养;接种2 µL孢子悬浮液(1×107/mL)于PDA平板中央,紫外线(40 W,距离45 cm)分别照射0、5、10、20、30、40、50、60 min后培养。每处理5次重复,第8天时测定菌落直径和产孢量。
取nq1-1孢子悬浮液(1×107/mL)500 µL于1.5 mL离心管中,分别置于4、10、15、20、25、28、30、35、40 ℃暗培养;配制pH分别为5.0、6.0、7.0、8.0、9.0、10.0 的 nq1-1 孢子悬浮液(1×107/mL),取各pH孢子悬浮液500 µL于1.5 mL离心管中,25 ℃暗培养;将配制好的孢子悬浮液(1×107/mL)置于培养皿中,揭开上盖,紫外线(40 W,距离45 cm)分别照射0、5、10、20、30、40、50、60 min后,取500 µL于1.5 mL离心管中25 ℃暗培养。24 h时镜检孢子萌发情况,每处理重复3次,每次检查3个视野孢子数,计算萌发率。
1.2.3 nq1-1菌株与杀菌剂的相容性筛选
选取22种田间常用杀菌剂(表1),根据各杀菌剂田间浓度配制方法用无菌水将其分别配制成不同浓度,并分别取1 mL(无菌水作为对照)与99 mL PDA培养基混合均匀,制成不同浓度的培养平板。将已在PDA培养基培养10 d的nq1-1菌株用直径为5 mm的打孔器在菌落边沿打取菌丝块,接入上述含药PDA培养基中,每处理5次重复,第10天测定菌落直径,计算各杀菌剂对nq1-1菌株菌丝生长的抑制率。

表1 22种杀菌剂的使用浓度Table 1 The working concentration of 22 different antiseptics
1.3 数据处理
采用农药室内生物测定数据处理系统(PBT),计算柑橘全爪螨的累积死亡率、校正死亡率、半致死浓度LC50值和半致死时间LT50值。用SPSS 20统计软件对数据进行差异显著性分析。
2 结果与分析
2.1 nq1-1菌株的杀螨和杀虫活性
随着nq1-1菌株孢子悬液浓度的增加,4龄家蚕和柑橘全爪螨雌成螨的累积死亡率都升高。当nq1-1菌株孢子浓度为1×108/mL时,家蚕4龄幼虫第11天的校正死亡率为77%,柑橘全爪螨雌成螨第7天的校正死亡率为44%。数据分析表明,4龄家蚕的 LC50为 1.59×107/mL,毒力回归方程为Y=2.310 7+0.373 4X(χ2=1.64),1×108/mL 处理 4 龄家蚕的LT50为7.9 d,毒力回归方程为Y=1.134 1+4.300 4X(χ2=3.98);柑橘全爪螨在处理第 7天时 LC50为7.51×108/mL,毒力回归方程为Y=2.935 9+ 0.232 6X(χ2=6.48),1×108/mL 处理柑橘全爪螨的 LT50为 8.0 d(对照柑橘全爪螨雌成螨平均寿命为11.7 d),毒力回归方程为Y=2.466 2+2.812 3X (χ2=4.24)。粉棒束孢nq1-1菌株对4龄家蚕的杀虫毒力较对柑橘全爪螨的强。
2.2 nq1-1菌株的生物学特性
2.2.1 培养基种类、碳源和氮源对 nq1-1菌株生长和产孢的影响
从表2可知,不同培养基上nq1-1菌株菌丝的生长速率及产孢量不同。nq1-1菌株在SDAY上生长最快,SDAY培养基为最适生长培养基,其次是Czapek、RPA和PSA培养基,在CMA培养基上生长最慢。nq1-1产孢量最高的培养基是PDA培养基,其次是SDAY培养基,在CMA培养基上产孢量低,方差分析显示其为最差产孢培养基。
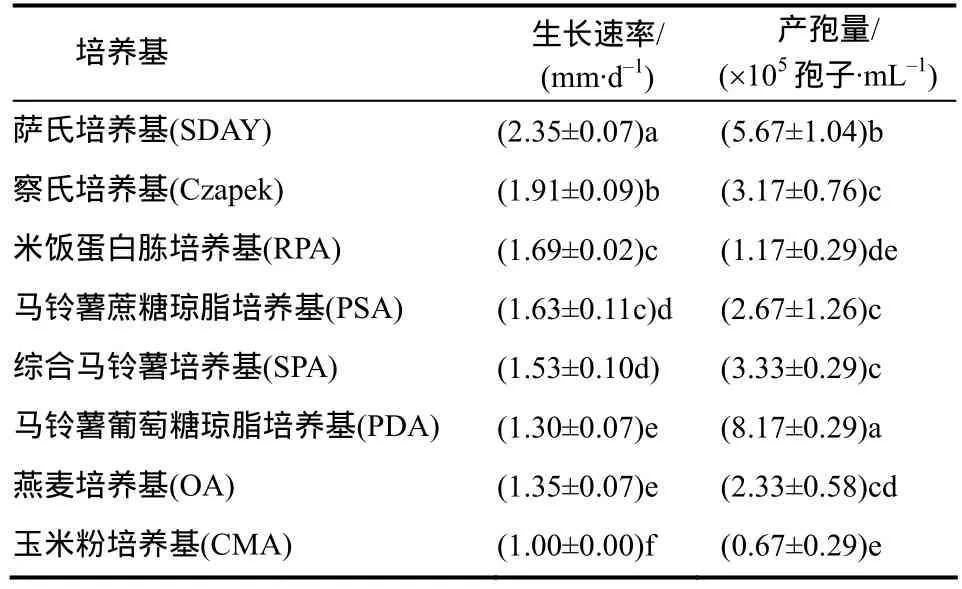
表2 nq1-1菌株在不同培养基上的生长速率和产孢量Table 2 Growth rate and spore production of strain nq1-1 on different media
从表3可知,nq1-1菌株在8种不同碳源和氮源培养基上均能正常生长和产孢。以葡萄糖为碳源时,最利于菌落生长,方差分析显示其为最适生长碳源,其次是蔗糖和甘油,在D-果糖和可溶性淀粉培养基上菌落生长最慢。以蔗糖、甘油、葡萄糖为碳源时,菌株产孢量与其他碳源产孢量差异显著,其次为乳糖和D-果糖,在淀粉为碳源的培养基上产孢量最低,方差分析显示其为产孢最差碳源。
以硝酸钠、尿素、硝酸铵、赖氨酸、蛋白胨 5者为氮源时,菌株生长速率和产孢量的差异不显著,均为适合生长氮源。在供试的8种氮源中,产孢量较高的是添加了甘氨酸、赖氨酸的培养基,二者与其他氮源产孢量差异显著;其次是蛋白胨和硝酸钠,而尿素为氮源时产孢量最低,方差分析显示其为产孢最差氮源。
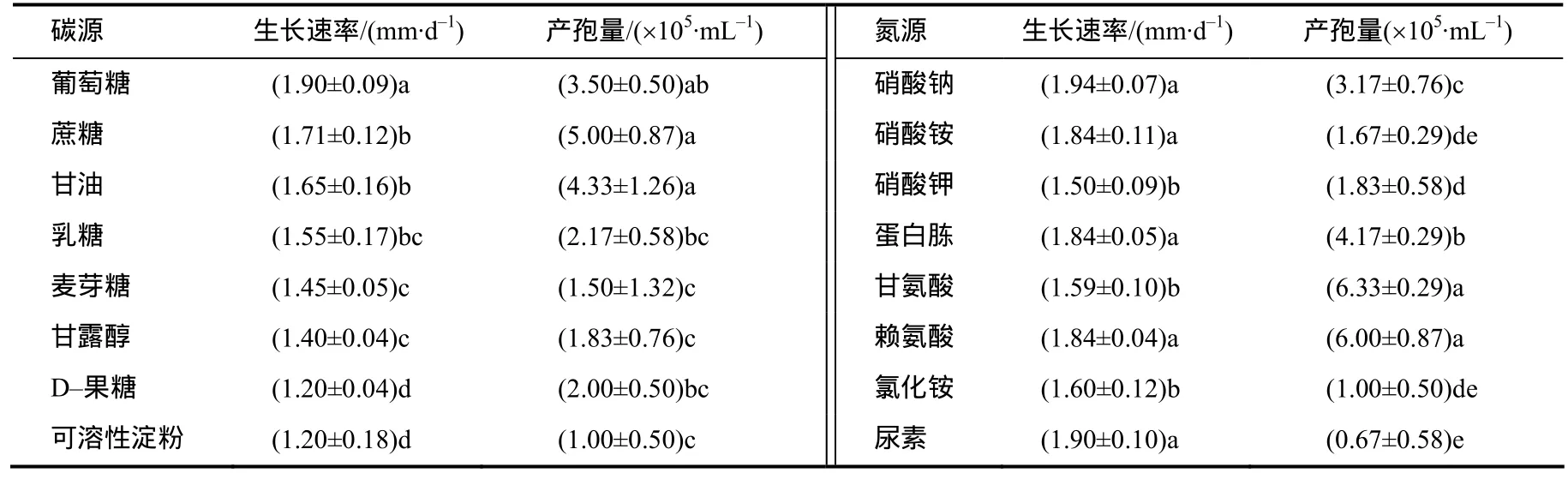
表3 不同碳源和氮源下nq1-1菌株的生长速率和产孢量Table 3 Growth rate and spore production of strain nq1-1 on different carbon sources and nitrogen sources
2.2.2 温度、pH和紫外线照射时间对nq1-1菌株生长、产孢和孢子萌发的影响
温度对 nq1-1菌株菌丝生长、产孢和孢子萌发有明显的影响(表4)。nq1-1菌株除在35 ℃和40 ℃不能正常生长和产孢外,在4~30 ℃均能生长、产孢。菌丝生长速率和产孢量在4~20 ℃逐渐上升,方差分析显示最适生长、产孢温度为 20 ℃;当温度超过20 ℃后,菌丝生长速率和产孢量逐渐下降;孢子在 4~28 ℃萌发率逐渐上升,方差分析显示25~28 ℃为最适萌发温度。
pH对菌丝生长、产孢和孢子萌发有明显影响,nq1-1菌株在pH为5.0~10.0均能生长、产孢和萌发(表5)。pH为6.0时,菌株的生长速率、产孢量和孢子萌发率最大,方差分析显示其为最适pH;当pH大于6.0后,该菌的生长速率、产孢量以及孢子萌发率逐渐降低。
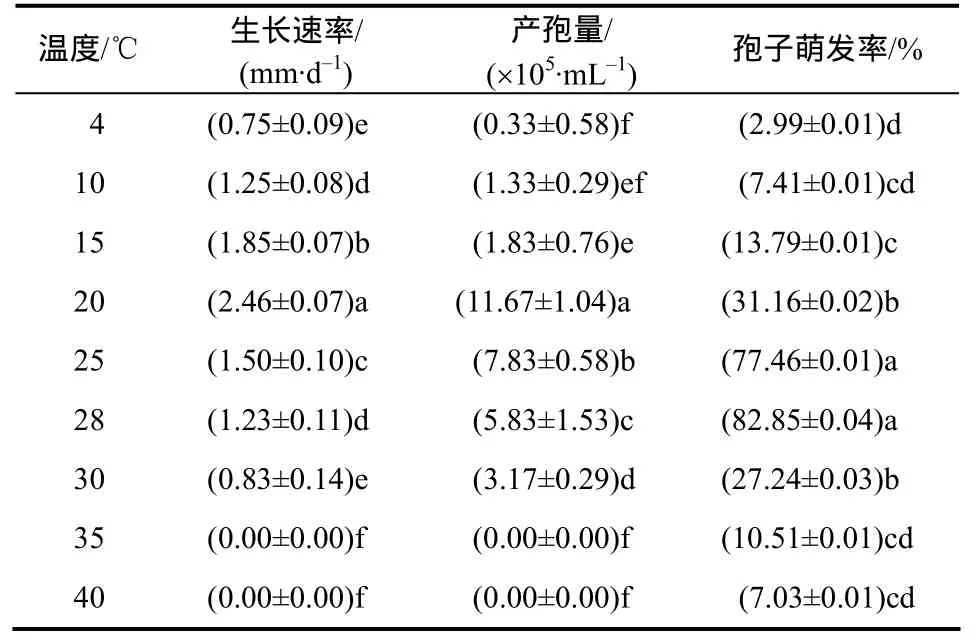
表4 nq1-1菌株在不同温度下的生长速率和产孢量及孢子萌发率Table 4 Growth rate, spore production and spore germination of strain nq1-1 at different temperatures
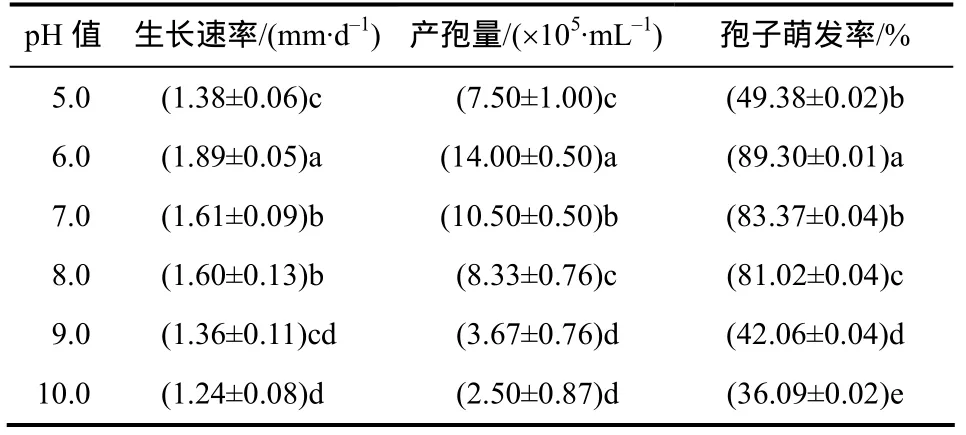
表5 nq1-1菌株在不同pH下的生长速率和产孢量及孢子萌发率Table 5 Growth rate, spore production and spore germination of strain nq1-1 on different pH
紫外线照射时间对nq1-1菌株的生长、产孢和孢子萌发有一定的抑制作用,且随着照射时间的延长,抑制作用增强,菌株生长速率、产孢量及孢子萌发率逐渐降低。菌株经紫外线照射30 min,其生长速率比对照降低15.85%,产孢量降低10.28%,孢子萌发率降低 24.79%;当紫外线照射时间至 60 min时,其生长速率比对照降低 29.27%,产孢量降低18.35%,孢子萌发率降低50.76%,紫外线照射对nq1-1菌株孢子具有较强的杀伤作用。
2.2.3 杀菌剂对nq1-1菌株生长的影响
22种杀菌剂对 nq1-1菌株菌丝生长的影响因杀菌剂的种类和浓度的不同而呈现差异(表7)。25%吡唑醚菌酯EC、25%嘧菌酯SC、25%咪鲜胺EC、70%丙森锌 WP、50%异菌脲 SC、12.5%烯唑醇 WP、50%多菌灵WP、60%唑醚·代森联WG、40%氟硅唑EC、40%腈菌唑WP、80%代森锌WP、45%敌磺钠WP 12种杀菌剂对菌株生长有抑制作用,其中以25%吡唑醚菌酯EC和40%腈菌唑WP的抑制作用最明显,抑制率均为100.00%;其次为25%咪鲜胺EC(90.53%)、50%异菌脲SC(82.11%);82.6%氧化亚铜WG、75%百菌清WP、50%喹啉铜WP、43%戊唑醇SC等10种杀菌剂与粉棒束孢nq1-1相容性较好。
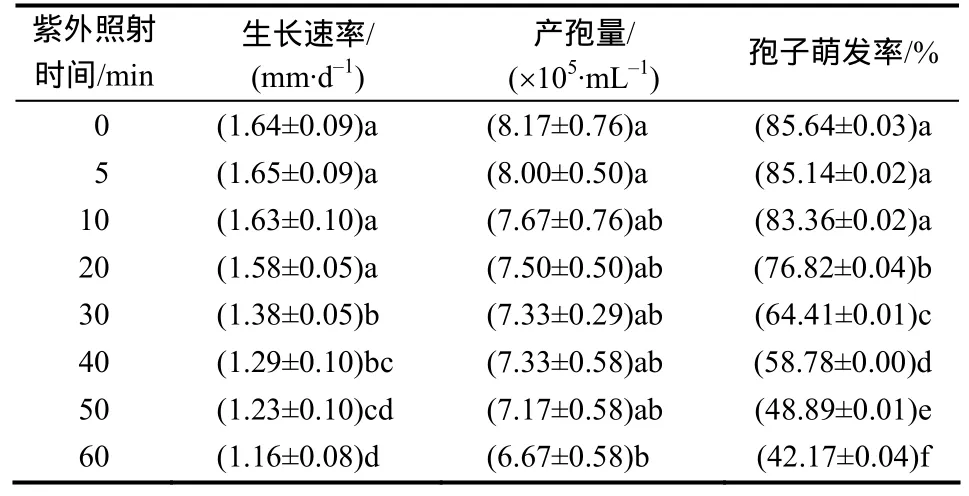
表6 nq1-1菌株在不同紫外线照射时间下的生长速率和产孢量及孢子萌发率Table 6 Growth r ate, spor e pr oduction and s pore ge rmination of strain nq1-1 under different UV irradiation
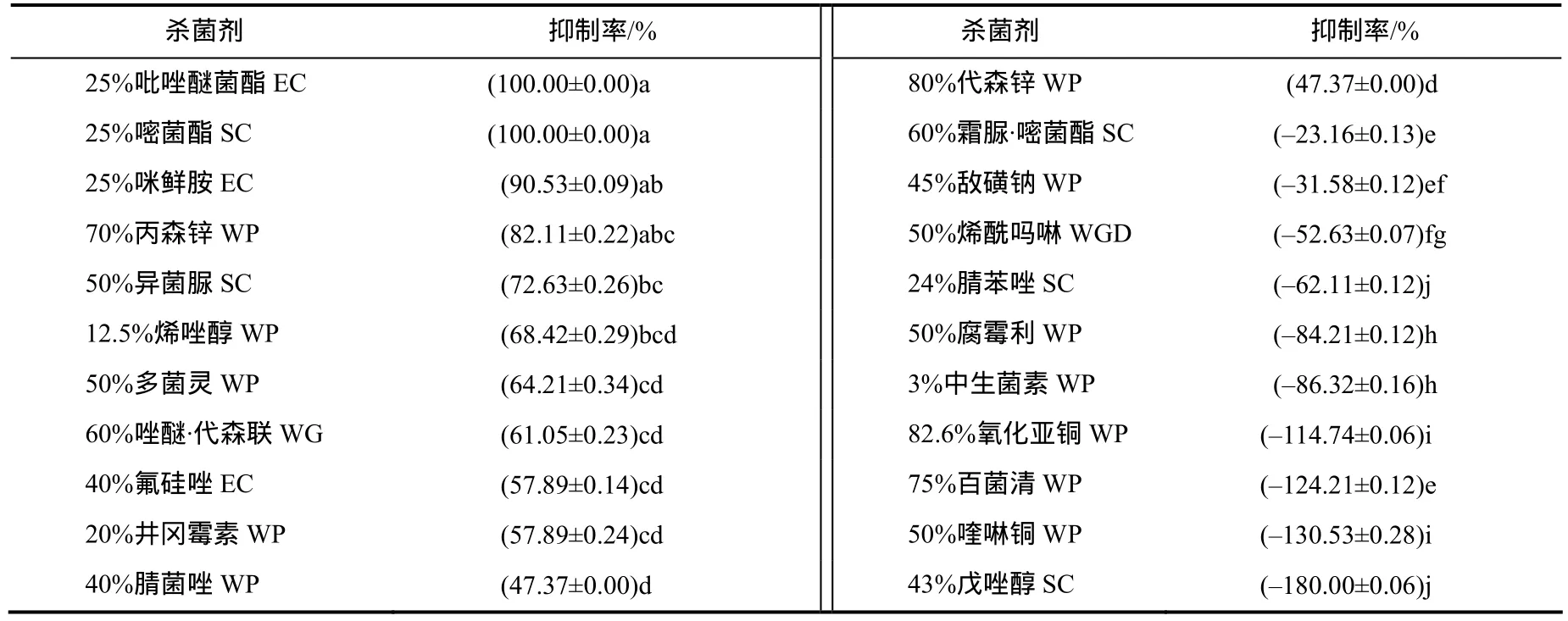
表7 22种杀菌剂对nq1-1菌株的生长抑制率Table 7 Growth inhibition ratio for strain nq1-1 with 22 antiseptics
3 讨论
施卫兵[3]曾报道 2株玫烟色拟青霉对螨卵的LC50分别为848和913/mm2;李丰伯等[16]研究发现,环链拟青霉孢子浓度为2.0×107~11.0×107/mL时,细纹新须螨幼虫死亡率44.4%~76.3%,LC50为6.196 7×107/mL;安建梅[17]报道粉拟青霉在孢子浓度为5×108/mL时,黄刺蛾 3至 4龄幼虫的死亡率为73.3%;韩燕峰等[18]发现在孢子浓度为 1×107/mL时,环链拟青霉和玫烟色拟青霉对小菜蛾2~3龄幼虫的致病死亡率达到 100%,说明玫烟色拟青霉和环链拟青霉对叶螨和鳞翅目昆虫具有很强的致病性。笔者分离得到的粉棒束孢(粉拟青霉)对柑橘全爪螨和家蚕均表现致病性,且菌株对家蚕4龄幼虫的侵染致死效果优于对柑橘全爪螨,但均逊于玫烟色拟青霉和环链拟青霉,菌株、寄主种类和地理来源的不同可能与病原真菌对昆虫的致病力相关,但究其根本还是因为菌株本身的遗传特性。后续拟采用于世将[19]构建的遗传转化体系对粉棒束孢nq1-1菌株进行毒力改造,筛选对柑橘全爪螨或其他螨类高毒力的转化子,并与其他杀螨真菌的毒力进行比较,扩大粉棒束孢的杀虫谱。
成本低廉的多糖形式碳源(蔗糖)和有机氮源(赖氨酸)以及中性偏微酸条件下更有利于nq1-1的生长和产孢,这与申慧荣[20]、安建梅[21]以及曾伟等[22]研究结果基本一致。张伟等[23]报道玫烟色棒束孢经紫外线照射1 min后,孢子萌发率为74.48%,照射时间延长至6 min时,孢子萌发率仅有4.16%,而粉棒束孢nq1-1菌株分生孢子经紫外线照射60 min后萌发率仍达42.17%,具有较强的抗紫外线能力,说明粉棒束孢nq1-1菌株具有较强的环境适应性,在田间施用具有优势。笔者发现 nq1-1菌株较耐低温而不耐高温。曾伟等[1,22]也曾报道粉拟青霉(粉棒束孢)是一种适宜低温生长的真菌,最适生长温度为20~25 ℃,并在20 ℃时对蝙蝠蛾幼虫作用最快,致病力也最强,14 d全部死亡;安建梅[17]也发现粉拟青霉(粉棒束孢)在20~25 ℃时对黄刺蛾幼虫的致死率最高,当温度继续升高时死亡率反而下降。柑橘全爪螨各螨态发育历期与温度有着密切的关系,20~30 ℃宜其发育和繁殖,与粉棒束孢(粉拟青霉)最适生长、产孢温度相符,在害螨暴发高峰期前合理施用,有效控制害虫大发生,可起到事半功倍的效果;而在防治家蚕、冬虫夏草寄主昆虫粉棒束孢病害时,可以在保证幼虫正常生长的基础上适当降低培养室内温度延缓粉棒束孢的生长、产孢和孢子萌发,或者利用高温处理饲养工具和基质减少侵染源。
25%吡唑醚菌酯EC、25%嘧菌酯SC、25%咪鲜胺EC、70%丙森锌WP、50%异菌脲SC、12.5%烯唑醇WP、50%多菌灵WP 7种杀菌剂在常规浓度下对粉棒束孢nq1-1菌株生长具有较强的抑制作用,对粉棒束孢(粉拟青霉)在大田的应用会产生不良影响,在使用时应避免这 7种杀菌剂同时或者混合使用。82.6%氧化亚铜WG对菌株nq1-1无明显作用,可以考虑将这些杀菌剂与粉棒束孢(粉拟青霉)制成混合制剂进一步测试其田间效果。
[1] 孟和生,王开运,姜兴印,等.柑橘全爪螨对哒螨灵抗性的选育及其生化机理[J].农药学学报,2000,2(3):30-34.
[2] DAGLI F,TUNC I.Dicofol resistance in Tetranychus cinnabarinus:resistance and stability of resistance in populations from Antalya,Turkey[J].Pest Management Science,2001,57:609-614.
[3] 施卫兵.生防真菌对叶螨类植物害螨的微生物防治研究[D].杭州:浙江大学,2007.
[4] 张晓娜,金道超,邹晓,等.杀二斑叶螨高毒力环链棒束孢菌株的筛选及其对尼氏真绥螨的影响[J].环境昆虫学报,2014,36 (3):372-380.
[5] 黄振,任顺祥,吴建辉.玫烟色拟青霉和吡虫啉对烟粉虱种群的联合控制作用[J].生态学报,2006,26(10):3250-3257.
[6] ZIMMERMANN G.The entomopathogenic fungi Isaria farinose(formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex(formerly Paecilomyces fumosoroseus):biology,ecology and use in biological control[J].Biocontrol Science and Technology,2008,18(9):865-901.
[7] 宋晓兵,彭埃天,程保平,等.利用虫生真菌生物防治柑橘木虱的研究进展[J].生物安全学报,2016,25(4):255-260.
[8] DEMIRCI F,MUŞTU M,KAYDAN M B,et al.Laboratory evaluation of the effectiveness of the entormopathogen:Isaria farinose,on citrus mealybug,Planococcuscitri[J].Journal of Pest Science,2011,84(3):337-342.
[9] 赵瑞兴,武觐文.粉拟青霉防治油(赤)松毛虫流行病研究[J].林业科学,1991,27(3):219-228.
[10] 武觐文.应用粉拟青霉防治松毛虫[J].安徽农业大学学报,1996,23(3):438-445.
[11] PERRY Polar,MOSES T K,Kairo,et al.Assessment of fungal isolates for development of a Myco-Acaricide for cattle tick control[J].Vector Borne and Zoonotic Diseases,2005,5(3):276-284.
[12] 马良进,张立钦,林海萍,等.松墨天牛病原菌及其致病性研究[J].中国生物防治,2009,25(3):220-224.
[13] 覃丰全,闫国增,贺伟,等.用球孢白僵菌和粉拟青霉防治美国白蛾的研究[J].山西农业科学,2012,40(2):140-145.
[14] 张德利,曾纬,陈仕江,等.蝙蝠蛾幼虫拟青霉病菌的分离与鉴定及致病力研究[J].广东农业科学,2011,38(17):57-58.
[15] 李一玉.球孢白僵菌Bb025对朱砂叶螨实验种群的控制作用研究[D].重庆:西南大学,2014.
[16] 李丰伯,汪传友,姚剑飞,等.环链拟青霉防治黄山风景区细纹新须螨[J].东北林业大学学报,2011,39(7):77-78.
[17] 安建梅.虫生真菌粉质拟青霉对黄刺蛾幼虫的致病性研究[J].山西农业大学学报,2000,20(2):105-107.
[18] 韩燕峰,张延威,梁建东,等.一些拟青霉菌株对小菜蛾致病性的初步研究[J].植物保护,2008,34(2):46-49.
[19] 于世将.渐狭蜡蚧菌(Lecdanicillium attenuatum)介导的RNAi对柑橘粉虱毒力探究[D].重庆:西南大学,2015.
[20] 申慧荣.贡嘎蝙蝠蛾粉拟青霉的遗传多样性及生物学特性研究[D].成都:四川农业大学,2014.
[21] 安建梅.虫生真菌粉质拟青霉(Paecilomyces farinosus)培养条件的研究[J].陕西师范大学学报(自然科学版),2003,17(1):72-76.
[22] 曾伟,张德利,涂永勤,等.贡嘎蝠蛾幼虫拟青霉病病原生物学特性研究[J].特产研究,2008(2):14-16.
[23] 张伟,吕利华,何余容.环境因子及常用农药对玫烟色棒束孢 SCAU-IFCF01孢子萌发的影响[J].中国生物防治,2013,29(1):49-55.
责任编辑:罗慧敏
英文编辑:罗 维
Biological characteristics and miticide activity of Paecilomyces farinosus to Panonychus citri
LONG Yanling1, HU Junhua1*, WANG Fan1, CHEN Ruoni2, CHEN Shijiang3, QIN Yuling2, QIN Shaorong2
(1.Citrus Research Institute, Southwest University, Chongqing 400712, China;2.Taiji Industry (Group) Co. Ltd.,Chongqing 400712, China;3.Academy of Chinese Materia Medica, Ganzi District Kangding Gongga Chinese Caterpillar Fungus Co. Ltd., Chongqing 400712, China)
The miticide activity of Paecilomyces farinosus strain nq1-1 to Panonychus citri(McGregor) was determined by the method of indoor toxicity, and single factor tests were used to explore the biological characteristics of the strain.The results indicated that the LC50and LT50of strain nq1-1 to Panonychus citri(McGregor) was respectively 1.59×107/mL and 7.9 d. SDAY and PDA medium were both the optimum mediums to strain nq1-1 for hyphal growth and sporulation. Sucrose and lysine were conducive to fungal growth and sporulation. The most suitable culture temperature for hyphal growth and sporulation were 20 ℃, and temperature at 25-28 ℃ was most conductive to the spore germination. The optimum pH for hyphal growth, sporulation and spore germination was 6.0. UV had no significant effect on strain nq1-1. Moreover, strain nq1-1 had a good compatibility with 82.6% cuprous oxide WG, 75%chlorothalonil WP, 50% oxine-copper WP. In conclusion, strain nq1-1 was toxic to Panonychus citri(McGregor), and grew best under slightly low temperature, neutral pH and high organic nutrition.
Paecilomyces farinosus; Panonychus citri(McGregor); miticide activity; biological control
Q949.32
A
1007-1032(2017)04-0411-06
2017-01-20
2017-05-19
“十二·五”农村领域国家科技计划项目(2014BAD16B0702-5);农业部公益性行业专项(201103020);太极集团有限公司项目(2015071);重庆市社会民生科技创新专项(cstc2016shmszx1147)
龙艳玲(1993—),女,四川广安人,硕士研究生,主要从事微生物基因组学与次生代谢研究,longevens@163.com;*通信作者,胡军华,博士,副研究员,主要从事微生物基因组学与次生代谢研究,hujunhua@cric.cn
猜你喜欢
草原与草坪(2022年5期)2023-01-29
传染病信息(2022年2期)2022-07-15
吉林农业科技学院学报(2022年1期)2022-05-14
安徽农业科学(2020年10期)2020-06-08
麦类作物学报(2019年6期)2019-07-24
西南农业学报(2018年11期)2018-12-11
安徽农业科学(2018年18期)2018-05-14
江苏农业科学(2016年4期)2016-06-14
中国当代医药(2015年26期)2015-03-01
现代检验医学杂志(2015年4期)2015-02-06
