
不同水深条件下沉水植物苦草(Vallisnerianatans)的形态响应和生长策略*
2017-05-17 03:05顾燕飞方根生
湖泊科学 2017年3期
顾燕飞,王 俊,王 洁,方根生,韩 璐**
(1:上海园林绿化建设有限公司,上海 200333) (2:上海市崇明水生水环境研究所,上海 202161) (3:上海水源地建设发展有限公司,上海 200092)
不同水深条件下沉水植物苦草(Vallisnerianatans)的形态响应和生长策略*
顾燕飞1,王 俊1,王 洁1,方根生2,3,韩 璐2,3**
(1:上海园林绿化建设有限公司,上海 200333) (2:上海市崇明水生水环境研究所,上海 202161) (3:上海水源地建设发展有限公司,上海 200092)
苦草(Vallisnerianatans)为我国常见的沉水植物之一,目前对水生态修复工程后苦草种群生长策略尚缺乏深入了解. 以水生态修复工程1年后的苦草种群为研究对象,探讨水深对苦草叶片、匍匐茎、块茎等形态特征和苦草株数、匍匐茎数以及生物量等种群参数的影响,并分析水深胁迫下苦草的生长策略. 结果表明,随着水深增加,尽管苦草单位面积株数和单位面积匍匐茎数均显著降低,但叶片长度、叶片宽度、叶片厚度、叶面积、匍匐茎长度、匍匐茎粗和块茎粗随水深显著增加,因而,苦草地上生物量也显著增加. 其中,叶片长度和叶面积变异较大,对水深变化敏感,而叶片宽度、叶片厚度、匍匐茎长度、匍匐茎粗和块茎粗变异不大,对水深变化敏感程度低. 在低光照胁迫下,苦草在亲株生长和子株输出之间存在权衡,一方面通过增加叶片投资保障亲株的资源利用效率,一方面通过降低亲株密度保障子株生长发育,从而实现种群空间生态位扩展和持续更新. 在水生态修复工程中,考虑苦草的种群更新能力和施工的可操作性,在透明度为1.0~1.5 m的条件下,苦草的适宜种植水深为0.5~1.0 m.
富营养化;生态修复;形态响应;生长策略;沉水植物;苦草
苦草(Vallisnerianatans)是我国常见的沉水植物之一,在长江中下游浅水湖泊、河流广泛分布[1],亦有较高的经济利用价值[2],在目前的富营养化水体生态修复工程中广泛应用[3].
植物是一种构件生物,对环境条件的响应主要体现在构件的形态和数量变化上[4]. 沉水植物的生长主要受水深、营养盐浓度和底质条件限制[5]. 苦草为水体底层植物,光补偿点低,能够忍受弱光[6],其生长发育尤其受水深的影响,这种影响必然反映在叶片、匍匐茎和根系等构件的形态和数量上. 有研究表明在水深2 m 范围内,刺苦草(Vallisneriaspinulosa)的地上和地下生物量随水深增加而增加[7]. Xiao等[8]研究也表明水深显著影响苦草的分株生物量、分株数量、世代数和匍匐茎长度等. 而温文科等[9]则报道了苦草密度与水深呈负相关,而平均叶长与水深呈正相关. 但是自然水域中有关不同水深条件下苦草叶片、匍匐茎等构件数量和形态特征的原位研究尚不多见.
在水环境生态修复过程中,恢复沉水植被是重要手段之一,对于维持水生生态系统功能完整性具有重要作用,探明不同水深条件下苦草的相关生物生态学特征的变化对于沉水植被恢复和水质的维持尤为关键. 目前有关苦草的生活史特征、水质净化作用和生理生态学方面已有较多报道[10-12]. 本文以城市景观水体生态修复后人工建植的苦草群落为研究对象,通过分析苦草密度和生物量等种群统计参数以及苦草叶片、匍匐茎和块茎等个体形态指标,揭示不同水深条件下苦草形态特征和生长策略的变化,为苦草沉水植被恢复和管理提供理论依据.
1 材料与方法
1.1 实验地点
研究地点位于上海市曲阳公园(30°17′19″~31°17′26″N,121°28′47″~121°29′01″E),该水体于2014年进行生态修复,栽植苦草恢复沉水植被. 经过一年的恢复,目前苦草生长良好,主要在距离湖岸线10 m的范围内呈环状分布,岸边主要栽培再力花(Thaliadealbata)、白花梭鱼草(Pontederiacordata)、千屈菜(Lythrumsalicaria)、菖蒲(Acoruscalamus)、黄菖蒲(Irispseudacorus)等挺水植物. 湖区水位依靠雨水作为补水调节,总体水位波动不大,水深维持在2.0~2.5 m. 底质主要是砂质壤土.

图1 研究区域及调查样点示意图Fig.1 Study area and location of samples
1.2 野外调查
在2015年8月进行野外调查. 调查时,主要依据水深(water depth,WD)设置样地,同时考虑岸边柳树和建筑物遮阴对苦草生长的影响. 总计设置7个样地,编号分别为P1~P7. 样地P1、P2、P3和P4为岸边浅水生境,平均水深分别为26.7、51.3、60.5和41.0 cm,其中样地P1、P2受遮阴影响,样地P3则为开阔水面,无遮阴影响. 样地P1、P2和P3无淤泥层. 样地P4较为特殊,为睡莲(Nymphaeasp.)、黄菖蒲种植区,不仅受岸边柳树的遮阴影响,还受睡莲和黄菖蒲的生物竞争影响,淤泥厚度约2~4 cm. 样地P5、P6和P7为深水生境,无遮阴影响,平均水深分别为101.5、153.5和169.5 cm,淤泥厚度分别约3、2和4~5 cm.
取样时,每个样地设置3个样方,取样方法为将苦草和土块一同挖起,在岸上将土块切割成长方形,现场测量长、宽,计算样方面积. 带回室内后,每个样方选取长势均匀的5株成株作为样株,测量单株叶片数(leaf number, LN)、叶片长度(leaf length, LL)、叶片宽度(leaf width, LW)、叶片厚度(leaf thickness, LT)、叶面积(leaf area, LA)、匍匐茎长度(stolon length, SL)、匍匐茎粗(stolon diameter, SD)、块茎粗(tuber diameter, TD)8个个体形态指标以及株数(plant number, PN)、匍匐茎数(stolon number, SN)、地上生物量(aboveground biomass, AB)和地下生物量(underground biomass, UB) 4个种群参数. 其中,8个个体形态指标以无性系分株为统计单位,单株叶片数为单个无性系分株的叶片数,每个分株选取最大叶片测量相关指标,叶片长度和叶面积为单个叶片长度和叶面积,叶片宽度和厚度为叶片最宽处的宽度和厚度,叶片样本数量n=15. 4个种群参数以样方为统计单位,株数为样方内无性系分株数,匍匐茎长度和匍匐茎粗分别为样方内所有匍匐茎长度和匍匐茎粗的均值,匍匐茎粗测量位置为匍匐茎的中间部位,匍匐茎数为样方内匍匐茎数量,块茎粗为地上、地下过渡部位的茎粗,生物量为风干重.
1.3 数据分析
在统计株数、匍匐茎数、地上生物量和地下生物量等种群参数时,将实测样方面积转换为单位面积,使用转换后的值进行后续的数据分析. 应用Duncan多重比较检验各指标在7个样地中的差异显著性. 使用变异系数查看各指标的变异情况,变异系数CV(%)=100×标准偏差/平均值. 应用Pearson相关系数考察各指标间及各指标与水深间的相关性. 所有统计的显著水平为P=0.05或P=0.01. 多重比较和Pearson相关性分析使用SPSS 13.0 (SPSS Inc., Chicago, USA) 软件完成.
2 结果
2.1 苦草种群特征变化
在4个种群统计参数中,单位面积株数和单位面积匍匐茎数呈现出较一致的变化规律,均以样地P2和P3最高,与其他5个样地差异达到统计显著水平(P<0.05,图2a、2g),相关性分析显示二者呈极显著正相关(r=0.912,P<0.01),并均与水深表现出负相关关系(表1). 如果以单位面积匍匐茎数与单位面积株数的比值作为苦草个体繁殖输出的衡量指标,则7个样地中,繁殖输出较高的为样地P2、P3、P6和P7,其次为样地P4和P5,样地P1最低(图3).
地上生物量和地下生物量变化规律不同. 地上生物量以样地P7最高,显著高于其他样地(P<0.05),其次为样地P2、P3、P5和P6,且相互间差异不显著,样地P1和P4则显著偏低(图2k). 地下生物量以样地P2、P3和P7较高,而样地P1、P4和P5的较低(图2l). 从偏相关分析结果看,地上生物量与叶片长度(r=0.631,P<0.01)和水深(r=0.769,P<0.01)均呈极显著正相关,但与单位面积株数的相关性不显著(P>0.05),地下生物量与单位面积株数呈极显著正相关(r=0.767,P<0.01),而与根粗、水深等指标无显著相关性. 从生物量分配格局看,随着水深增加,地上生物量呈增加趋势,而地下生物量呈减少趋势 (图4).
单位面积株数、单位面积匍匐茎数、地上生物量和地下生物量4个种群参数在7个样地中表现出较大的变异,变异系数均在60%以上(图5),但是,仅地上生物量与水深呈显著正相关,其他3个参数与水深相关性不显著(表1),表明水深并不是引起这几个种群参数产生变异的主要因素,存在其他尚未观测的重要环境变量.

图2 苦草种群特征和个体形态指标在各样地中的变化(不同字母代表差异显著(P<0.05),下同)Fig.2 Changes of population parameters and individual morphological indexes of Vallisneria natans among plots (Different letters indicate that differences are significant (P<0.05), the same below)
2.2 苦草个体形态指标变化
从叶片形态指标来看,单株叶片数、叶片宽度和叶片厚度在7个样地间变化不大(图2b、2d、2e),变异系数均低于40%,介于19.18%~29.89%之间. 单株叶片数与叶片长度(r=0.577,P<0.05)、叶片宽度(r=0.702,P<0.01)和叶片厚度(r=0.664,P<0.01)均呈显著正相关,但与水深相关性不显著(P>0.05). 叶片长度和叶面积在各样地中变化规律一致,均以样地P6和P7较高,样地P1、P2和P3较低(图2c、2f),两个指标的变异系数分别为50.5%和67.5%(图5),最高值分别是最低值的3倍多和5倍多,表现出较大的可塑性. 而叶片宽度和叶片厚度尽管总体上表现出增加的趋势,但变异程度低于叶片长度和叶面积,分别为21.0%和24.1%(图5). 叶片长度、叶片宽度、叶片厚度、叶面积之间以及与水深之间均呈现显著的两两正相关关系(表1).
匍匐茎是苦草种群扩展的主要方式,匍匐茎的数量和长度决定了其扩展能力. 匍匐茎长度和匍匐茎粗在7个样地间的变异程度低,变异系数分别为25.5%和26.9%(图5),但是从样地P1~P7,匍匐茎长度和匍匐茎粗整体上均呈增加的趋势(图2h、2i),与水深、叶片长度、叶片宽度、叶片厚度和叶面积呈显著正相关(P<0.01,表1). 表明随着植株体大小的增加,匍匐茎的扩展能力亦增加(图2h、2i).

图3 苦草单位面积匍匐茎数与单位面积株数比值在各样地中的变化Fig.3 Changes of the ratio of stolon number per area and plant number per area of Vallisneria natans among plots
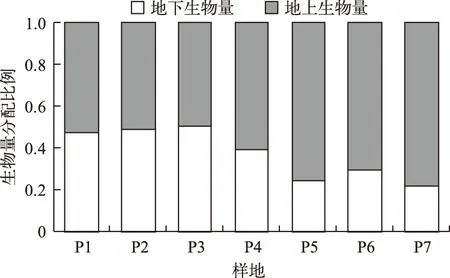
图4 各样地苦草生物量分配格局的变化Fig.4 Changes of biomass allocation patterns of Vallisneria natans among plots

图5 苦草各形态指标变异系数变化Fig.5 Changes of coefficient variation among morphological indexes of Vallisneria natans
块茎粗在7个样地间变异程度也较低,变异系数为29.9%(图5),从样地P1~P7表现出增加的趋势(图2j),与水深、叶片长度、叶片宽度、叶片厚度、叶面积以及匍匐茎长度和匍匐茎粗均呈显著正相关(P<0.01),而与单位面积株数和单位面积匍匐茎数呈显著负相关(P<0.05,表1).
3 讨论
3.1 不同水深条件下苦草的形态响应
植物是构件生物,通过调节构件的形态和数量适应环境的变化,但是,不同的构件响应方式和响应程度有所不同. 本研究表明,在测量的个体形态指标中,苦草叶片长度、叶片宽度、叶片厚度和叶面积等指标均随着水深增加而增加,其中,叶片长度、叶面积对水深的变化最为敏感,其变异系数分别达到50.5%和67.5%,而叶片宽度、叶片厚度、匍匐茎长度、匍匐茎粗和块茎粗变异系数均不足40%,对水深变化敏感程度相对较低. 而且,单位面积株数、单位面积匍匐茎数、地下生物量与水深之间并不存在显著的相关性,这3个指标的变化很不规则,其中样地P1和P4的单位面积株数、单位面积匍匐茎数和地下生物量明显偏低,而同为浅水生境的样地P2、P3的3个指标却出现最高值(图2),但是样地P2存在遮阴影响,而样地P3不存在遮阴的影响,综合这几个方面看,可以说明以下几个问题:1)在浅水条件下,岸边柳树和建筑物的遮阴作用对苦草植株和匍匐茎数量影响微弱;2) 睡莲、黄菖蒲等通过遮阴、资源竞争等生物作用能够显著地降低苦草植株和匍匐茎密度;3)存在其他尚未观测的重要生态因子限制苦草的生长发育,包括底质条件等. 如Xie等[13]研究表明苦草的生物量分配主要受底质条件影响,而非上覆水的营养元素. Xiao等[8]和谢贻发等[14]的研究结果也都表明苦草的生物量累积和分配主要受底质类型影响. 因此,解释本研究中样地P1的各种群参数偏低的情况还需要进一步开展底质条件的相关分析.
曹昀等[15]采用控制实验的方法研究了60~170 cm水深下苦草的生长和生物量分配,结果表明苦草的叶片长度、叶片宽度、叶面积都随着水深的增加而减小,且在150~170 cm水深范围内叶片长度明显变小,但对生物量的分配作用不明显. 而本研究中,水深范围为26.7~170 cm,但苦草叶片长度、叶片宽度和叶面积却随着水深增加而增加. 据符辉等[16]报道,水深>4 m才对苦草克隆生长和地上生物量产生明显的抑制作用. 因此,水深对苦草生长的影响还与透明度有关. 另据温文科等[9]报道,在水深120 cm范围内,苦草密度与水深呈负相关,而平均叶长与水深呈正相关. 本研究结果与曹昀等[15]结果出现差异与实验中苦草的生长周期和水体透明度有关,在曹昀等的报道中,实验周期为60 d,水体透明度约80 cm,而本研究中水体透明度在1 m以上,并且,苦草已经栽植1年,经过了一个完整的生长季,这可能导致实验结果出现差异.

表1苦草各形态指标与水位间偏相关检验(n=21)
*P<0.05,**P<0.01;水深WD;单位面积株数PN;单位面积匍匐茎数SN;叶片长度LL;叶片宽度LW;叶片厚度LT;单株叶片数LN;匍匐茎长度SL;匍匐茎粗SD;块茎粗TD;叶面积LA;地上生物量AB;地下生物量UB.
3.2 深水胁迫下苦草无性繁殖特征和生长策略
苦草的繁殖输出主要依靠无性繁殖,匍匐茎是其种群扩展的主要方式[17]. 在本研究中,苦草的繁殖输出主要受单位面积株数、单位面积匍匐茎数、匍匐茎长度、匍匐茎粗等因素的影响. 随着水深增加,苦草的单位面积株数、单位面积匍匐茎数表现为降低趋势,匍匐茎长度和匍匐茎粗则表现为增加趋势,而单位面积匍匐茎数与单位面积株数比值的变化并不明显. 表明尽管水深增加没有显著影响苦草个体的繁殖输出,但会降低种群密度,从而间接减少整个种群的繁殖输出.
对沉水植物而言,由于水深增加降低了光照强度,并间接改变水体温度、养分、底泥再悬浮等[16],因此,水深增加实际上是一种环境胁迫. 植物面对环境胁迫需要寻找一种权衡,一方面保证亲株的资源获取能力和种群规模,一方面保证子株输出,维持种群的续存. 就生物学特性而言,苦草分布于水体底层,其繁殖方式是产生匍匐茎,茎顶端发生顶芽分化,产生分株,这与穗花狐尾藻(Myriophyllumspicatum)、轮叶黑藻(Hydrillaverticillata)等水体中上层植物不同,在此前提下,如果苦草在深水条件下产生大量分株,会进一步降低底层的光照,子株将会因得不到足够的光照而无法正常发育,即亲株与子株之间对光资源存在非对称性竞争,这与森林生态系统中冠层树种对底层幼树的邻域效应(neighborhood effect)类似. 据此,可以认为,在水深胁迫下,一方面,苦草通过增加叶片的生物量分配,获取更多的光资源,以此来保证亲株的资源利用效率,另一方面,降低亲株密度,为底层幼苗生长提供必要的光照条件,并通过增加匍匐茎粗来提高亲株向子株的养分输出,形成有效的生理整合,已有研究表明苦草匍匐茎中存在大量淀粉等营养物质[18],另外,还通过增加匍匐茎长度来降低亲株对子株由于非对称性竞争而产生的负面作用,通过这种方式在亲株生长和子株输出之间找到平衡点,以此兼顾种群的空间生态位扩展和持续更新.
3.3 水环境生态修复工程中苦草的管理
在水环境生态修复工程中,作为初级生产者,发育良好的沉水水生植被对于维持水质、营造健康的水生生态系统至关重要. 通过本项研究,初步证实水深显著影响苦草的种群特征和个体形态特征,尤其影响苦草的密度、叶片长度和地上生物量分配,在深水环境中,苦草通过调节亲代的种群密度实现子代的更新,但是苦草本身作为一种能够忍受弱光的底层沉水植物,在浅水条件下,遮阴对其生长影响十分微弱. 这些结果提示我们,在构建苦草沉水植被时,考虑苦草的种群更新能力、施工的可操作性和施工成本,在透明度为1.0~1.5 m的条件下,苦草的适宜种植水深为0.5~1.0 m,能够保障苦草种群具有较高的种群密度和较强的更新能力,有效抑制底泥再悬浮;在超过1 m的区域,苦草以稀植为宜,可以保障种群的生态位扩展和持续更新.
[1] Chen Lei, Ye Qigang, Pan Lizhuetal.Vallisneriaspecies in lakes of the middle-lower reaches of the Yangtze River of China.JournalofPlantEcology(ChineseVersion), 2008, 32(1): 106-113.[陈磊, 叶其刚, 潘丽珠等. 长江中下游湖泊两种混生苦草属植物生活史特征与共存分布格局. 植物生态学报, 2008, 32(1): 106-113.]
[2] Wang Yanli, Xiao Yu, Pan Huiyunetal. Analysis of nutrient composition and comprehensive utilization of submersed aquatic macrophytes (Vallisnerianatans).JournalofEcologyandRuralEnvironment, 2006, 22(4): 45-47, 70. [王艳丽, 肖瑜, 潘慧云等. 沉水植物苦草的营养成分分析与综合利用. 生态与农村环境学报, 2006, 22(4): 45-47, 70.]
[3] Ma Jianmin, Yan Guoan, Ren Nanetal. Restoration and structural optimization of aquatic vegetation in enclosure and fence in East Lake of Wuhan.ChineseJournalofAppliedEcology, 1997, 8(5): 535-540. [马剑敏, 严国安, 任南等. 东湖围隔(栏)中水生植被恢复及结构优化研究. 应用生态学报, 1997, 8(5): 535-540.]
[4] Xiao Keyan, Yu Dan. Concepts and studies on clonal population of aquatic plant.ActaHydrobiologicaSinica, 2008, 32(6): 920-925. [肖克炎, 于丹. 克隆种群的有关概念在水生植物中应用和研究进展. 水生生物学报, 2008, 32(6): 920-925.]
[5] Xie Yifa. Study on interaction of submerged macrophytes and nutrients in the water and sediment[Dissertation]. Guangzhou: Jinan University, 2008. [谢贻发. 沉水植物与富营养湖泊水体、沉积物营养盐的相互作用研究[学位论文]. 广州: 暨南大学, 2008.]
[6] Su Wenhua, Zhang Guangfei, Zhang Yunsunetal. The photosynthetic characteristics of five submerged aquatic plants.ActaHydrobiologicaSinica, 2004, 28(4): 391-395. [苏文华, 张光飞, 张云孙等. 5种沉水植物的光合特征. 水生生物学报, 2004, 28(4): 391-395.]
[7] Yuan Longyi, Li Shouchun, Li Weietal. Effects of water depth on growth and reproductive strategy ofVallisneriaspinulosa.JournalofJiangxiNormalUniversity(NaturalScience), 2007, 31(2): 156-160. [袁龙义, 李守淳, 李伟等. 水深对刺苦草生长和繁殖策略的影响研究. 江西师范大学学报: 自然科学版, 2007, 31(2): 156-160.]
[8] Xiao Keyan, Yu Dan, Wu Zhonghua. Differential effects of water depth and sediment type on clonal growth of the submersed macrophyteVallisnerianatans.Hydrobiologia, 2007, 589(1): 265-272.
[9] Wen Wenke, Shao Liu, Wu Jianyongetal. Distribution properties and effects of light intensity onVallisnerianatansin Litang River.JournalofShanghaiOceanUniversity, 2014, 23(6): 897-903. [温文科, 邵留, 吴建勇等. 里塘河亚洲苦草分布特点及光照因子影响分析. 上海海洋大学学报, 2014, 23(6): 897-903.]
[10] Geng Xianhua, Yu Dan, Huang Yongmingetal. Growth and physio-biochemistry responses ofVallisneriaSpiralisL. to CO2enrichment.ActaHydrobiologicaSinica, 2004, 28(3): 304-309. [耿显华, 于丹, 黄永明等. 高浓度CO2下苦草的生长和生理生化反应.水生生物学报, 2004, 28(3):304-309.]
[11] Li Kuanyi, Liu Zhengwen, Wang Chunzhongetal. Effect of low dissolved oxygen on the growth ofVallisneriaSpiralis.ResourcesandEnvironmentintheYangtzeBasin, 2006, 15(5): 670-673. [李宽意, 刘正文, 王春忠等. 低溶解氧对苦草生长的影响. 长江流域资源与环境, 2006, 15(5): 670-673.]
[12] Wang Lizhi, Wang Guoxiang. Effects ofVallisnerianatansdensity on phosphorus release by disturbance.EcologyandEnvironmentalSciences, 2012, 21(6): 1096-1101.[王立志, 王国祥. 苦草密度对扰动引起各形态磷释放的影响. 生态环境学报, 2012, 21(6): 1096-1101.]
[13] Xie Yonghong, An Shuqing, Wu Bofeng. Resource allocation in the submerged plantVallisnerianatansrelated to sediment type, rather than water-column nutrients.FreshwaterBiology, 2005, 50(3): 391-402.
[14] Xie Yifa, Li Chuanhong, Liu Zhengwenetal. Effects of sediments on the growth and morphology ofVallisnerianatans.JournalofAgro-EnvironmentScience, 2007, 26(4): 1269-1272. [谢贻发, 李传红, 刘正文等. 基质条件对苦草(Vallisnerianatans)生长和形态特征的影响. 农业环境科学学报, 2007, 26(4):1269-1272.]
[15] Cao Yun, Zhang Sujuan, Liu Yanyanetal. Effects of water gradient on seedlings growth and biomass ofVallisnerianatans.EcologyandEnvironmentalSciences, 2014, 23(8): 1332-1337. [曹昀, 张素娟, 刘燕燕等. 水深梯度对苦草生长和生物量的影响. 生态环境学报, 2014, 23(8):1332-1337.]
[16] Fu Hui, Yuan Guixiang, Cao Teetal. Clonal growth and foraging behavior of a submerged macrophyteVallisnerianatansin response to water depth gradient.JLakeSci, 2012, 24(5): 705-711. DOI:10.18307/2012.0510. [符辉, 袁桂香, 曹特等. 水深梯度对苦草(Vallisnerianatans)克隆生长与觅食行为的影响. 湖泊科学, 2012, 24(5): 705-711.]
[17] Donnermeyer GN, Smart MM. The biomass and nutritive potential ofVallisneriaamericanaMichx in navigation pool 9 of the upper Mississippi river.AquaticBotany, 1985, 22(85): 33-44.
[18] Wang Yongping, Qin Boqiang. A comparison on morphology and anatomy ofVallisnerianatansfrom oligotrophic and eutrophic lakes.JLakeSci, 2011, 23(1):56-60. DOI:10.18307/2011.0109. [王永平, 秦伯强. 贫、富营养湖泊中苦草的形态解剖结构比较. 湖泊科学, 2011, 23(1): 56-60.]
Morphological response and growth strategy of the submerged macrophyteVallisnerianatansunder different water depths
GU Yanfei1, WANG Jun1, WANG Jie1, FANG Gensheng2,3& HAN Lu2,3**
(1:ShanghaiGardening-landscapingConstructionCo.Ltd.,Shanghai200333,P.R.China) (2:ChongmingInstituteofAquaticOrganismsandWaterEnvironment,Shanghai202161,P.R.China) (3:ShanghaiWaterSourceConstructionDevelopmentCo.Ltd.,Shanghai200092,P.R.China)
Vallisnerianatansis one of the most common submerged macrophytes in lakes of China. In present, the growth strategies ofV.natanspopulation in the water ecological restoration project remain unclear. In this paper, the responses ofV.natansmorphological traits such as leaf, stolon and tuber and population parameters such as plant number, stolon number and biomass to water depth were discussed on basis of one year effects after the implementation of water ecological restoration project. The growth strategies ofV.natanswere also analyzed. The results showed that the plant number per area and stolon number per area significantly decreased with the increasing of water depth, whereas the above-ground biomass increased significantly. The leaf length, leaf width, leaf thickness, leaf area, stolon length, stolon diameter and tuber diameter also increased significantly with the water depth increasing. Among them, the leaf length and leaf area varied dramatically because they were sensitive to water depth, whereas leaf width, leaf thickness, stolon length and diameter, and tuber diameter varied lightly and were insensitive to water depth. A trade-off between parent shoot growth and daughter shoot output was found under stress of less light.V.natansguarantees the resource utilization efficiency of parent shoots by increasing investments to leaf. On the other hand,V.natansimproves the growth of daughter shoots by decreasing the parent shoot density, and thus the plant realizes the spatial niche expansion and population regeneration. In water ecological restoration project, considering the regeneration ability ofV.natanspopulation and construction maneuverability, water depth at appropriate 0.5 m to 1.0 m is suggested. Keywords: Eutrophication; ecological restoration; morphological response; growth strategy; submerged macrophyte;Vallisnerianatans
*上海建工集团股份有限公司重点科研项目课题(14JCSF-25)和上海市崇明科委项目(CKS2015-03)联合资助. 2016-03-09收稿; 2016-06-24收修改稿. 顾燕飞(1975~),女,本科,高级工程师;E-mail:188701219@qq.com.
;E-mail: hl_821212_2004@126.com.
猜你喜欢
安徽化工(2022年1期)2022-02-15
科学技术创新(2021年22期)2021-08-16
至爱(2020年10期)2020-11-20
临床检验杂志(电子版)(2020年1期)2020-04-03
新农民(2019年9期)2019-02-19
中南民族大学学报(自然科学版)(2018年4期)2018-12-29
吉林农业(2018年18期)2018-09-25
江苏农业科学(2017年3期)2017-05-02
学苑创造·B版(2015年12期)2016-06-23
江苏农业科学(2015年5期)2015-10-20
