
马铃薯匍匐茎糖类物质变化规律的研究
2017-05-02 11:47张桂芝姜海洋马光恕廉华杨升张军
江苏农业科学 2017年3期
张桂芝 姜海洋 马光恕 廉华 杨升 张军民

摘要:以克新13号、费乌瑞它、延薯4号3个熟期不同的马铃薯品种为试验材料,采用大田旱作模式、随机区组设计,通过测定苗后5~30 d匍匐茎中糖的成分和含量以及糖代谢相关酶的活性,研究马铃薯匍匐茎发生和形成过程中糖类物质变化规律。结果表明,蔗糖、可溶性糖的含量呈现先上升后下降再回升的变化趋势(延薯4号的可溶性糖含量除外),葡萄糖、果糖的含量则是先降低再升高的趋势;蔗糖含量与SS活性的变化趋势近似,SPS活性呈现“M”形,转化酶两谷一峰变化趋势。SS、SPS是影响蔗糖合成的关键酶,转化酶是分解转化蔗糖的关键酶。匍匐茎决定着块茎的形成,通过此试验研究旨在为马铃薯块茎形成提供理论依据,为指导马铃薯的高产优质栽培提供技术支撑。
关键词:马铃薯;匍匐茎;糖代谢;酶活性
中图分类号: S532.01文献标志码: A
文章编号:1002-1302(2017)03-0055-04
收稿日期:2015-12-22
基金项目:2011年度黑龙江省新农村科技服务计划项目(编号:1251xnc111)。
作者简介:张桂芝(1969—),女,硕士,高级农艺师,从事蔬菜育种及生理研究工作。E-mail:damao98@163.com。
通信作者:马光恕,教授,主要从事蔬菜栽培生理研究。E-mail:mgs_lh@163.com。
2015年,马铃薯被我国确立为主粮作物。马铃薯营养丰富且耐旱、抗性强、适宜种植范围广,可兼做粮食、蔬菜、饲料和工业原料,对亚洲乃至全世界72亿人口的粮食安全都是至关重要的[1-2]。
糖是植物光合作用的产物,是植物器官组织赖以生长发育的重要能量和物质基础[3]。蔗糖、葡萄糖、果糖等可溶性糖是植物糖类物质的重要组分,蔗糖是植物体内碳水化合物贮存和运输的主要形式,协调植物源库关系的信号分子,蔗糖和相关酶[蔗糖合成酶(SS)、蔗糖磷酸合成酶(SPS)、转化酶]在植物糖代谢中扮演着重要角色,影响器官的发生和果实品质形成[4-5]。转化酶与蔗糖合成酶的共同作用是影响普通栽培型番茄果实中糖积累的重要因子[6],高浓度的糖是促进块茎发育的重要因子[7],相对于其他可溶性单、双糖类,蔗糖诱导结薯的能力更突出和高效[8]。对番茄[6]、大豆[9]、甜菜[10]、玉米[11]、大葱[12]等作物的研究表明可溶性糖代谢及其酶活性变化与植物的生长发育密切相关,SS既能催化蔗糖合成又能催化其分解,SPS被认为是催化蔗糖合成的主要酶,转化酶催化蔗糖分解为单糖[13]。
匍匐茎的形成和发育直接影响块茎形成进而影响产量,前人研究了匍匐茎发生形态变化、发生节位、肥料、生长物质、光照等因素作用[14-18],而匍匐茎的发生和形成過程中自身糖类物质代谢及相关酶活性变化情况尚未见报道,所以研究匍匐茎的糖类物质变化规律,了解和掌握匍匐茎发生生长生理机制可以为马铃薯高产高效栽培及合理施肥提供重要理论依据。[LM]
1材料与方法
1.1试验材料
供试马铃薯品种为脱毒一代良种,分别是早熟品种费乌瑞它、中晚熟品种克新13号与延薯4号。
1.2试验方法
试验设在黑龙江省大庆市大同区航天育种基地,土壤类型为草甸黑钙土,0~20 cm耕层土壤基本农化性状为:土壤碱解氮178.00 mg/kg,速效磷20.00 mg/kg,速效钾 240.00 mg/kg,有机质含量2.45%,pH值8.04,含盐总量 0.12%。
试验设计采取随机区组设计,3次重复。大田旱作模式,垄作方式种植,人工点播。小区为10行区,行长10 m,行距0.65 m,株距0.24 m,试验区四周设两行保护行。一次性施入有机肥1.5万kg/hm2,磷酸氢二铵120 kg/hm2,尿素 210 kg/hm2,硫酸钾270 kg/hm2。2015年4月26日播种,5月26日出全苗,生育期间进行常规田间管理。
1.3项目测定
田间出苗后及时分别挂牌,出苗后至块茎形成期每隔5 d随机取样,进行马铃薯匍匐茎形成期糖代谢指标的测定,共取样6次。
可溶性糖、蔗糖、葡萄糖、果糖含量的测定均采用蒽酮比色法[19];还原糖含量的测定参考李合生等的方法[20],采用 3,5-二硝基水杨酸法;蔗糖合成酶、蔗糖磷酸合成酶活性,酶的提取和测定参照Nielsen等的方法[21];转化酶活性参照Zhu等的方法[22]提取。
1.4数据分析
试验数据采用Excel、DPS软件进行统计分析。
2结果与分析
2.1匍匐茎糖类物质含量变化
2.1.1匍匐茎蔗糖的含量变化规律
马铃薯匍匐茎蔗糖含量变化规律如图1所示:不同品种马铃薯匍匐茎蔗糖含量在出苗后5~30 d均呈现先升高后缓慢下降的变化趋势。出苗后10 d,3个品种匍匐茎蔗糖含量均达到峰值,费乌瑞它增长幅度最高,为0.69 mg/(g·d);克新13号增长幅度最低,为 0.24 mg/(g·d)。出苗后10~20 d快速下降,在出苗后 20 d,克新13号降至最低值,后缓慢上升。费乌瑞它和延薯4号则在出苗后25 d时降至最低值,后有所回升。
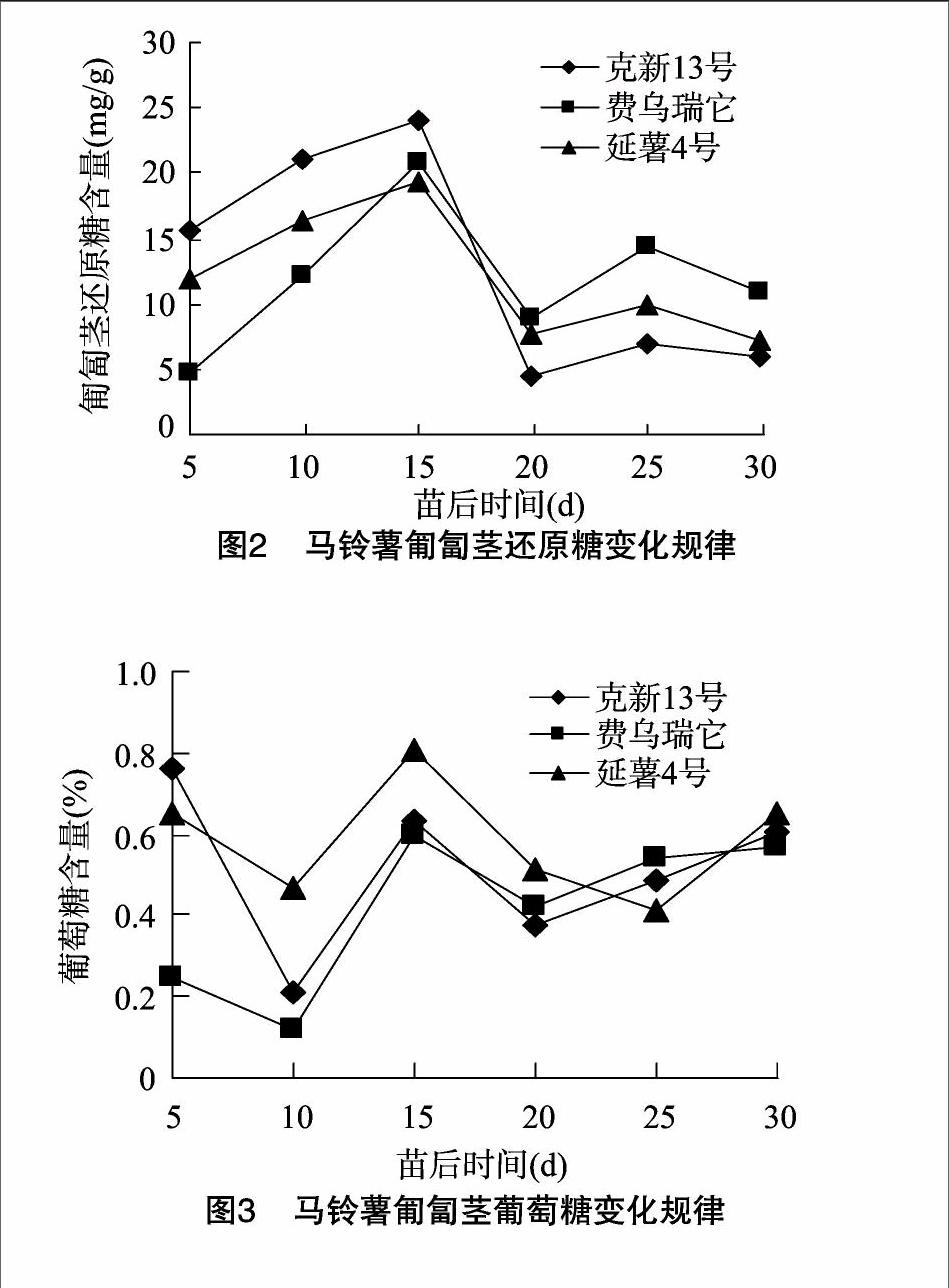
2.1.2匍匐茎还原糖的含量变化规律
由图2可以看出,不同马铃薯品种匍匐茎中的还原糖含量在苗后5~30 d均呈现先升高然后迅速下降后再小幅度上升后下降的变化趋势。出苗后15 d,3个品种匍匐茎还原糖含量均达高峰值,费乌瑞它增长幅度最高,为1.61 mg/(g·d);延薯4号增长幅度最低,为0.74 mg/(g·d)。出苗后15~20 d快速下降,在出苗后20 d时,克新13号和延薯4号至最低值,费乌瑞它匍匐茎还原糖含量仍高于5 d时的含量;出苗后20~25 d,3个品种匍匐茎还原糖含量均又上升,至25 d时达一个低于15 d时的小高峰,仍以费乌瑞它增长幅度最高,为1.101 mg/(g·d);延薯4号增长幅度最低,为0.46 mg/(g·d);出苗后25~30 d,不同马铃薯品种匍匐茎含量均缓慢下降,但出苗30 d时不同品种马铃薯还原糖含量均高于20 d的含量。
[FK(W10][TPZGZ2.tif][FK)]
2.1.3匍匐茎葡萄糖的含量变化规律
马铃薯匍匐茎葡萄糖含量变化规律如图3所示。不同品种马铃薯匍匐茎葡萄糖含量在出苗后5~30 d均呈现“W”形变化趋势,先下降后升高然后下降后再上升。出苗后10 d,克新13号和费乌瑞它匍匐茎葡萄糖含量降至最低值,克新13号下降幅度最大,费乌瑞它降幅最小,延薯4号亦下降至较低值,但它的葡萄糖含量高于前2个品种;出苗后10~15 d,不同马铃薯品种匍匐茎含量均迅速上升,15 d时达峰值;出苗后15~20 d,匍匐茎葡萄糖含量下降,克新13号和费乌瑞它降至较低值,延薯4号继续下降并在25 d时降到最低值然后上升;出苗20~30 d克新13号和费乌瑞它匍匐茎葡萄糖含量亦逐渐上升。
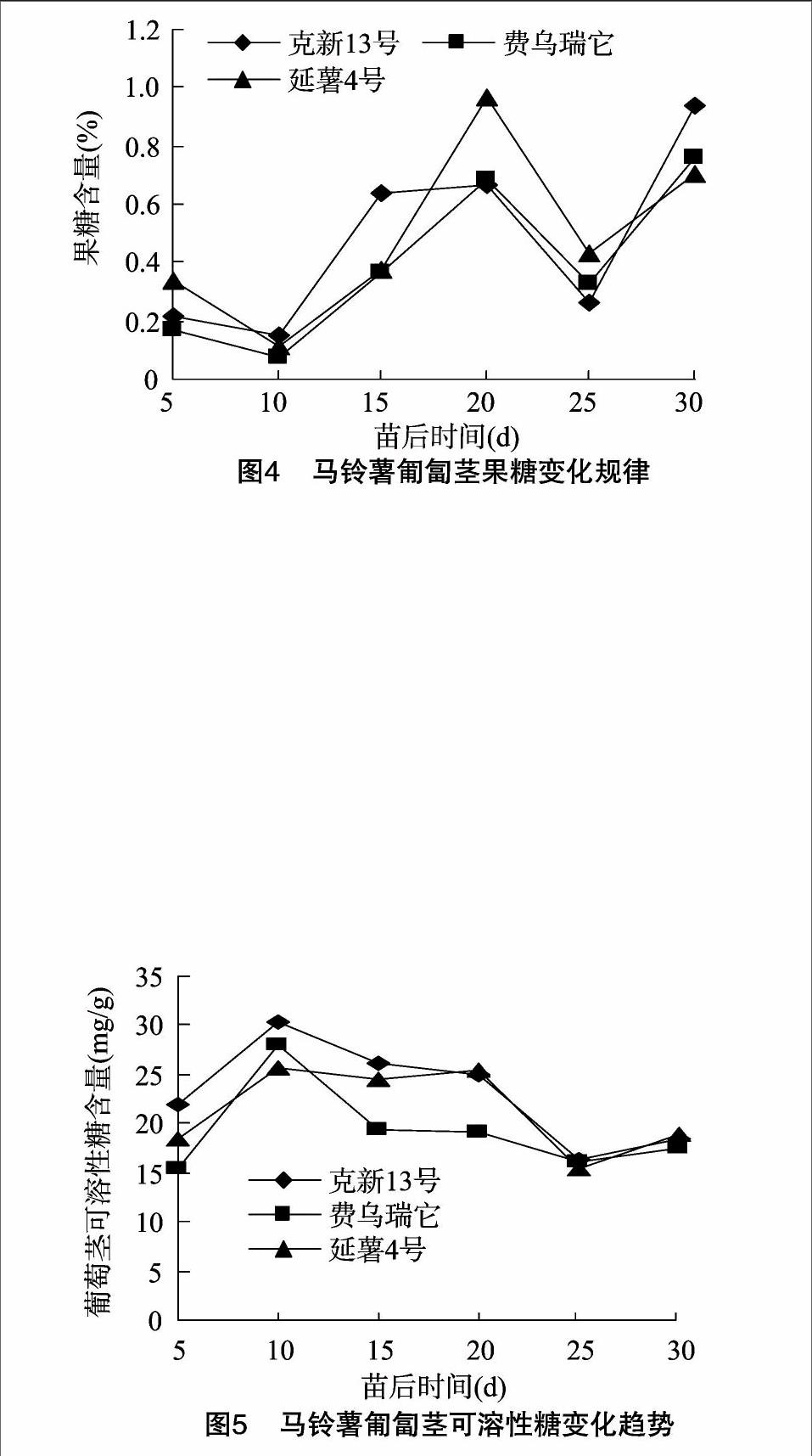
2.1.4匍匐茎果糖含量变化规律
马铃薯匍匐茎果糖含量变化规律如图4所示。不同品种马铃薯匍匐茎果糖含量在出苗后5~30 d均呈现先下降后上升又下降再上升似“W”字形的变化趋势。出苗后10 d,3个品种匍匐茎果糖含量均下降到最低值;出苗后10~20 d快速连续上升,20 d时延薯4号达峰值。延薯4号增幅最大,日增幅为0.09%,克新13号增幅最小,日增幅为0.05%;出苗后20~25 d,不同品种马铃薯匍匐茎果糖含量又再次下降,25 d时降至较低值;25~30 d时迅速上升,克新13号增幅最大,日增幅为0.13%,延薯4号增幅最小,日增幅为0.05%。
2.1.5匍匐茎中可溶性糖含量变化规律
试验结果表明,不同品种马铃薯匍匐茎可溶性糖含量在出苗后5~30 d表现为先上升后逐渐下降再上升的变化趋势(图5,延薯4号除外)。在出苗后5~10 d快速增加,在出苗后10 d达峰值。费乌瑞它增长幅度最高,为2.51 mg/(g·d);延薯4号增长幅度最低,为1.44 mg/(g·d);在出苗后10~25 d时逐渐下降(延薯4号除外),其中在苗后10~20 d可溶性糖含量缓慢下降,在苗后20~25 d时迅速下降;出苗后25~30 d可溶性糖积累量快速增加,延薯4号增加幅度最大。
2.2匍匐茎糖代谢相关酶的活性变化
2.2.1蔗糖合成酶(SS)的活性变化
马铃薯匍匐茎SS活性变化规律如图6所示。不同品种马铃薯匍匐茎SS活性在出苗后5~15 d均呈现先升高后下降的变化趋势,与匍匐茎蔗糖含量的变化趋势近似。出苗后5~10 d时SS酶活性迅速增强,在苗后10 d时达峰值,延薯4号SS酶活性增幅最大,费乌瑞它增幅最小,分别是0.46、0.26 mg/(g·min);出苗后10~15 d时SS酶活性下降;出苗后15~30 d时酶活性变化无明显峰谷值,较平缓,且维持在较高水平,出苗后30 d時SS活性有所回升。
2.2.2蔗糖磷酸合成酶(SPS)的活性变化
由图7可以看出,不同马铃薯品种匍匐茎中SPS活性在苗后5~30 d均呈现“上升—下降—上升—下降”的变化趋势。出苗后15 d,3个品种匍匐茎SPS活性均达高峰值,延薯4号SPS活性增幅最大、费乌瑞它增幅最小,分别是0.83、0.50 mg/(g·min);出苗后15~20 d时SPS活性快速下降,克新13降幅最大,费乌[CM(25]瑞它降幅最小,分别是1.57、0.80[KG*3]mg/(g·min)出苗后20~25 d 时SPS活性均小幅度回升,延薯4号回升幅度 [JP3]0.88 mg/(g·min),费乌瑞它回升幅度0.23 mg/(g·min)[JP],克新13回升幅度居于前两者之间;出苗后25~30 d,SPS活[JP3]性小幅度下降,3个品种下降幅度在0.33~0.79 mg/(g·min)[JP]之间。
2.2.3[JP2]转化酶的活性变化
马铃薯匍匐茎转化酶活性变化规律如图8所示。不同品种马铃薯匍匐茎中转化酶活性在出苗后5~30 d呈现“山”字形变化趋势,先降低后升高再降低再升高。[JP3]出苗后5~15 d,转化酶活性逐渐下降,到15 d时出现低谷,克新13号下降幅度0.006 mg/(g·min)、费乌瑞它和延薯4号下降幅度0.003 mg/(g·min);出苗后15~20 d时,转化酶活性[JP2]快速上升,克新13号上升幅度最大,为 0.026 mg/(g·min),费乌瑞它最小,为0.012 mg/(g·min);出苗后20~25 d,转化酶活性快速下降,不同品种下降幅度与15~20 d时上升幅度相近似;出苗后25~30 d,转化酶活性有所回升。[JP]
[TPZGZ8.tif][FK)]
3讨论与结论
糖是植物光合作用的产物,是植物器官组织赖以生长发育的重要能量和物质基础,蔗糖是植物糖类重要组分[23]。从本研究的试验结果看,在匍匐茎生长发育过程中,蔗糖含量影响着其他糖含量的变化[24],可溶性糖含量的高峰值出现时期与蔗糖出现峰值时期近似,蔗糖含量降到最低时还原糖含量达峰值,同时葡萄糖、果糖的含量亦升至峰值。可见匍匐茎发生前期蔗糖是可溶性糖的重要组分,蔗糖可以在基因表达水平上对细胞内的代谢进行调控[25],冯伟等研究表明小麦旗叶可溶性糖、蔗糖含量和灌浆速率随籽粒灌浆进程呈先上升后降低变化趋势[26]。另外,本试验结果与马春梅等[27]、王丽慧等[28]、李墨爱等[29]在大豆、大蒜、南林-895杨苗茎等作物的试验研究与本试验结果一致,生产中应采取相应措施促进蔗糖代谢调控能力,提高结薯率,增加产量。
本试验发现,匍匐茎蔗糖含量与SS酶活性的变化趋势相一致,且随着SS、SPS酶活性升高而增加,尤其SS酶活性达最高时蔗糖积累也处于最高水平,然后随着转化酶活性增强而下降,在20 d时转化酶活性最强,蔗糖不断被分解转化,蔗糖含量下降到较低值,葡萄糖和果糖含量逐渐增加。匍匐茎作为蔗糖的运输器官蔗糖积累较少,它从地上部得到蔗糖后传递到顶端膨大形成的块茎中,以利于块茎的形态建成。这说明在匍匐茎中SS和SPS的活性与蔗糖含量有高度相关性,这与安云蓉[30]、李丹等[10]、苑智华等[31]、周兰兰等[32]的试验结果一致,但SS和SP、转化酶如何协调马铃薯匍匐茎蔗糖代谢还需做进一步的研究。
通过对马铃薯匍匐茎糖代谢规律进行研究,研究结果如下:由于品种间差异,3个品种蔗糖、还原糖、葡萄糖、果糖、可溶性糖含量及糖代谢相关酶活性数值大小、峰顶、峰谷出现时间均有差异,但趋势基本相似。其中,蔗糖、还原糖、可溶性糖均呈现先上升后下降再回升的变化趋势,葡萄糖、果糖呈现先下降后上升再下降又回升的变化趋势,蔗糖合成酶、蔗糖磷酸合成酶活性均呈现先上升后下降再回升的变化趋势,转化酶活性则是呈现“下降—上升—下降—上升”的变化趋势。
[HS2]参考文献:
[1]马光恕,廉华,王彦宏,等. 硼素对马铃薯淀粉合成和积累的影响[J]. 核农学报,2013,27(3):384-390.
[2]吕金庆,许剑平,杨金砖. 黑龙江省马铃薯生产机械化现状及发展趋势[J]. 农机化研究,2009,31(8):239-241.
[3]金荣. 糖对不同磷素水平马铃薯植株的生理调控[D]. 兰州:甘肃农业大学,2013.
[4]马春梅,郭海龙,龚振平,等. 不同基因型大豆糖分积累规律的研究[J]. 作物杂志2011(1):25-29.
[5]小液,戚行江,金伟,等. 杨梅果实糖分积累及其代谢活性变化规律[J]. 浙江农业学报2013,25(1):48-52.
[6]齐红岩,李天来,张洁,等. 番茄果实发育过程中糖的变化与相关酶活性的关系[J]. 园艺学报,2006,33(2):294-299.
[7]段晓艳. 蔗糖诱导马铃薯块茎形成及相关基因表达[D]. 昆明:云南师范大学,2008.
[8]罗玉,李灿辉. 不同糖处理及光周期对马铃薯块茎形成的影响[J]. 红河学院学报,2011(6):94-99.
[9]刘业丽,栾怀海,何琳,等. 大豆可溶性糖积累规律研究[J]. 黑龙江农业科学2014(5):45-48.
[10]李丹,王晔,赫磊,等. 甜菜蔗糖代谢相关酶活性与糖积累关系的研究[J]. 作物杂志,2009(3):27-31.
[11]覃鸿妮,蔡一林,孙海燕,等. 种植密度对不同株型玉米蔗糖代谢和淀粉合成相关酶活性的影响[J]. 中国生态农业学报,2010,18(6):1183-1188.
[12]梁艳荣,胡哓红,姜伟,等. 大葱生长发育过程中可溶性糖、可溶性蛋白质及游离氨基酸含量变化规律的研究[J]. 华北农学报,2007,22(6):119-122.
[13]张明方,李志凌. 高等植物中与蔗糖代谢相关的酶[J]. 植物生理学通讯,2002,38(3):289-295.
[14]张国平,周伟军. 作物栽培学[M]. 杭州:浙江大学出版社,2001:9.
[15]刘克礼,高聚林,张宝林. 马铃薯匍匐茎与块茎建成规律的研究[J]. 中国马铃薯,2003,17(3):151-156.
[16]雷尊国,黄团,邓宽平,等. 不同肥料及施肥方式对马铃薯繁种匍匐茎、块茎形态建成的影响研究[J]. 种子,2013,32(2):78-81.
[17]项洪涛. 三种植物生长调节剂对马铃薯碳代谢生理及产量品质的影响[D]. 大庆:黑龙江八一农垦大学,2013.
[18]王翠松,张红梅,李云峰. 马铃薯块茎发育过程中的影响因子[J]. 中国马铃薯,2003,17(1):29-33。
[19]张万明,王志明,陈开陆,等. 马铃薯淀粉深加工废液可溶性糖含量的测定[J]. 安徽农业科学,2009,37(26):12373-12376.
[20]李合生,孙群,赵世杰,等. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:169-172.
[21]Nielsen T H,Skiarbek H C,Karlsen P.Carbohydrate metabolism during fruit development in sweet pepper (Capsicum annuum) plants[J]. Physiol Plant,1991,82(2):311-319.
[22]Zhu Y J,Komor E,Moore P H. Sucrose accumulation in the sugarcane stem is regulated by the difference between the activities of soluble acid invertase and sucrose phosphate synthase[J]. Plant Physiol,1997,115(2):609-616.[HJ1.65mm]
[23]胡瑞芳,姜慧,李玥瑩. 蔗糖代谢相关酶的研究进展[J]. 北方园艺,2012(1):167-170.
[24]胡瑞芳. 不同品种甜高粱糖分积累规律与糖代谢相关酶关系的研究[D]. 沈阳:沈阳师范大学,2012.
[25]Koch K E,Ying Z,Wu Y,et al. Multiple paths of sugar-sensing and a sugar/oxygen overlap for genes of sucrose and ethanol metabolism[J]. Journal of Experimental Botany,2000,51(343):417-427.[JP]
[26]冯伟,王永华,郭天财,等. 株行距对小麦旗叶蔗糖代谢及产量的影响[J]. 生态学杂志,2009,28(12):2515-2520
[27]马春梅,郭海龙,龚振平,等. 大豆叶片可溶性糖代谢动态变化与相关酶活性的研究[J]. 作物杂志,2012(5):71-75.
[28]王丽慧,方云花,刘青元,等. 大蒜可溶性糖积累与分配特性研究[J]. 浙江农业学报2014,26(4):933-940.
[29]李墨爱,尹增芳,宁代峰,等. NAA处理下南林-895杨无菌苗茎中糖含量及其相关酶活性的变化[J]. 南京林业大学学报(自然科学版),2010,34(2):43-46.
[30]安云蓉. 甜高粱蔗糖合成酶基因的表达规律及其与糖分含量关系的研究[D]. 天津:天津农学院,2013.
[31]苑智华,何秀丽,徐哲,等. 唐菖蒲球茎形成期蔗糖和淀粉代谢及其相关酶活性[J]. 林业科学,2008,44(8):47-51.
[32]周兰兰,栗孟飞,孙萍,等. 酿酒葡萄枝条生长发育过程中蔗糖代谢及相关酶活性的变化[J]. 植物生理学报,2015,51(6):962-968.
姚立志,朱洪文,魏伟,等. 小麦品种苏麦188主要特征特性及高产栽培技术[J]. 江苏农业科学,2017,45(3):59-61.
猜你喜欢
现代养生·下半月(2015年8期)2015-11-16
分析化学(2015年4期)2015-06-08
生态学报(2015年4期)2015-03-10
生态学报(2015年23期)2015-02-11
