
圆尾鲎与中国鲎异速生长研究
2017-04-19 03:24KH3D陈秀荔罗玲娟彭敏杨春玲朱威霖李琼珍罗邦王韶韶刘伟茹李咏梅
西南农业学报 2017年1期
[KH-*3D]陈秀荔,罗玲娟,彭敏,杨春玲,朱威霖,李琼珍,罗邦,王韶韶,刘伟茹,李咏梅**
(1.广西壮族自治区水产科学研究院,广西水产遗传育种与健康养殖重点实验室,广西南宁530021;2.中国科学院遗传与发育生物学研究所,北京100101)
圆尾鲎与中国鲎异速生长研究
[KH-*3D]陈秀荔1,罗玲娟2*,彭敏1,杨春玲1,朱威霖1,李琼珍1,罗邦1,王韶韶1,刘伟茹1,李咏梅1**
(1.广西壮族自治区水产科学研究院,广西水产遗传育种与健康养殖重点实验室,广西南宁530021;2.中国科学院遗传与发育生物学研究所,北京100101)
为了研究鲎形体与体重之间的异速生长关系,进一步探讨其生长特性,分别测定分析圆尾鲎(Carcinoscorpius rotundicauda)和中国鲎(Tachypleus tridentatus)不同年龄段、不同龄期及不同性别的形态参数(头胸甲宽度及体重)。结果表明,圆尾鲎与中国鲎各年龄段、各龄期及不同龄期的雌、雄鲎头胸甲宽度与体重均为极显著正相关,决定系数R2﹥0.81。异速生长曲线斜率-回归系数b(体重的log值与前体宽度的log值)均大于1,总体表现为随着龄期的增长而不断增加,且雌鲎体重相对于头胸甲宽度的增长率大于雄鲎。
中国鲎;圆尾鲎;异速生长;回归系数
中国鲎(Tachypleus tridentatus)和圆尾鲎(Carcinoscorpius rotundicauda)为我国仅有的2种鲎,隶属于节肢动物门(Arthropoda)、有鳌亚门(Chelicerta)、肢口纲(Merostomata)、剑尾目(Xiphosura)、鲎科(Tachypleidae),已有2亿多年的历史,是名副其实的“活化石”(Lamsdell et al,2013)[1]。鲎和其他海洋节肢动物一样蜕皮生长,自受精到孵化要经过4次蜕皮,之后数年内每年蜕皮1~2次才能发育成熟[2]。尽管有关鲎的研究已经进行了一个世纪[3],不同种类的鲎有相同的生活史[4],但有关鲎的寿命及确切的蜕皮次数等问题仍未得到解决[5-6],以及不同物种在不同阶段的生长率数据亟待补充。最好的测定鲎生长的方法就是跟踪调查自然环境下个体从孵化至发育成熟的整个过程[7],但这个方法需要9~12年或者更久[2]。除了追踪已蜕皮个体[5]或是所蜕皮[8-9]的形态指标外,对鲎生长的直接测量也逐步在实验室内展开。由于实验室内人工繁育受精卵至成鲎的难度太大[10],迄今为止关于鲎的连续生长数据还比较少[7-8]。
鉴于自然追踪、人工养殖等直接测量的困难,研究者们尝试通过对鲎不同部分的异速生长研究以期了解其生长发育的目的。异速生长作为生物界中广泛存在的基本规律之一,对比较某种生物或某类生物的部分与整体或不同部分之间的生长差异提供了重要信息[11]。异速生长即生物个体大小与生理生态属性间的非线性数量关系[12],可以通过数学模型来表达,水生生物的异速生长关系通常用体长-体重函数关系式(W=aLb)表示[13],而有关鲎的研究中人们往往用头胸甲最大宽度作为鲎的生长指标,用体重的增加量来衡量鲎的摄食情况[14],而头胸甲宽度与体重的关系就成为衡量鲎健康生长的重要指标[15,16],所以鲎的异速生长关系往往使用头胸甲宽度-体重函数(y=axb)表示[17-18]。通过异速生长分析,即可归纳出鲎某一部分的生长随另一部分或整个生物体的变化而发生的呈幂函数关系的相应改变[13],这种变化不仅反映了鲎的资源分配策略,还为探究鲎的生长与形状、结构与生理之间的关系奠定基础[19]。
1 材料与方法
1.1 试验材料
不同年龄阶段的圆尾鲎和中国鲎均采自于广西合浦县西场镇邻近海域。所有捕获的圆尾鲎先按照年龄大小分别养殖,并用黄色透明的塑料标签(10 mm×5 mm)用超强力胶水标记于鲎的腹甲左侧边缘,分别在4个分离的塑料容器内(30 cm×45 cm× 30 cm)内室温下暂养2~4 d,容器内每日注入盐度28‰的新鲜海水,水深20 cm,同时不间断的曝气,养殖方法参考Lee and Morton的方法[15],测量完成后将所有的鲎放回原生活区域。
1.2 试验方法
1.2.1 形态观察与测量分别对不同年龄阶段圆尾鲎与中国鲎进行形态观察。将不同的鲎洗涤干净并用滤纸擦拭干后进行各项形态指标及体重的测量。通过数显游标卡尺测量鲎的各项形态指标(精确到0.1 mm),通过电子天平称量其体重(精确到0.01 g)。
1.2.2 数据处理利用软件SPSS17.0进行统计分析。分别对圆尾鲎与中国鲎的头胸甲宽度(g)与体重(M)及其他部位间的异速生长关系进行回归分析,获得异速生长曲线的斜率-回归系数b(体重的log值与前体宽度的log值)及截距,并对回归系数进行F检验,P<0.01为差异极显著,P<0.05差异显著,P>0.05差异不显著。
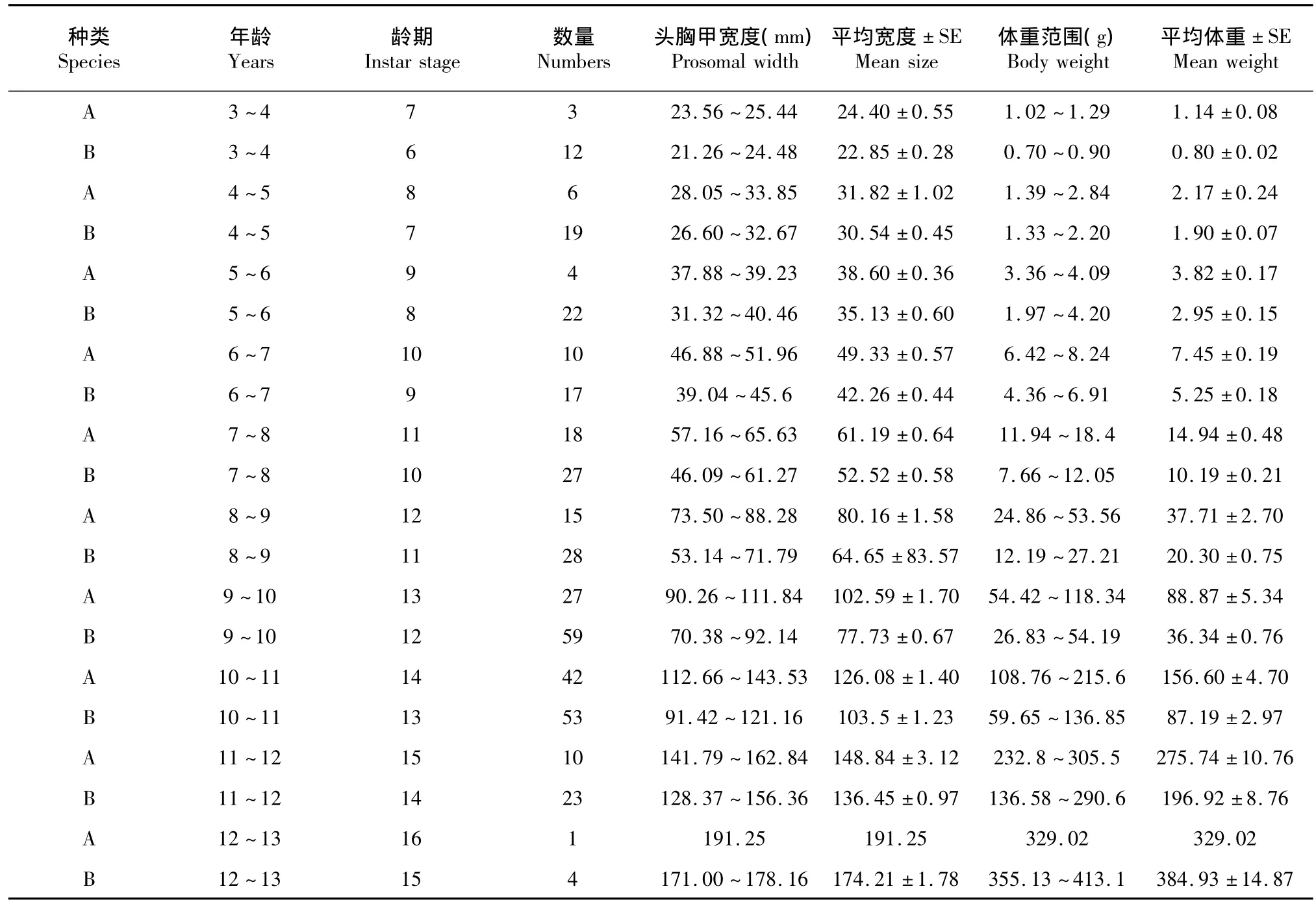
表1 北部湾圆尾鲎和中国鲎头胸甲宽度与体重Table 1Prosomal width(mm)and body weight of C.rotundicauda and T.tridentatus
2 结果与分析
2.1 圆尾鲎与中国鲎头胸甲宽度频率分布
根据头胸甲宽度并参照Sekiguchi[7]和Chiu[19]的方法界定采集样本的年龄,圆尾鲎的年龄为7~16龄,中国鲎的年龄为6~15龄。测定和比较圆尾鲎与中国鲎各龄期头胸甲宽度与体重(表1~2)。表中圆尾鲎与中国鲎分别用A和B表示。
2.2 圆尾鲎头胸甲宽度与体重的关系
2.2.1 不同年龄段圆尾鲎头胸甲宽与体重的关系
分别对不同年龄段(7~9、10~12、13~15、7~16龄)圆尾鲎的头胸甲宽度与体重进行回归分析。体重与前体宽度的log值得出的异速生长方程见表2。
2.2.2 不同龄期圆尾鲎头胸甲宽与体重的关系由于7~9及15龄圆尾鲎样本含量较少,故仅对不同龄期(10、11、12、13、14龄)圆尾鲎的头胸甲宽度与体重进行回归分析,体重的log值与前体宽度的log值得出的异速生长方程见表3。
2.2.3 不同性别圆尾鲎头胸甲宽与体重的关系由于7~9龄及15龄的圆尾鲎样本中雌、雄样本个数小于3,故分别对10~14龄圆尾鲎雌、雄鲎的头胸甲宽度与体重进行回归分析。10~14龄圆尾鲎雌、雄鲎头胸甲宽度与体重平均值与范围见表4。10~14龄圆尾鲎雌、雄鲎头胸甲宽度与体重关系见图3,体重的log值与前体宽度的log值得出的异速生长方程见表5。
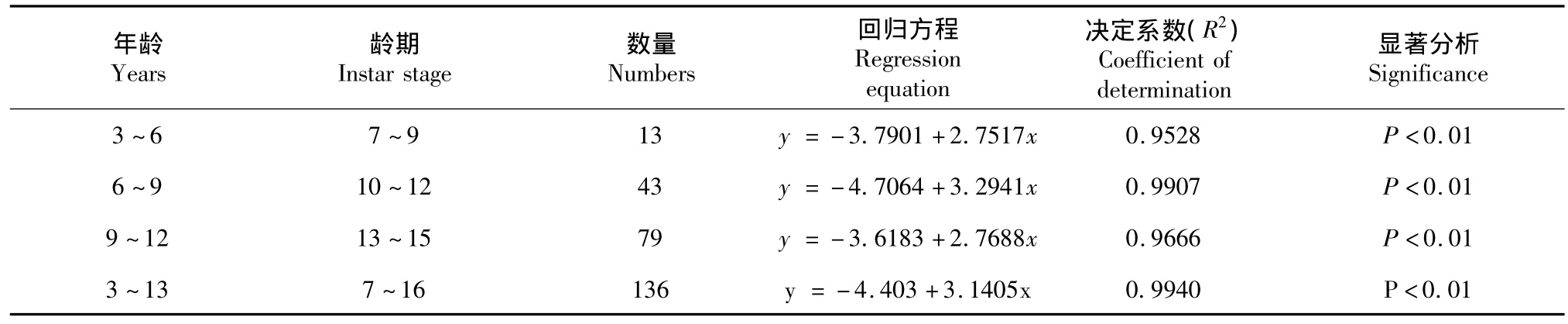
表2 不同年龄段圆尾鲎头胸甲宽度对数与体重对数的关系Table 2C.rotundicauda:Log prosomal width and log body weight relationship of different ages

表3 不同龄期圆尾鲎头胸甲宽度对数与体重对数的关系Table 3C.rotundicauda:Log prosomal width and log body weight relationship of different instars
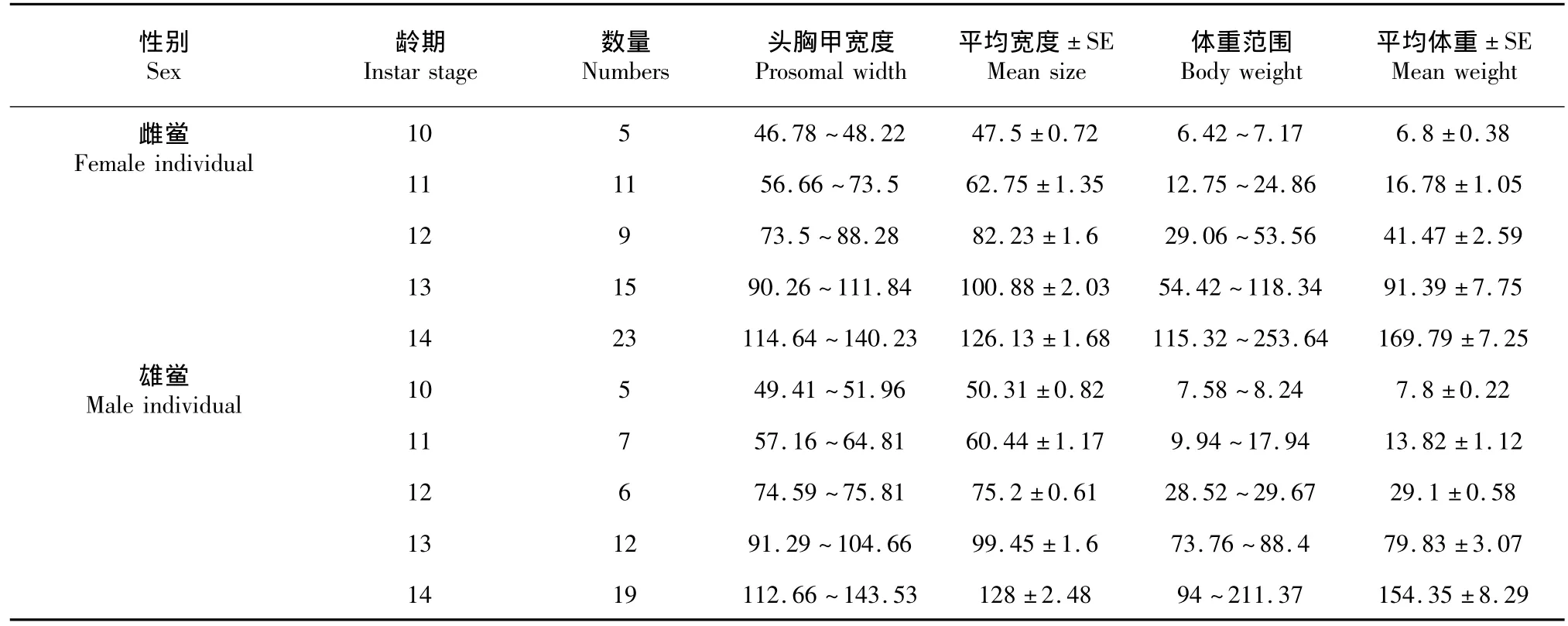
表4 不同性别圆尾鲎头胸甲与体重Table 4C.rotundicauda:prosomal width(mm)and body weight(g)of different sex individuals

表5 不同性别圆尾鲎头胸甲宽度对数与体重对数的关系Table 5Log prosomal width and log body weight relationship of different sex C.rotundicauda
2.3 中国鲎头胸甲宽度与体重的关系
2.3.1 不同年龄段圆尾鲎头胸甲宽度与体重的关系分别对不同年龄段(6~9、10~12、13~15、6~15龄)中国鲎的头胸甲宽度与体重进行回归分析,体重的log值与前体宽度的log值得出的异速生长方程见表6。

表6 不同年龄段中国鲎头胸甲宽度对数与体重对数的关系Table 6T.tridentatus:Log prosomal width and log body weight relationship of different ages
2.3.2 不同龄期中国鲎头胸甲宽度与体重的关系
由于6~9龄及15龄中国鲎样本含量较少,故仅对不同龄期(10、11、12、13、14龄)中国鲎的头胸甲宽度与体重进行回归分析,体重的log值与前体宽度的log值得出的异速生长方程见表7。
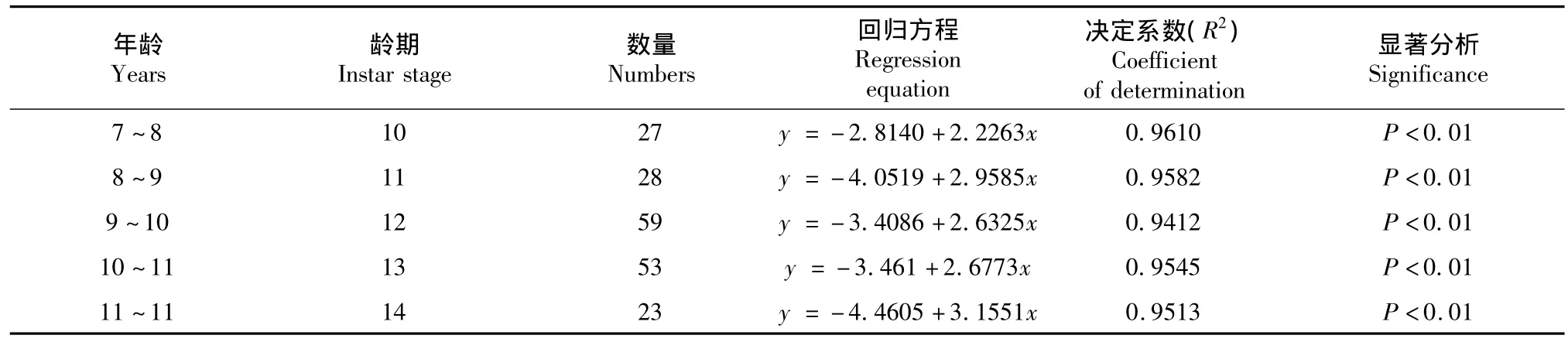
表7 不同龄期中国鲎头胸甲宽度对数与体重对数的关系Table 7T.tridentatus:Log prosomal width and log body weight relationship of different instars
2.3.3 不同性别中国鲎头胸甲宽度与体重的关系
由于6~9及15龄的中国鲎样本中雌、雄样本个数均较小,故仅对分别对10~14龄中国鲎雌、雄鲎的头胸甲宽度与体重进行了回归分析。
体重的log值与前体宽度的log值得出的异速生长方程见表9。
3 讨论
不同年龄段、不同龄期、不同性别圆尾鲎与中国鲎的头胸甲宽度与体重均呈现为正相关,回归系数b值均大于1,且相关系数均在0.90以上,为高度相关(表1~8)。以回归系数评价圆尾鲎与中国鲎的异速生长率,评价指标是当b>1意味着正生长即体重增长率大于头胸甲宽度增长率,反之b<1意味着负生长即体重增长率小于头胸甲宽度增长率。b=1即等速生长[9],而本研究中不同年龄段、不同龄期及不同性别圆尾鲎和中国鲎的b值都大于1,这意味着体重增长率大于头胸甲宽度的增长率,它们处于正生长期。这一结论与不同地区及不同种群的研究结果一致。
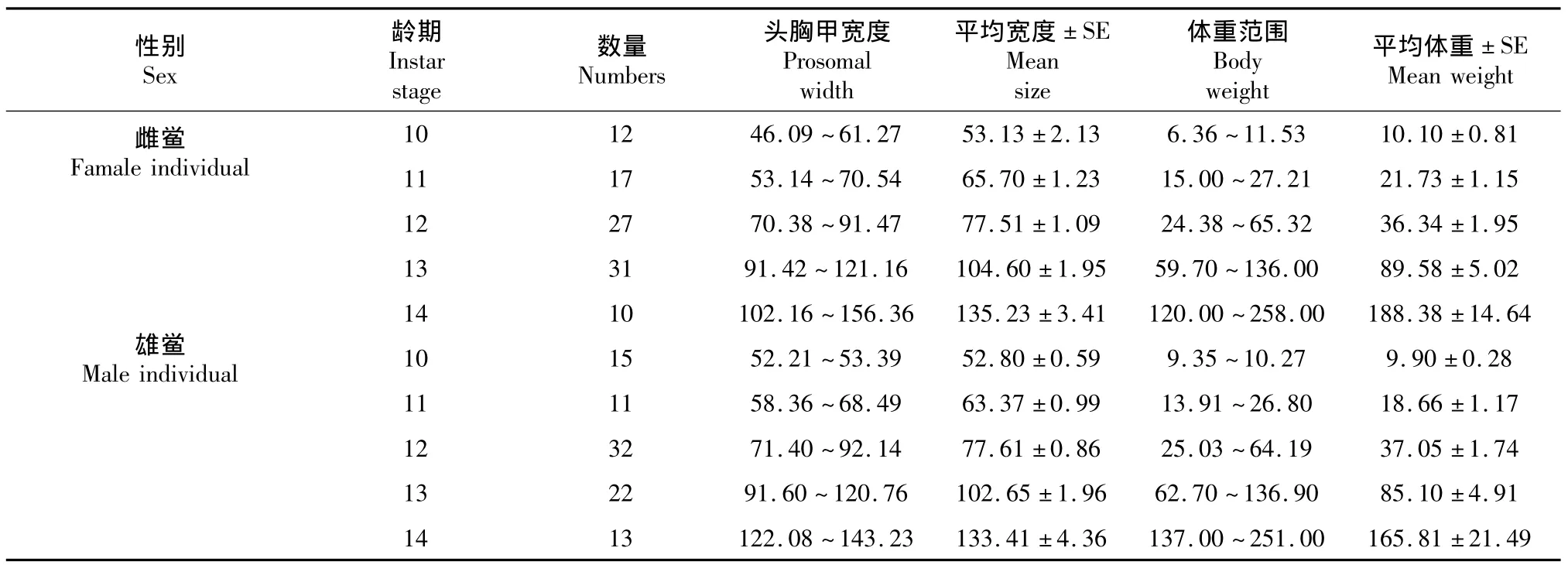
表8 不同性别中国鲎头胸甲与体重Table 8T.tridentatus:Prosomal width(mm)and body weight(g)of different sex individuals
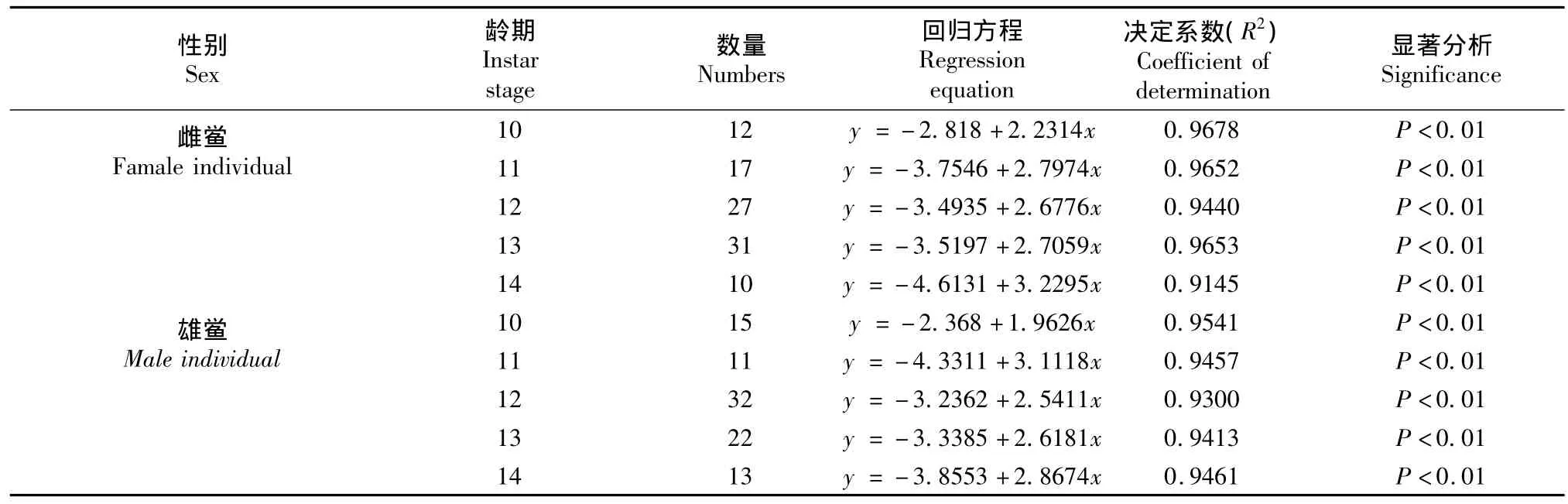
表9 不同性别中国鲎头胸甲宽度对数与体重对数的关系Table 9T.tridentatus:Log prosomal width and log body weight relationship of different sex individuals
其中圆尾鲎与中国鲎不同年龄段回归系数均呈现先升高后降低的趋势。总体评价发现7~16龄圆尾鲎与6~15龄圆尾鲎体重的增长率约为头胸甲宽度的3.1倍。其中7~9龄圆尾鲎与6~9龄中国鲎异速生长系数b分别为2.7517与3.0183,较总体水平偏低,但这一差异在10~12龄年龄段得到补偿,分别达到了3.2941和3.1733,而在接近性成熟时达到平稳。同时Zadeh[20]测得马来西亚1~7龄圆尾鲎头胸甲宽度与体重的b值为2.6727,较本实验中7~9龄圆尾鲎头胸甲宽度与体重回归系数2.7517低,Lee和Morton[15]对从香港沙滩中采样获取的头胸甲宽度在17.1~91.1 mm(6~12龄)之间的中国鲎幼鲎进行人工养殖和体重及头胸甲宽度的测定,发现6~12龄中国鲎头胸甲宽度与体重的回归系数b值为2.9682,较13~15龄中国鲎稍高,如果不考虑地区差异因素,总体符合圆尾鲎体重与头胸甲宽度异速生长率先升高后降低的规律,但实际情况还有待进一步验证。而Chiu[19]对香港圆尾鲎与中国鲎的研究也证实了鲎头胸甲宽度与体重的回归系数会随年龄的增加而变化,并确定了香港中国鲎的回归系数b与年龄间的关系。此外,Zadeh[20]测量1~6龄南方鲎幼鲎头胸甲宽度与体重的回归系数b为2.4845,而暂无有关美洲鲎不同年龄段头胸甲宽度与体重异速生长的研究。
总体观察圆尾鲎与中国鲎各龄期头胸甲宽度与体重异速生长率除11龄外,均表现为随着龄期的增长异速生长率不断升高,暗示随着龄期的增长圆尾鲎与中国鲎变得更宽,体重也不断增加。这与不同年龄段头胸甲宽度与体重异速生长规律相符,也暗示10~12龄年龄段异速生长率的攀高主要是11龄幼鲎体重增长率显著升高造成的。其中10龄圆尾鲎与中国鲎幼鲎头胸甲宽度与体重异速生长率均为2.2左右,在11龄均攀升到2.9,12龄时降低至2.5539和2.6325,而后随着龄期的增长不断升高至14龄达到2.6501和3.1551。这一现象在南方鲎中也有发现,Chatterji[21]报道了南方鲎头胸甲宽度与体重关系随着年龄的增长不断增加的现象,Ismail[16]对马来西亚南方鲎的研究发现随着龄期的增长南方鲎头胸甲宽度与体重异速生长率升高。但由于鲎龄期鉴定较难、同一龄期特别是较小龄期样本含量较小等客观因素,对各龄期异速生长研究较少,而大都偏于对繁殖季节时涌向沙滩产卵的成鲎。
此外本研究还比较了各龄期雌、雄鲎头胸甲宽度与体重的异速生长差异,并发现圆尾鲎与中国鲎10~14龄中除11龄为雄鲎异速生长率大于雌鲎外,其余各龄期均为雌鲎高于雄鲎,且各龄期中国鲎雄鲎头胸甲与体重的生长均优于圆尾鲎雌鲎。这一规律亦存在于其他地区的圆尾鲎、中国鲎及南方鲎中。Chiu和Morton[20]研究测得香港圆尾鲎雌鲎和雄鲎b值分别为2.69和2.00,而中国鲎b值均较低,雌鲎(2.13)和雄鲎(1.89)。同时,Chatterji[21]研究发现印度南方鲎(T.gigas)成鲎雌鲎和雄鲎的b值(截距)2.90(~4.05)和2.55(~3.29),且成鲎体重的增加量大于雌鲎壳长的立方,大于雄鲎壳长的平方。Ismail[16]认为马来西亚珍多海滩南方鲎雌、雄鲎的头胸甲宽度与体重的回归系数b为2.0408和1.1923,珍拉丁海滩南方鲎b值雌鲎为3.0172,雄鲎为1.8474。
同时,回归系数b不仅随着性别的变化而变化,物种、种群及栖息地的不同b值也有差异。本研究发现广西北部湾地区圆尾鲎头胸甲宽度与体重异速生长的回归b值与截距分别为3.1405(~4.403)和3.0934(~4.3007),Morton[22]计算得出香港中国鲎b值为2.48,截距为~3.25。Chatterji[24]对西孟加拉邦鲎的研究表明印度圆尾鲎的b值(截距)为3.89(~0.61)。Vijayakumar等[24]对137只南方鲎进行了形态测量与回归分析,得出体重增长率要高于头胸甲宽度的增长率,相关系数r为0.92。
鲎的种类很少,分布狭隘。由于现存4种鲎的生活环境及地理差异,不同鲎种群之间存在着显著的形态差异,可按照Chatterji[10]分类。其中中国鲎成鲎体型最大(头胸甲宽度约180 mm),圆尾鲎成鲎体型最小(头胸甲宽度约100 mm)[25]。除体型差异外,剑尾是鲎另一个重要的形态特征之一,可凭借剑尾形状对鲎进行大体划分。其中,圆尾鲎剑尾的横截面为圆形内含一个子三角形且三角形中空,其他三种鲎剑尾横截面均为三角形有相似的中空。美洲鲎和圆尾鲎剑尾腹面为凸圆形,但其他两种亚洲鲎剑尾腹面为凹面。此外,南方鲎剑尾顶部边缘有一排小细刺,而中国鲎两边各有一排尖尖的小刺[4]。而我国广西北部湾地区为圆尾鲎与中国鲎的共生区,广西广东沿海地区素有食鲎的习惯,经常错把含有剧毒的圆尾鲎误认为中国鲎,导致食鲎中毒事件发生,为了减少类似中毒事件的发生,我国学者进行了大量有关圆尾鲎与中国鲎的形态学研究[26-29]。鲎血液能提取鲎试剂,还能分离抗革蓝氏阴性及阳性菌、真菌、流感病毒A、口腔泡疹病毒、HIV~1的鲎素类抗菌肤。圆尾鲎含有河豚毒素,中国鲎所含毒素尚不清楚,所以鲎是有待于进一步开发的珍贵海洋药用动物资源。近年来,随着鲎试剂的生产和应用,人们在经济利益的驱使下大肆捕鲎制试剂,鲎资源遭到严重破坏。因此,通过异速生长分析,不仅可以归纳反映鲎的资源分配策略,有助加以保护鲎资源,还为探究鲎的生长与形状、结构与生理之间的关系奠定基础。
[1]Lamsdell J C,Xue J,Selden P A.A horseshoe crab(Arthropoda: Chelicerata:Xiphosura)from the Lower Devonian(Lochkovian)of Yunnan,China[J].Geological Magazine,2012,150(2):367-370.
[2]Shuster Jr C N,Sekiguchi K.Growing up takes about ten years and eighteen stages[J].The American Horseshoe Crab.Harvard University Press,Cambridge,2003:103-132.
[3]Carlos W N,Paul H,Jia-Xing Y,et al.Joint assembly and genetic mapping of the Atlantic horseshoe crab genome reveals ancient whole genome duplication[J].GigaScience,2014,3(9):1-21.
[4]Sekiguchi K.Biology of horseshoe crabs[M].Science House Co Ltd,Tokyo,1988.
[5]Carmichael R H,Rutecki D,Valiela I.Abundance and population structure of the Atlantic horseshoe crab Limulus polyphemus in Pleasant Bay,Cape Cod[J].Marine Ecology Progress Series,2003,246:225-239.
[6]Tanacredi J T.Horseshoe crabs imperiled?[M].In Limulus in the Limelight,Springer US,New York,2001:7-16.
[7]Sekiguchi K,Seshimo H,Sugita H.Post-embryonic development of the horseshoe crab[J].The Biological Bulletin,1988a,174(3): 337-345.
[8]Shuster Jr C N.A pictorial review of the natural history and ecology of the horseshoe crab Limulus polyphemus,with reference to other Limulidae[J].Progress in Clinical and Biological Research,1982,81:1-52.
[9]Shuster Jr C N.Study these,the story of the horseshoe crab[J].Staff Rep,958,10:4-5.
[10]Chatterji A.The Horseshoe Crab:A Living Fossil[M].Project Swarajya,Orissa,1994.
[11]李春萍,李刚,肖春旺.异速生长关系在陆地生态系统生物量估测中的应用[J].世界科技研究与发展,2007,29(2):51-57.
[12]贺纪正,曹鹏,郑袁明.代谢异速生长理论及其在微生物生态学领域的应用[J].生态学报,2013,33(9):2645-2655.
[13]李忠炉.黄渤海小黄鱼,大头鳕和黄鮟鱇种群生物学特征的年际变化[D].青岛:中国科学院海洋研究所,2011.
[14]Schreibman M P,Zarnoch C B.Aquaculture methods and earlygrowth of juvenile horseshoe crabs(Limulus polyphemus)[M].In Biology and Conservation of Horseshoe Crabs,Springer US,New York,2009:501-511.
[15]Christine N.Lee,Brian Morton.Experimentally derived estimates of growth by juvenile Tachypleus tridentatus and Carcinoscorpius rotundicauda(Xiphosura)from nursery beaches in Hong Kong[J].Journal of Experimental Marine Biology and Ecology,2005,318(1):39-49.
[16]Ismail N,Jolly J J,Dzulkiply S K,et al.Carapace Width and Total Body Weight Relationship of Horseshoe crab Tachypleus gigas from Chendor and Cherating,Pahang,Malaysia[J].Empowering Science,Technology and Innovation Towards a Better Tomorrow,2011,6(4):440-444.
[17]Sekiguchi K,Nakamura K,Seshimo H.Morphological variation of a horseshoe crab,Carcinoscorpius rotundicauda,from the Bay of Bengal and the Gulf of Siam[C].In Proc Jap Soc Syst Zool,1978,15:24-30.
[18]Sahu A C,Dey L.Spawning density and morphometric characteristics of the horseshoe crab Tachypleus gigas(Müller)on the Balasore coast of Bay of Bengal,India[J].Sci Vis,2013,13(2):76-84.
[19]Chiu H M C,Morton B.The biology,distribution,and status of horseshoe crabs,Tachypleus tridentatus and Carcinoscorpius rotundicauda(Arthropoda:Chelicerata)in Hong Kong:recommendations for conservation and management[M].The Swire Institute of Marine Science,The University of Hong Kong,Hong Kong,1999.
[20]Zadeh S S,Christianus A,Saad C R,et al.Comparisons in prosomal width and body weight among early instar stages of Malaysian horseshoe crabs,Carcinoscorpius rotundicauda and Tachypleus gigas in the laboratory[M].In Biology and Conservation of Horseshoe Crabs,Springer US,2009:267-274.
[21]Chatterji A.Studies of the biology of some carps[D].Ph.D.Aligarh Muslim University,Aligarh,India,1976.
[22]Morton B,Lee C N W.The Biology and Ecology of Juvenile Horseshoe Crabs Along the Northwestern Coastline of the New Territories,Hong Kong:Prospects and Recommendations for Conservation:Final Report to China Light and Power Company Limited[M].Swire Institute of Marine Science,The University of Hong Kong,Hong Kong,2003.
[23]Chatterji A,Vijayakumar R,Parulekar A H.Growth and morphometric characteristic in the horse-shoe crab,Carcinoscorpius rotundicanda(Latreille)from Canning(West Bengal),India[J].Pakistan Journal of Scientific and Industrial Research,1988,31:352-353.
[24]Vijayakumar R,Das S,Chatterji A,et al.Morphometric characteristics in the horseshoe crab Tachypleus gigas(Arthropoda:Merostomata)[J].Indian Journal of Marine Sciences,2000,29(4):333-335.
[25]Hu M.Aspects of ecology and biology of two sympatric horseshoe crab species(Tacleypleus tridentatus and Carcinoscorpius rotundicauda)[D].City University of Hong Kong,2011.
[26]梁广耀.圆尾鲎(东南亚鲎)Carcinoscorpius rotundicauda在我国北部湾的发现[J].海洋湖沼通报,1986(3):60-62.
[27]孙建运.圆尾鲎在我国的新纪录[J].动物分类学报,1987 (2):9.
[28]陈波,夏昊.东方鲎及其人工繁育技术[J].中国水产,2002(6):33.
[29]廖永岩,李晓梅,洪水根.中国鲎幼体阶段(黄皮鲎)的形态特点[J].动物学报,2002,48(1):93-99.
(责任编辑 温国泉)
Allometric Growth of Carcinoscorpius rotundicauda and Tachypleus tridentatus
CHEN Xiu-li1,LUO Ling-juan2*,PENG Min1,YANG Chun-ling1,ZHU Wei-lin1,LI Qiong-zhen1,LUO Bang1,WANG Shao-shao1,LIU Wei-ru1,LI Yong-mei1**
(1.Guangxi Key Laboratory of Aquatic Genetic Breeding and Healthy Aquaculture,Guangxi Academy of Fishery Sciences,Guangxi Nanning 530021,China;2.Institute of Genetics and Developmental Biology,Chinese Academy of Sciences,Beijing 100101,China)
In order to investigate the relationship between the physique and weight of horseshoe crabs and made a further analysis of its correlation with the allometric growth,the morphometry parameters(carapace width and body weight)of Carcinoscorpius rotundicauda and Tachypleus tridentatus of different ages in both sexes were measured respectively,and their allometric analysis had been carried out at the same time.The result showed that C.rotundicauda and T.tridentatus in different ages,instars and sex between prosomal width and body weight were extremely significant positive correlation,the coefficient of determination R2was greater than 0.81.Allometric growth curve slope-regression coefficient b(relationship between log prosomal width and log body weight)were all greater than 1.Overall performance showed that the carapace width and total body weight were increased with aging of C.rotundicauda and T.tridentatus,and female horseshoe crabs showed better growth than males.
Carcinoscorpius rotundicauda;Tachypleus tridentatus;Allometry;Regression coefficient
Q959.22
A
1001-4829(2017)1-0238-07
10.16213/j.cnki.scjas.2017.1.041
2016-10-08
广西水产遗传育种与健康养殖重点实验室开放课题(GXKL-AQUA-2013-C-03)
陈秀荔(1975-),女,内蒙赤峰人,博士,副研究员,主要从事水产遗传育种研究;*为共同第一作者,罗玲娟(1978-),女,陕西宝鸡市,硕士,工程师,主要从事动物免疫学研究,**为通讯作者:李咏梅,E-mail:Liyongmei915@163.com。
猜你喜欢
湖南城市学院学报(自然科学版)(2022年6期)2022-11-14
昆明医科大学学报(2022年4期)2022-05-23
国际商业技术(2022年4期)2022-04-21
南通职业大学学报(2021年3期)2021-11-17
矿业科学学报(2020年5期)2020-09-18
家庭影院技术(2020年7期)2020-08-24
统计与决策(2018年14期)2018-08-22
江苏农业科学(2017年10期)2017-07-21
江苏农业科学(2017年10期)2017-07-21
中国有色金属学报(2015年6期)2015-03-18
