
基于香蕉叶片解剖结构的抗寒性评价
2017-04-19 03:24KH3D何海旺赵明武鹏龙芳邹瑜
西南农业学报 2017年1期
[KH-3D]何海旺,赵明,武鹏,龙芳,邹瑜
(广西农业科学院生物技术研究所,广西南宁530007)
基于香蕉叶片解剖结构的抗寒性评价
[KH-*3D]何海旺,赵明,武鹏,龙芳,邹瑜*
(广西农业科学院生物技术研究所,广西南宁530007)
利用石蜡切片技术对不同抗寒性的香蕉叶片解剖结构进行研究,为香蕉的抗寒育种提供参考。结果表明,香蕉叶片属典型的异面叶,具有复表皮结构。海绵组织厚度在不同香蕉种质间的变化最大,其次为栅海比。通过聚类分析和相关性分析筛选出海绵组织厚度和栅海比作为香蕉种质抗寒性评价的解剖结构指标。通过隶属函数对18个香蕉种质进行了综合评价,其抗寒性大小顺序为:LT>粉杂>金粉1号>广东粉蕉>HA>LE>LNF>SNY>SNT>NLF>HR>桂蕉青7号>HLT>LNS>SDW>桂蕉6号>ZNF>HLE。通过聚类分析,可以将这些种质分为3类,其中LT、粉杂和金粉1号在供试种质中抗寒性最强。
香蕉;叶片;解剖结构;抗寒性评价
香蕉(Musa spp.)属芭蕉科(Musaceae)芭蕉属(Musa)多年生草本植物[1],可分为香牙蕉(AAA)、大蕉(ABB)、粉蕉(ABB)和龙牙蕉(AAB)4大类。香蕉属热带果树,喜热怕冷[2]。广西虽为全国第二大香蕉产区,但其地处热带亚热带地区,大部分种植区为香蕉种植次适宜区[3],由于近年来寒害发生频繁,严重影响蕉农的收入及种植积极性。2016年1月,在广西大部分香蕉产区出现严重的低温雨雪天气,导致大部分蕉园损失惨重。因此,选育并推广抗寒的香蕉品种对广西香蕉的安全越冬尤为重要。
种质资源的抗寒性评价是香蕉抗寒品种选育的前提,据报道,植物的形态和生理生化等均可作为评价植物抗寒性的指标[4]。但因为植物叶片是植物体暴露在空气中面积最大的器官,长期生活在开放的环境中,其解剖结构容易受到生存环境的影响而发生改变[5],且要观察叶片的解剖结构只需普通的光学显微镜,而不需要繁杂的操作程序和昂贵的仪器。因此,叶片的解剖结构也常被用做植物抗寒性评价的重要指标,叶片解剖结构指标已经在核桃[6]和含笑属[7]等多种植物的抗寒性评价上得到应用。刘星辉[8],刘杜玲[9]研究认为,香蕉叶片的解剖结构与品种的抗寒性密切相关,可作为香蕉抗寒性的鉴定指标,但利用解剖结构指标对香蕉种质进行抗寒性评价却鲜有报道。本文主要针对不同香蕉种质苗期叶片的横切面进行形态学观察,试图筛选出适合香蕉早期抗寒性评价的叶片解剖结构指标并初步评价其抗寒性,为香蕉的抗寒育种提供参考。
1 材料与方法
1.1 材料
从国内各香蕉产区收集18份不同类型的香蕉种质用于抗寒性评价,其编号及名称见表1,所有种质经过组培生根后,于8月份移栽至营养杯中,基质为经杀菌消毒的黄壤。育苗棚为塑料大棚,不盖遮阳网。从移栽到2叶期内,保证空气湿度为95%以上,2叶期后经常浇水保证基质湿润。抽生新叶后,用0.1%~0.5%的复合肥(15-15-15)水溶液进行叶面喷施,每隔一星期施1次。移栽60 d左右,叶龄长至7叶1心期时,取展开的倒数第2张叶片,避开中脉及边缘,切取1cm×1cm小块,用FAA液固定并保存备用。
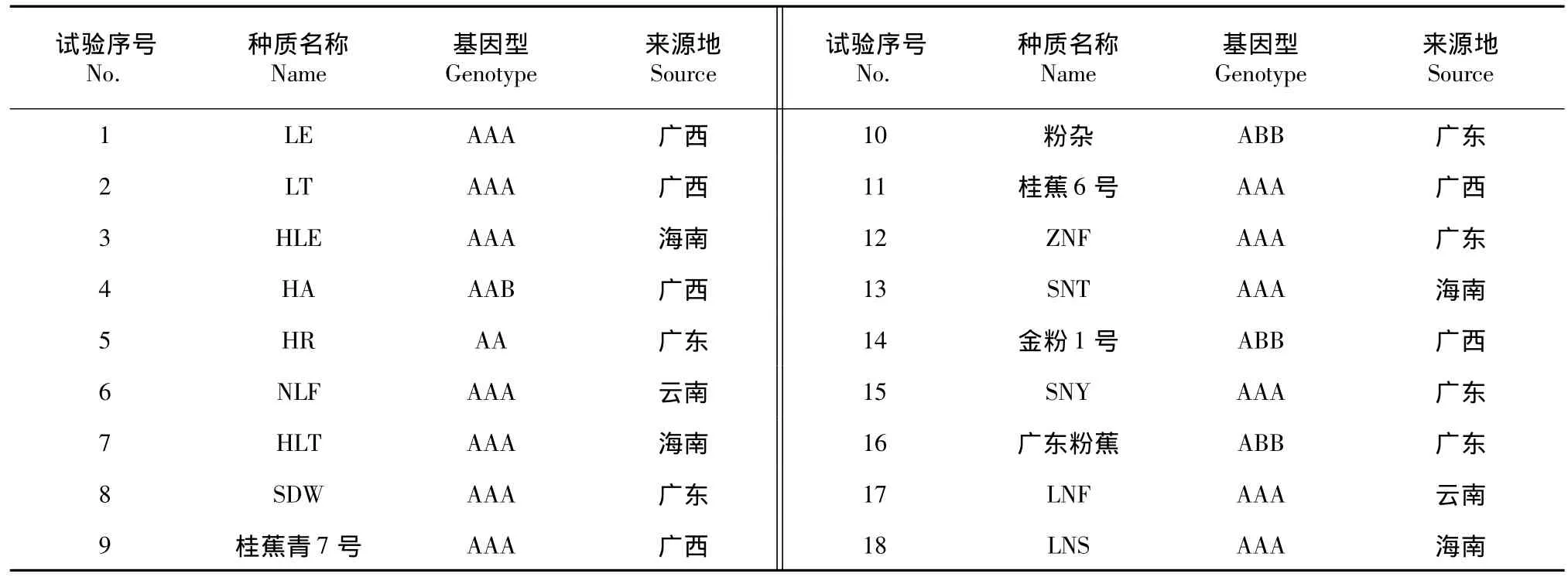
表1 供试验的18份香蕉种质Table 1Test of 18 banana germplasms
1.2 叶片解剖结构观测
采用石蜡切片法[10]对香蕉种质的叶片横切面进行解剖结构观测。
1.3 数据分析
收集的数据采用Excel 2007和SPSS13.0软件进行分析。其中:
细胞结构紧密度(CTR)=栅栏组织厚度/叶片厚度
细胞结构疏松度(SR)=海绵组织厚度/叶片厚度
栅海比=栅栏组织厚度/海绵组织厚度
相关指数:R2i=∑r2/(n-1)
正隶属函数:Uij=(Xij-Xjmin)/(Xjmax-Xjmin)
反隶属函数:Uij=1-(Xij-Xjmin)/(Xjmax-Xjmin)
2 结果与分析
2.1 香蕉叶片横切面的解剖结构
对18份香蕉种质的叶片进行石蜡切片试验,观察其横切面的解剖结构。由图1可以看出,香蕉叶片的上下表皮均由两层细胞组成,外层细胞较小、排列紧密,内层细胞则较大,属于典型的复表皮结构。在显微镜下还能观察到所有的试验材料表皮外均覆盖有角质层。香蕉的叶肉具有明显的栅栏组织和海绵组织,属于异面叶。栅栏组织由两层或多层的细胞组成,细胞长柱状、排列紧密。海绵组织位于栅栏组织和下表皮之间,细胞排列较为疏松。因为供试的材料均来自组培营养杯苗,所以在叶片的中间还存在一个较大的空腔。
2.2 不同香蕉种质叶片解剖特征比较
对供试材料的上表皮角质层(X1)、上表皮(X2)、栅栏组织(X3)、海绵组织(X4)、下表皮(X5)及下表皮角质层(X6)等组织的厚度进行了测量统计。从表2可以看出,香蕉叶片上下表皮的角质层厚度不一样,上表皮角质层平均厚度为1.86 μm,而下表皮角质层的厚度仅为1.54 μm,角质层最厚的是3号种质(HLE),其上、下表皮角质层厚度分别为3.02和1.94 μm。香蕉叶片上表皮的平均厚度(56.00 μm)也明显大于下表皮(36.40 μm),其中,上表皮最厚的是香牙蕉类型的9号种质(桂蕉青7号),而粉蕉类型种质(10、14和16号)的上表皮则相对较薄,14号种质(金粉1号)在供试材料中最薄;下表皮最厚的为3号种质源(HLE),而红香蕉类型的种质LE(1号)则最薄。栅栏组织厚度(29.01~75.88 μm)与上表皮厚度(36.11~71.47)相差不大,平均为54.67 μm,占叶片总厚度的32%,最厚的为10号种质(粉杂)。海绵组织厚度的变异系数为40.49%,在所有测定的指标中最大,不同香蕉种质的海绵组织厚度差异较大,其中最大的为3号种质(HLE),14号种质(金粉1号)则最小。为更合理的利用叶片的解剖特征对香蕉种质进行抗寒性评价,统计出部分与抗寒性密切相关的比值,如CTR (X7)、SR(X8)和栅海比(X9)。在这些比值中,栅海比在不同的香蕉种质中变化最大,变异系数达33.95%。
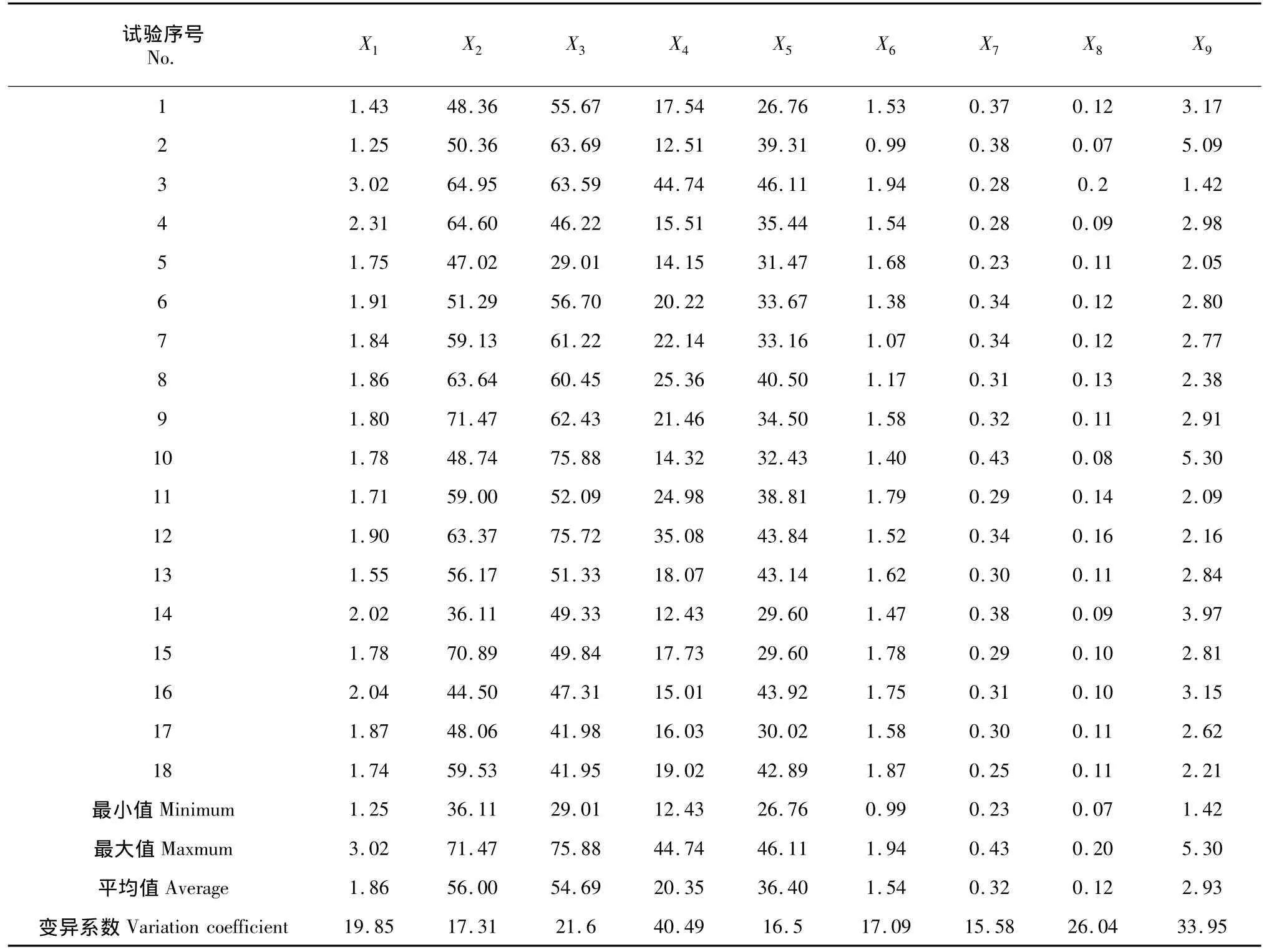
表2 18个香蕉种质的叶片解剖特征比较Table 2Comparison of 18 bananas leaf anatomy characteristics
2.3 叶片抗寒性解剖结构指标的筛选
为合理地利用各项解剖结构指标对香蕉种质进行抗寒性评价,首先需要从多项指标中筛选中有代表性的指标,再以此为依据进行评价。利用离差平方和法(ward's method)对测量统计的9项指标进行系统聚类分析,从聚类结果(图2)可以看出,在虚线A处,9项指标聚成3类,而在虚线B处则聚为2类。当这9项指标聚为2类时,类间的距离比较大,各类的特点突出,相关性小,其中第1类包括X1(上表皮角质层)、X6(下表皮角质层)、X7(CTR)、X8(SR)和X9(栅海比)共5项指标,第2类包括X2(上表皮)、X3(下表皮)、X4(栅栏组织)和X5(海绵组织)共4项指标。
对各项指标进行分类后,利用它们间的相关系数(表3)计算出每个指标的相关指数,在同一类中,相关指数越大的指标,其在所在分类中的代表性就越大。从表4可以看出,在第1类的5个指标中,相关指数最大的为X9(栅海比),达0.4370,确定为该类的典型指标;第2类的4个指标中,X4(海绵组织)的相指数最大(0.2673),确定为该类的典型指标。
2.4 香蕉种质的抗寒性综合评价
根据筛选出的2项香蕉叶片解剖指标(海绵组织厚度和栅海比),应用隶属函数法对18份香蕉种质的抗寒性进行综合评价。因为海绵组织厚度与植物抗寒性呈反相关,所以按反隶属函数公式计算其隶属度[6],而栅海比则与抗寒性呈正相关,所以其属函数按正隶属函数公式计算[11]。按平均隶属度(表5)的大小对18份种质抗寒性进行排序:2(LT)>10(粉杂)>14(金粉1号)>16(广东粉蕉)>4 (HA)>1(LE)>17(LNF)>15(SNY)>13(SNT)>6(NLF)>5(HR)>9(桂蕉青7号)>7(HLT)> 18(LNS)>8(SDW)>11(桂蕉6号)>12(ZNF)>3(HLE)。再以平均隶属度为依据,用离差平方和法(ward's method)进行系统聚类(图3),18个香蕉种质明显聚为3类,第Ⅰ类包括2、10和14号共3份种质,第Ⅱ类包括1、4、5、6、7、9、13、15、16、17和18号共11份种质,第Ⅲ类包括3、8、11和12号共4份种质。根据前人的经验,平均隶属度越大,其抗寒性越高[7],所以这3类香蕉种质的抗寒性大小依次为:Ⅰ>Ⅱ>Ⅲ。

图1 不同抗寒性的香蕉叶片横切面结构图(40×)Fig.1Leaf transection structure of different cold resistance banana(40×)

图2 9项叶片解剖结构指标的聚类分析Fig.2Hierarchical cluster analysis of 9 leaf anatomy structure parameters
3 讨论
据唐立红[12]报道,紫斑牡丹叶片的表皮厚度、角质膜厚度、叶脉突起度和栅栏组织厚度等指标与抗寒性呈正相关,而海绵组织厚度和气孔密度等指标则与抗寒性呈负相关。但也有人认为,表皮厚度、栅栏组织厚度和海绵组织厚度等单一指标,容易受到植物所在的环境及生理状态的影响而发生改变,所以转而采用细胞结构紧密度(CTR)以及细胞结构疏松度(SR)这两个比值来衡量植物的抗寒性,这两个比值相对单一指标更稳定。刘星辉等[8]对福建的3个主栽品种的叶片组织细胞结构进行了观察,认为耐寒性较强的柴蕉的CTR值较其他的高,而SR值则较低,吕庆芳等[9]不同的香蕉品种为试验材料也得出相似的结论。由此可见,CTR与抗寒性成正相关,而SR则与抗寒性成反相关,它们与柑桔的相关系数高达0.8947[13]。杨宁宁等[14]认为叶片的栅海比也与冬油菜的抗寒性呈显著正相关。植物的抗寒性虽然跟叶片多项解剖结构指标存在着密切关系,但如果以所有的指标做为依据评价植物的抗寒性,不仅给分析过程带来困难,而且还会影响到结果的准确性和合理性[7],所以,必须对众多的指标进行筛选。刘杜玲等[11]利用变异系数、相关分析和聚类分析等方法对叶片的解剖结构指标进行了筛选,认为叶片厚度、气孔密度和栅海比可以做为早实核桃品种抗寒性评价的主要叶片解剖结构指标。本文也采用同样的方法对香蕉叶片的8个解剖结构指标进行筛选,认为栅海比和海绵组织厚度可以作为香蕉抗寒性评价的主要指标。
以筛选出的叶片解剖结构指标为依据,采用隶属函数对各个香蕉种质进行了抗寒性评价。根据评价的结果,参试的3个粉蕉品种(粉杂、金粉1号和广东粉蕉)的均排前列,说明它们的抗寒性较强。金粉1号是广西的粉蕉主栽品种,在广西玉林市等香蕉种植次适宜区有大面积种植,较耐霜冻[15],2016年1月的一场罕见寒害中,90%以上的香牙蕉受寒害而干枯死亡,而种于香牙蕉旁边的金粉1号的植株及果实受到寒害的影响则较小,说明粉蕉的抗寒性明显高于香牙蕉。3个粉蕉类型的品种,以粉杂的抗寒性最强,可能与其父母本(从BB型野生香蕉与粉蕉杂交的后代中选育而成[16])具有较强的抗寒性有关。广东粉蕉是在广东收集的一个栽培种,可能因其选育的母本世代繁衍的区域较广西温暖,因此金粉1号的抗寒性比其稍强。前期的人工冷害试验中,也发现HLE在所有种质中的抗寒性是最低的。所以,利用叶片解剖结构指标对香蕉种质的抗寒性进行初步评价,其可信度较高,但仍需进一步的田间以及生理试验进行验证。另外在本次的抗寒性评价中发现LT种质较为特别,LT来自广西收集的一个香牙蕉变异株,但其抗寒性综合评价为最高,与粉杂和金粉1号2个粉蕉品种归类为最抗寒的Ⅰ类。在我们前期的苗期抗寒试验中,也发现其抗寒能力较其他的香牙蕉为强,因此该品种可能是较为抗寒的变异株,可以做为重点考察对象进行进一步的抗寒性鉴定。
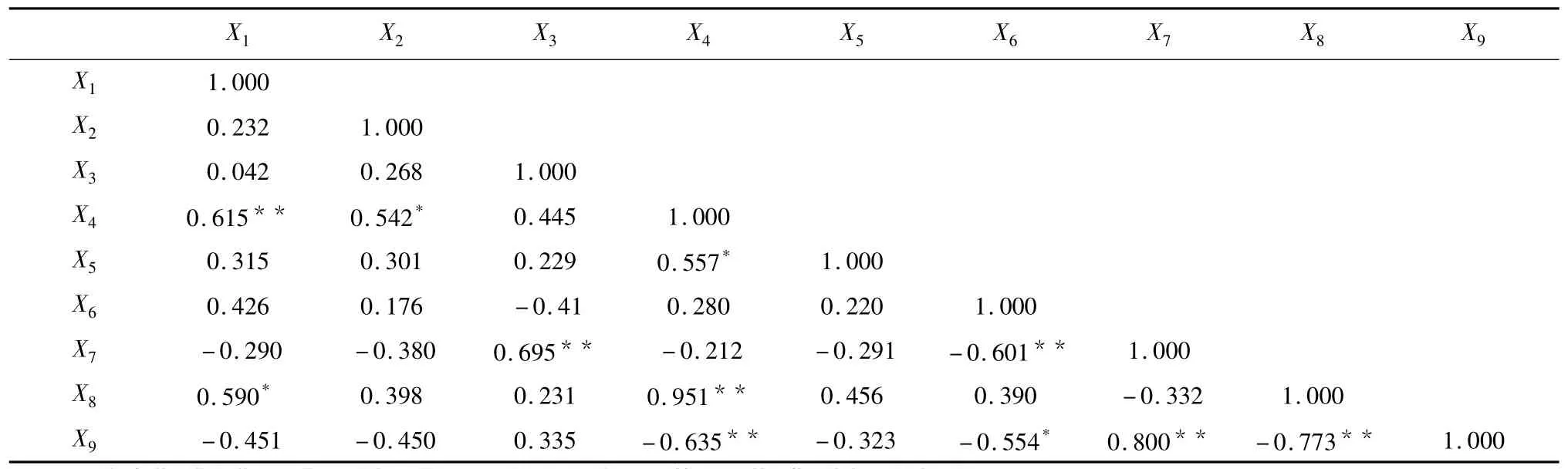
表3 叶片解剖结构指标的相关矩阵Table 3Correlative matrix of leaf anatomy structural parameters
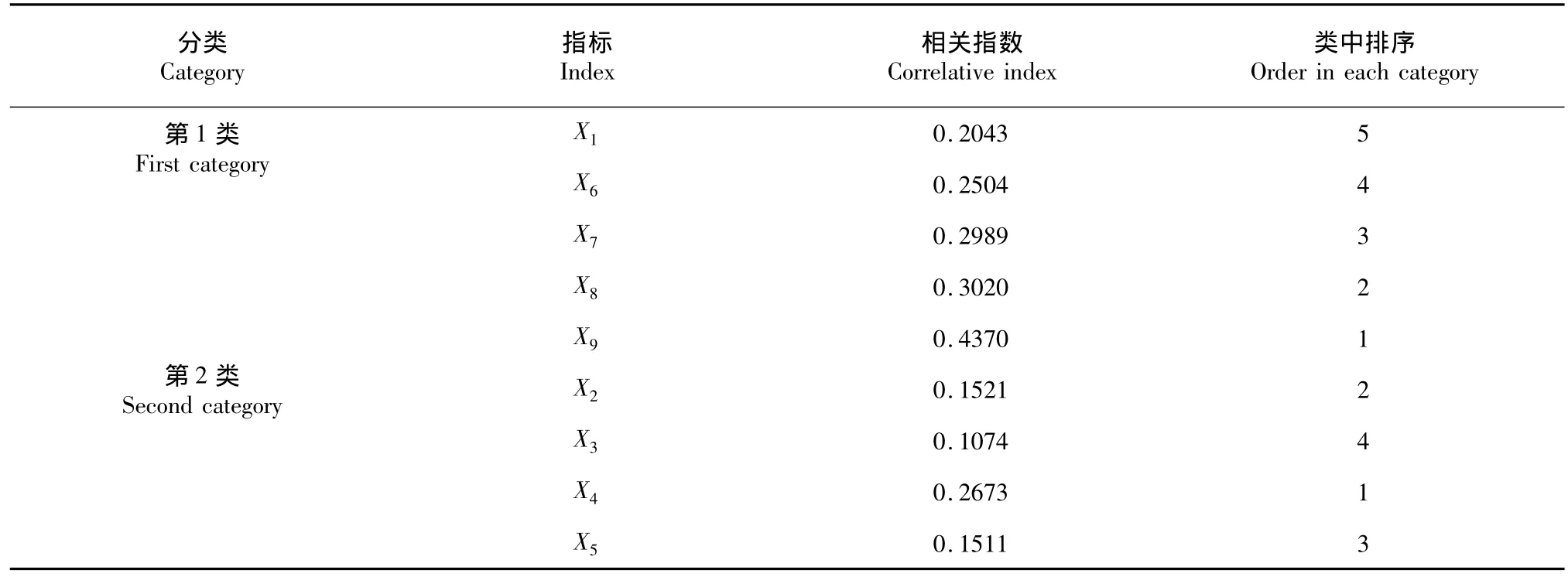
表4 各类中指标相关指数及排序Table 4Correlative index and order in each category

表5 18份香蕉种质的隶属度及抗寒性评价Table 5Membership and cold resistance evaluation of 18 banana germplasms
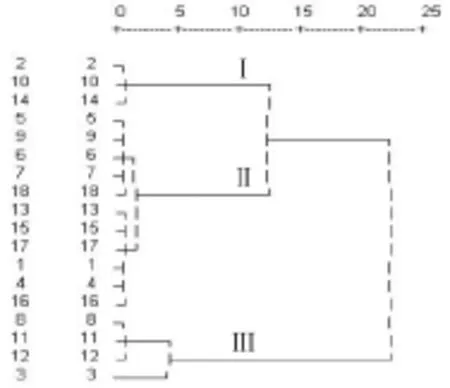
图3 18份香蕉种质抗寒性的系统聚类分析Fig.3Hierarchical cluster analysis on cold resistance of 18 banana
[1]赖瑞联,薛辉康,钟春水,等.闽江流域野生蕉(Musa itinerans)遗传多样性和遗传结构的ISSR分析[J].植物遗传资源学报,2016,17(2):217-225.
[2]孙勇,王丹,仝征,等.香蕉幼苗叶片响应低温胁迫的比较蛋白质组学研究[J].中国农学通报,2015,31(34):216-228.
[3]邹瑜,林贵美,牟海飞,等.广西南宁引种“大丰1号”香蕉品种的研究[J].中国南方果树,2009,38(5):13-14.
[4]姚立新,朱锐,马雯彦,等.植物抗旱、抗寒性鉴定与生理生化机理研究进展[J].安徽农业科学,2009,37(25):11864-11866.
[5]王家训,李春成.武汉地区几种柑桔叶片抗寒性结构特征的研究[J].华中师院学报(自然科学版),1984(3):115-121.
[6]马婷,肖良俊,贺娜,等.12个云南栽培核桃品种叶片解剖结构及其抗寒性综合评价[J].广东农业科学,2016,43(5):2,55-59.
[7]亓白岩,殷云龙,於朝广,等.木兰科含笑属8种植物叶片解剖结构性状与抗寒性的关系[J].江苏农业科学,2013,41(4):150-153.
[8]刘星辉,王宏华,蔡建明,等.香蕉叶片组织细胞结构和生理特性与耐寒性的关系[J].福建农学院学报,1990,19(2):181-185.
[9]吕庆芳,丰锋,张秀枝.香蕉叶片组织细胞结构特性与耐寒性的关系[J].广东海洋大学学报,2000,20(2):48-51.
[10]于晓刚,张文忠,韩亚东,等.粳稻颖果石蜡切片中染色时间的摸索及其解剖结构的观察[J].作物杂志,2010(5):80-83.
[11]刘杜玲,张博勇,彭少兵,等.基于早实核桃不同品种叶片组织结构的抗寒性划分[J].果树学报,2012,29(2):205-211.
[12]唐立红.不同品种紫斑牡丹叶片结构与抗寒性关系的初步研究[J].北方园艺,2010,23:95-97.
[13]简令成,孙德兰,施国雄,等.不同柑桔种类叶片组织的细胞结构与抗寒性的关系[J].园艺学报,1986,13(3):163-168.
[14]杨宁宁,孙万仓,刘自刚,等.北方冬油菜抗寒性的形态与生理机制[J].中国农业科学,2014,47(3):452-461.
[15]邹瑜,林贵美,牟海飞,等.粉蕉新品种“金粉1号”的选育[J].中国南方果树,2011,40(1):47-48.
[16]许林兵,黄秉智,杨护.香蕉品种与栽培彩色图说[M].北京:中国农业出版社,2008:35.
(责任编辑 王冠玉)
Cold Resistance Evaluation Based on Leaf Anatomy Structure of Banana
HE Hai-wang,ZHAO Ming,WU Peng,LONG Fang,ZOU Yu*
(Bio-technology Research Institute,Guangxi Academy of Agricultural Sciences,Guangxi Nanning 530007,China)
The Leaf transection structure of different cold resistance banana have been studied by using paraffin section technology to offer references for breeding of cold resistance of banana.The result showed the banana leaf belonged to typical bifacial leaf with a compound epidermis.Thickness of spongy tissue was the most variable in different bananas,followed by palisade tissue and spongy tissue ratio.Leaf tissue structure indexes were screened by correlation and hierarchical cluster analysis,spongy tissue thickness and palisade tissue and spongy tissue ratio were selected as the anatomy structure indexes to evaluate the cold resistance of banana.Then the cold resistance evaluation of 18 bananas was done by subordinate function,they declined in sequence from LT,Fenza,Jinfen 1,Guangdongfenjiao,HA,LE,LNF,SNY,SNT,NLF,HR,Guijiaoqing 7,HLT,LNS,SDW,Guijiao 6,ZNF,to HLE.Also,these germplasms could be classified into three groups by hierarchical cluster analysis.The cold-resistance of LT,Fenza and Jinfen 1were higher than others.
Banana;Leaf;Anatomy structure;Cold resistance
S668.1
A
1001-4829(2017)1-0193-06
2016-09-01
现代农业产业技术体系项目(CARS-32-15);广西农业科学院基本科研业务专项(桂农科2015YT54,桂农科2017YM29);广西农业科学院科技发展基金项目(桂农科2016JZ03)
何海旺(1979-),男,广西容县人,博士,助理研究员,主要从事香蕉栽培与育种研究,*为通讯作者,E-mail:zy@ gxaas.net。
D OI:10.16213/j.cnki.scjas.2017.1.033
猜你喜欢
热带作物学报(2022年5期)2022-06-01
建材发展导向(2021年14期)2021-08-23
祝您健康·文摘版(2019年4期)2019-06-11
家教世界·V家长(2019年3期)2019-06-11
小天使·三年级语数英综合(2017年11期)2017-12-05
阅读与作文(小学高年级版)(2017年10期)2017-10-11
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
饮食科学(2016年3期)2016-07-04
中国果菜(2016年9期)2016-03-01
中国马铃薯(2015年5期)2016-01-09
