
不同基因型腐烂茎线虫侵染不同甘薯品种后生物学特性研究
2017-04-19 03:23KH3D王宏宝李茹付佑胜赵桂东梁照文张凯
西南农业学报 2017年1期
[KH-*3D]王宏宝,李茹,付佑胜,赵桂东*,梁照文,张凯
(1.江苏徐淮地区淮阴农业科学研究所,江苏淮安223001;2.宜兴出入境检验检疫局,江苏宜兴214206)
不同基因型腐烂茎线虫侵染不同甘薯品种后生物学特性研究
[KH-*3D]王宏宝1,李茹1,付佑胜1,赵桂东1*,梁照文2,张凯1
(1.江苏徐淮地区淮阴农业科学研究所,江苏淮安223001;2.宜兴出入境检验检疫局,江苏宜兴214206)
通过在不同抗性甘薯品种上接种不同基因型腐烂茎线虫来研究线虫群体繁殖量、性比的生物学特性。结果表明,基因型B群体DeYL随着接种数量的增加,线虫在不同甘薯品种下繁殖数量均显著增加,其中在栗子香品种上的繁殖量增加最高,与其它2个甘薯品种相比,线虫繁殖量在(P<0.05)上差异显著;随着接种量的增加,在栗子香上线虫性比从0.77增加到1.44,表现线虫向雌虫方向分化;在徐薯18上同样表现为线虫向雌虫分化,相关性分析发现,在胜利百号、栗子香、徐薯18甘薯品种上,线虫群体繁殖量与性比均呈正相关,相关系数分别为0.987、0.477、0.984;同一接种量下3个线虫群体的繁殖量与性比测定结果显示:3个线虫群体繁殖量、雌雄比在不同甘薯品种下差异显著,证实了线虫生长环境对于线虫的生长发育,性比有一定的影响;3个线虫群体均表现为感病品种栗子香上繁殖量高,雌雄比高,线虫性别偏向雌虫分化。
腐烂茎线虫;营养环境;繁殖量;雌雄比
腐烂茎线虫(Ditylenchus destructor Thome)是一种对植物危害严重的病原线虫。该类线虫最初发现于马铃薯上,可导致马铃薯腐烂,因此国外也称之为Potato nematode[1]。该线虫被亚太植物保护组织、许多国家和地区及我国列为重要的植物检疫性有害生物[2-3]。腐烂茎线虫在中国首先发现于甘薯上,主要侵染块根并使其腐烂,引起甘薯茎线虫病,随后在中药材(如当归、薄荷、人参)以及马铃薯上都分离到该类线虫[4-5]腐烂茎线虫自1937年传入我国北方地区以来,通过甘薯种薯运输、种苗调运、农事操作以及花卉植物的种苗、苗木运输等途径,已扩散到河北、河南、北京、山东、江苏、安徽、吉林、内蒙古等12个省(区)[6-9],并成为北方甘薯产区最严重的病害;受害田块一般可减产20%~40%,严重的可减产80%以上,甚至绝收[10]。腐烂茎线虫除危害甘薯外,还可危害蚕豆、花生、小麦、玉米、山药、胡萝卜萝卜、大蒜等几十种农作物或经济作物。
线虫种群的性别比例对于种群自身的发育繁殖、后代繁衍有重要作用,据报道称,加拿大阿尔伯特大学科学家曾获得了一项十分有趣的发现:营养状况以及种群密度可以改变动物线虫的自然性别,根据他们的研究结果,动物的自然属性以及基因构造却能通过营养状况以及生存环境发生变化[11]。有报道研究表明,昆虫病原索科线虫性别是由营养环境决定的[12],这种机制在动物界极其特殊。那么,对于腐烂茎线虫在中抗、感病甘薯品种下其种群数量、性比否会发生变化国内报道不多。
本试验旨在通过不同抗性甘薯品种接种不同线虫种群,来研究不同线虫种群繁殖量与雌雄比的变化关系及发育情况,试图了解甘薯品种差异对于腐烂茎线虫种群后代群体繁殖量及雌雄比的影响及其相关性变化,该研究对于揭示线虫群体在不同生长环境下雌雄比与群体繁殖量的生物学特性具有重要的科研价值,也为通过外源物影响线虫性别比例来控制影响线虫生长发育、后代繁殖具有重要意义。
1 材料与方法
1.1 腐烂茎线虫群体纯化培养
腐烂茎线虫由南京农业大学提供,为两类基因型A和B[13]线虫群体,线虫培养保存方法,先在解剖镜下挑出生活力较强的线虫100条,用3%H2O2消毒3 min,接种到经乙醇(75%)消毒5 min的健康薯块上,蜡封。25℃培养30 d后,转移到4℃冰箱备用[14-15]。甘薯材料:胜利百号(中抗),徐薯18 (感病),栗子香(感病)。
1.2 薯片培养皿接种法
薯块采用直径13 mm打孔器获得,厚度约1 cm,薯块中心再用5 mm打孔器打出约5 mm深的洞将一定量线虫用移液枪接入,接种薯块放置于1%琼脂糖凝胶培养皿中密封后置于25℃培养箱中培养,接种量根据需要分别接种线虫10,25,50条;培养40 d后浅盘法分离线虫,每个处理重复3次。
1.3 线虫繁殖量和雌雄比测定
通过浅盘法分离到的线虫收集在10 mL试管中,先定下体积,然后充分混匀后,用移液枪吸取50 μl线虫悬浮液滴在玻片上并置于低倍显微镜下计数后统计出线虫的总数,性比(雌雄比)的测定:吸取100条左右的线虫在显微镜高倍镜下记录线虫的雌雄比[16],每个处理重复3次。
1.4 发育进度的测定
以性成熟个体的比例作为发育进度的指标[17],分析线虫在不同甘薯品种下生长发育情况。将上述方法3.2中分离的线虫用移液枪吸取约100条在显微镜下观察并统计性成熟个体的比例。每个处理设3次重复,性成熟百分比=性成熟个体数/总线虫数×100%。
1.5 结果统计与数据处理
应用Spss16.0软件进行差异显著性与相关性分析,旨在发现腐烂茎线虫群体在生物学特性上的相关关系。
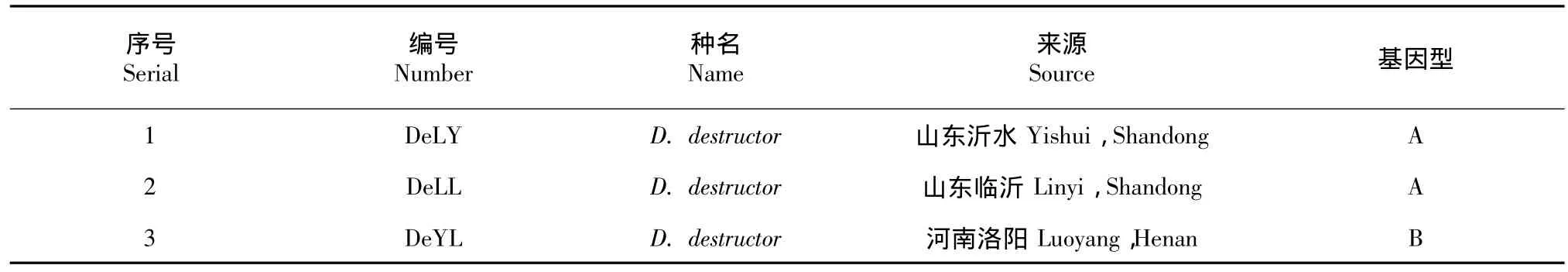
表1 供试腐烂茎线虫编号和来源Table 1Number and origin of tested Ditylenchus destructor
2 结果与分析
2.1 同一线虫群体在不同品种上的繁殖量与雌雄比
表2 数据显示,在不同甘薯品种中,同一接种量下线虫群体的繁殖量与性比不尽相同,证实线虫生长环境对于线虫的生长发育,性比有一定的影响;表现为感病品种栗子香、徐薯18适合线虫的生长繁殖,线虫在这样的环境下利于自身繁殖,表现为繁殖量高,雌雄比高,线虫性别偏向雌虫分化;而在中抗品种胜利百号甘薯品种上,虽然线虫繁殖量远不如2个感病品种多,但随着接种量增加,繁殖量增加,线虫的性比分化也偏向雌虫分化。
线虫群体DeYL随着接种数量的增加,线虫在不同甘薯品种下繁殖数量均有增加,其中在栗子香品种上的繁殖量增加最高,与其它2个甘薯品种相比,线虫繁殖量在P=0.05上差异显著;随着接种量的增加,在栗子香上线虫性比从0.77增加到1.44,表现线虫向雌虫方向分化;在徐薯上随着线虫繁殖量的增加,线虫的性比从0.44增加到2.4,同样在该环境下线虫向雌虫分化。
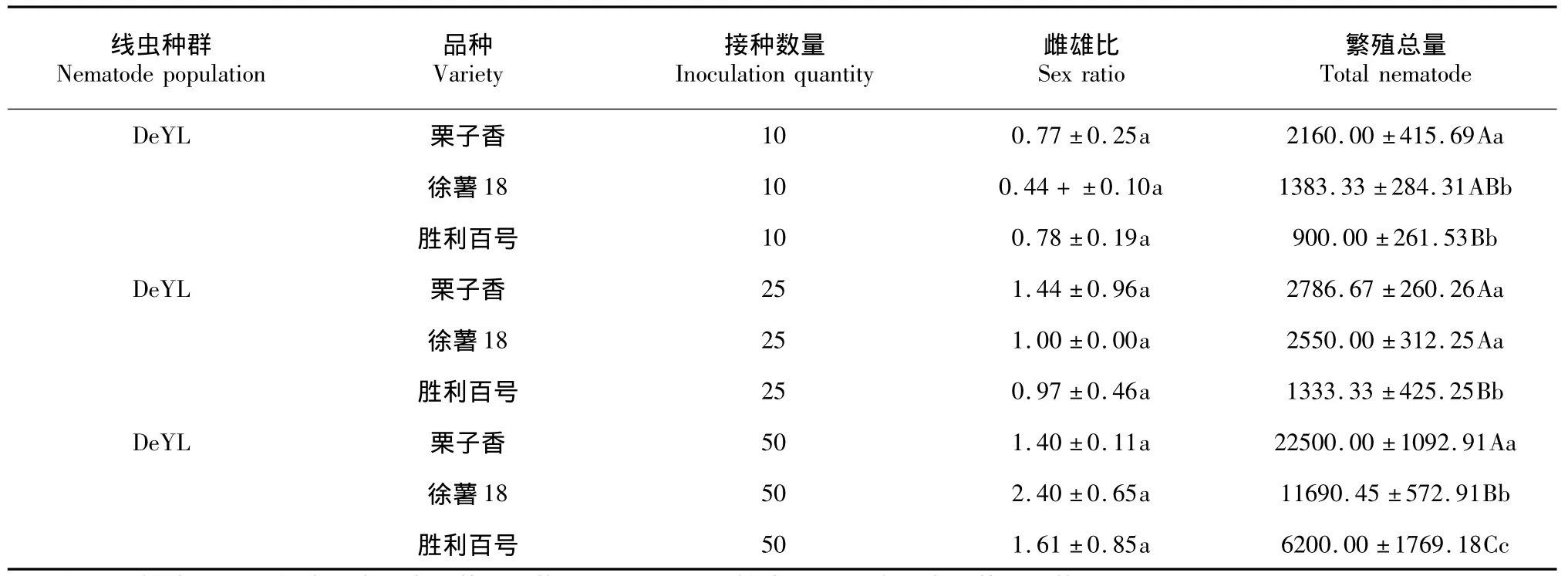
表2 同一线虫群体在不同品种上的繁殖量与雌雄比Table 2Reproductive capacity and sex ratio of DeYL nematode population in different varieties
2.2 同一线虫群体在不同品种上的繁殖量与雌雄比相关性分析
表3 相关性分析发现,在胜利百号、栗子香、徐薯18甘薯品种上,线虫群体繁殖量与性比均呈正相关,相关系数分别为0.987、0.477、0.984。其中,在栗子香上线虫群体繁殖量与性比相关性最低,仅为0.477,分析认为在该品种上线虫量随着提高线虫繁殖量提高远超过其它2个品种,笔者认为在同样甘薯重要的营养环境下,线虫群体达到一定密度后反而会限制线虫向雌虫方向分化,这项发现似乎与Prahlad V等人对秀丽小杆线虫性别与种群数量研究中提到的线虫群体的性别分化与种群密度有联系[11]相关;另外,结论也显示该线虫群体在不同甘薯品种下繁殖量与性比相关性比较,在栗子香上相关性与其它2个品种的相关性差异显著,这从侧面也反映在不同甘薯品种下性比变化差异显著,这一结论同样与加拿大科学家Prahlad V等人的研究发现一致,即周围的环境变化可以影响线虫自身的雌雄性别比例[11]。

表3 不同接种数量下线虫群体数量与雌雄比的相关性Table 3Correlation between the number of nematode population and the ratio of male and female under different inoculation quantities
2.3 同一线虫群体在不同品种上发育进度测定
同一接种量下DeYL线虫群体在不同甘薯品种上繁殖的成虫比例如图1所示,DeYL线虫群体在不同甘薯品种上成虫比例表现不尽相同,DeYL线虫群体在基础接种量为10条的时候,在3个甘薯品种上成虫比例比较接近,在P=0.05上差异不显著;而在基础接种量为25、50条的情况下,在3个甘薯品种上成虫比例在P=0.05上差异显著,当接种量为50条的时候,在胜利百号上的成虫比例显著高于其它2个感病品种,分析认为,在线虫总量增多的情况下,幼虫往往所占比例高,实验发现在徐薯18和栗子香上二龄幼虫占比远高于胜利百号,这样来看在线虫总量多的情况下,成虫占比低也是可以理解的。
2.4 同一接种量下不同线虫群体的繁殖量与性比测定
表4 数据显示,在不同甘薯品种下,同一接种量下不同线虫群体的繁殖量与性比也有差异,其中繁殖量在不同甘薯品种下差异显著;同样证实了线虫生长环境对于线虫的生长发育,性比有一定的影响; 3个线虫群体均表现为感病品种栗子香上繁殖量高,雌雄比高,线虫性别偏向雌虫分化,线虫在这样的环境下利于自身繁殖;其中,基因型B群体DeYL、基因型A群体DeLY线虫群体在感病品种栗子香、徐薯18上的繁殖量比中抗品种的高,这从侧面说明抗性甘薯品种在抗线虫方面表现出不利于线虫增值侵染的优越性;而基因型A群体DeLL在3个不同甘薯品种下均表现出性比向雌虫方向分化,比如在中抗品种上的性比1.56均高于其它2个线虫群体在该品种上的性比,当然这种情况一方面可能是不同线虫群体自身遗传特异性表现出的差异,也或是由于线虫群体密度达到一定量后性比向雌虫方向分化,笔者对于后者可能性比较感兴趣,因为从数据上可以看出,基因型A群体DeLL线虫群体在中抗品种上性比1.56对应的繁殖量为1616.67,而基因型B群体DeYL、基因型A群体DeLY线虫群体在该品种上对应的性比0.97、0.64和繁殖量1333.33、866.67;即不考虑线虫自身遗传差异性外,线虫性比分化与繁殖量有一定关联。
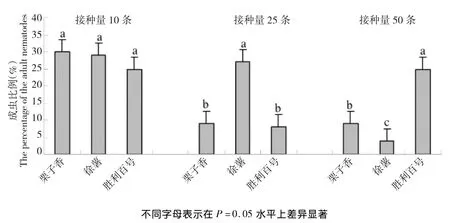
图1 同一线虫群体在不同品种上发育进度测定Fig.1Adult percentage of DeYL nematode group in different varieties of sweet potato
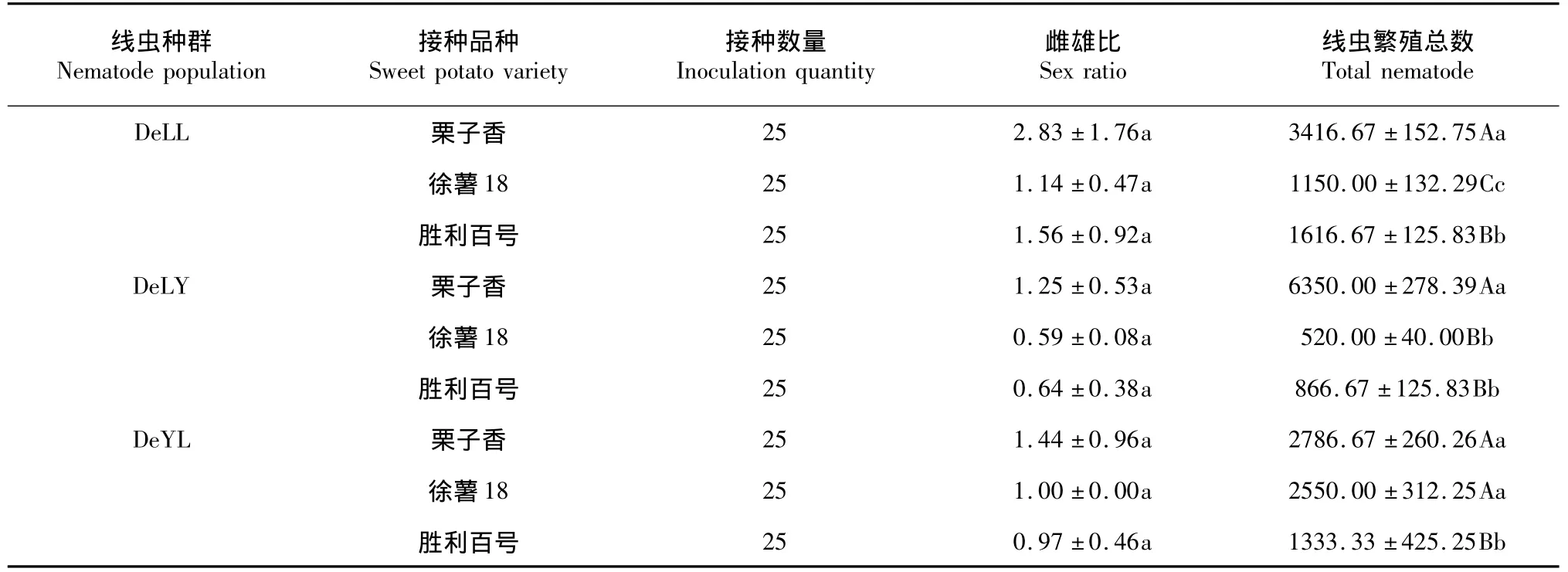
表4 不同线虫群体在不同品种上的繁殖量与雌雄比Table 4Correlation between the number of nematode population and the ratio of male and female in different inoculation quantities
2.5 同一接种量下不同线虫群体的繁殖量与性比的相关性
如果不考虑不同甘薯品种的因素对线虫的影响,单从线虫群体数量上分析群体数量对线虫性别的影响,随着线虫繁殖量的增加,从表5相关性分析中发现,在胜利百号、栗子香、徐薯18甘薯品种上,线虫群体繁殖量与性比均呈正相关,其中DeLL群体、DeLY群体的相关系数分别为0.999、1.000,在P=0.05达显著水平;其中,DeYL群体繁殖量与性比相关性也较高,为0.699。

表5 不同线虫群体繁殖量与雌雄比相关性分析Table 5Correlation analysis of reproduction quantity and ratio of male and female on different nematode populations
2.6 不同线虫群体在不同甘薯品种上的发育进度
如图2所示,不同线虫群体在不同甘薯品种上成虫比例表现存在差异,3个线虫群体的成虫比例在徐薯18和栗子香上占比高于胜利百号,而且在徐薯18上的发育进度在25%~28%,在P=0.05水平上显著高于胜利百号,说明3个线虫群体在感病品种上发育进度好,成虫占比高。
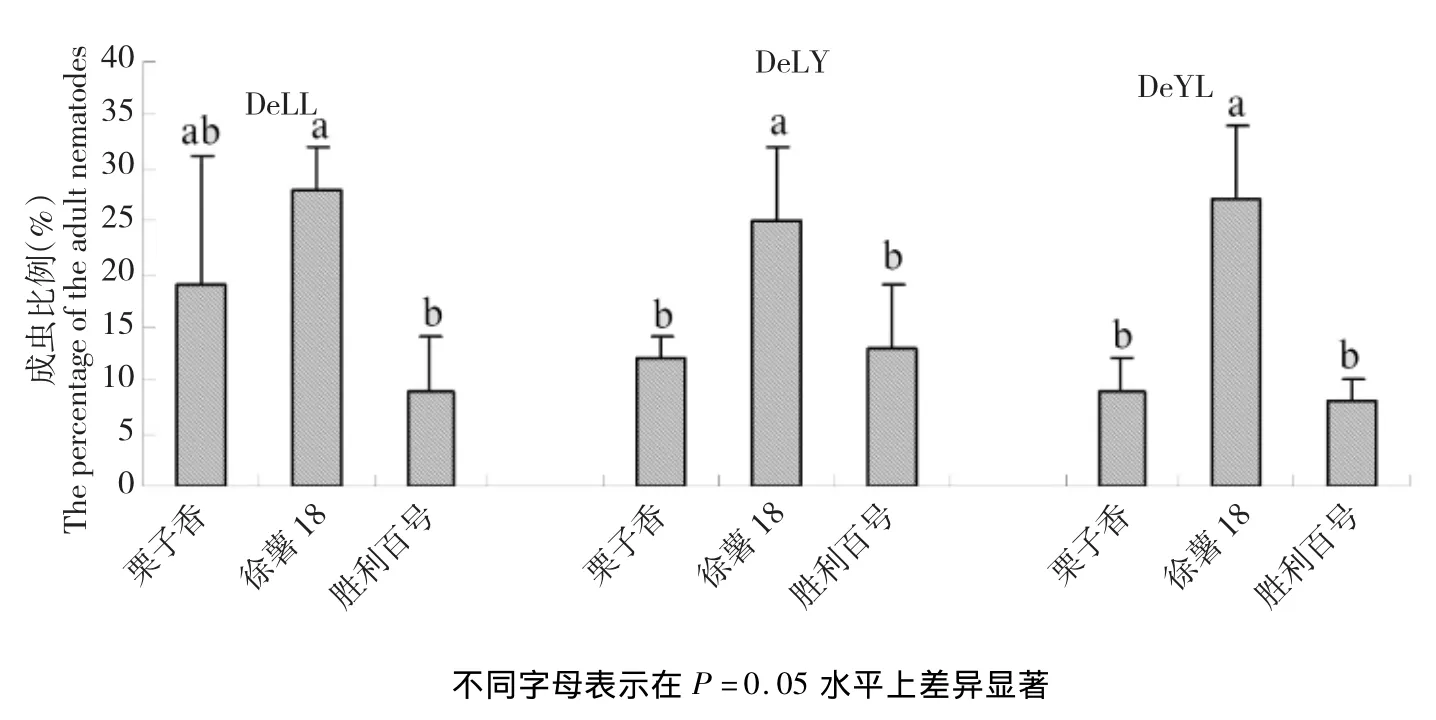
图2 不同线虫群体在不同甘薯品种上成虫百分比Fig.2Adult percentage of different nematode groups in different varieties of sweet potato
3 讨论
在动物家族中,性别的决定方式有多种,但大体可分为两类:基因决定和环境决定。在基因性别决定(GSD)机制中,性别差异由雌雄两性的内在基因不同造成;而在环境性别决定机制中,依赖环境信号的诱导决定动物雌雄。与基因性别决定模型相比,人们对环境性别决定的基因机制还了解得很少。
腐烂茎线虫在不同品种上的繁殖量与雌雄比相关性分析发现,DeYL群体在胜利百号、栗子香、徐薯18甘薯品种上,线虫群体繁殖量与性比均呈正相关,其中,在栗子香上线虫群体繁殖量与性比相关性较低,仅为0.477,分析认为在该品种上随着线虫量提高,线虫繁殖量增加数量远超过其它2个品种,笔者认为在线虫侵染的初期,线虫群体密度低,线虫为了快速稳固自身群体优势,线虫繁殖量与雌雄比存在正相关,即随着种群密度的增加,线虫偏向于雌虫方向分化,或者也可以认为是线虫尽可能向雌虫分化,为的就是多繁育后代增加群体密度;然后,有趣的是,线虫并不是随着群体密度的增加雌雄比一直增加,当线虫量达到一定量后,性比分化会维持在一定范围内,如,DeYL群体在接种25、50条的情况下,繁殖量为16 000,21 600条,然而性比却保持在1.4左右并没有增加,笔者认为在同样甘薯营养环境下,线虫群体达到一定密度后反而会限制线虫向雌虫方向分化,这一结果似乎说明线虫群体性别分化不仅与营养环境有关,也与种群密度有关系;国内有报道称,昆虫病原索科线虫性别是由营养环境决定的[12],这种机制在动物界极其特殊。已有的生物学研究结果表明,中华卵索线虫性别分化是由线虫的感染强度及其宿主体内营养物质的丰度所决定。感染强度越大,每条线虫吸收的营养越少,越有可能发育为雄虫,反之,则为雌虫[18];对于腐烂茎线虫性别分化研究,与Andre P S报道[12]的昆虫病原索科线虫性别是由营养环境决定较为一致。同时这一结论与加拿大科学家Prahlad V等人的研究发现也较为一致,即周围的环境变化可以影响线虫自身的雌雄性别比例[11],这在某种程度上解释动物是如何适应变化着的环境具有重要意义。这种在性别上的分化现象除了线虫自身染色体核型控制外,不同抗性甘薯营养似乎对线虫的性别分化起到了调控作用,至于不同营养条件下寄主的哪些营养成分或信号物质参与了线虫性别的分化有待于进一步研究。
[1]刘维志.植物病原线虫学(1版)[M].北京:中国农业出版社,2000.
[2]Hooper D J.Ditylenchus destructor[M].CIH description of plantparasitic nematodes No.21.CAB International Wallingford,UK,1973.
[3]姚文国,崔茂林.马铃薯有达生物及其检疫[M].北京:中国农业出版社,2001.
[4]丁再福,林茂松.甘薯、马铃薯和薄荷上的茎线虫的鉴定[J].植物保护学报,1982,9(3):169-172.
[5]陈品三,郑经武.当归麻口病致病茎线虫的鉴定研究[J].植物保护,1988,14(6):12-14.
[6]陈品三.甘薯茎线虫病·中国农业作物病虫害(上册)[M].北京:中国农业出版社,1979:460-465.
[7]王志民,赵云蛾.河南省甘薯茎线虫病危害严重[J].植保技术与推广,1994,14(2):40.
[8]范文中,孙淑云,孙艳梅.警惕甘薯茎线虫病流行[J].吉林农业科学,1999,24(4):32.
[9]叶松枝,李跃杰,刘海元.甘薯茎线虫病的发生与防治技术[J].植保技术与推广,1997,17(3):20-21.
[10]周忠,马代夫.甘薯茎线虫病的研究现状和展望[J].杂粮作物,2003,23(5):288-290.
[11]Prahlad V,Pilgrim D,Goodwin E B.Roles for mating and environment in C.elegans sex determination[J].Science,2003,302 (5647):1046-1049.
[12]Andre P S.Evolution of the control of sexual identity in nematodes[J].Seminars in Cell&.Developmental Biology,2007,18(3):362-370.
[13]王金成,季镭,黄国明,等.腐烂茎线虫不同地理种群ITS区序列比对及系统发育[J].河北农业大学学报,2007,30(5):79-83.
[14]林茂松,贺丽敏,文玲.甘薯形态结构对茎线虫病的抗性机制[J].中国农业科学,1996,29(3):8-12.
[15]林茂松.室内人工接种测定甘薯品种对马铃薯腐烂线虫的抗性[J].南京农业大学学报,1989,12(3):44-47.
[16]王宏宝,戚龙君,王金成,等.马铃薯腐烂线虫不同群体同工酶表型与致病力研究[J].浙江大学学报(农业与生命科学版),2009,35(4):425-432.
[17]黄小娟.化学胁迫下松材线虫的繁殖与交配行为[D].浙江农林大学,2014.
[18]Zhou Y F,Shi Y X,Xie X W,et a1.Leaf spot of spinach caused by Stemphylium spinaciae,sp.Nov.[J].Mycosystema,2011,30(3): 379-383.
(责任编辑 陈虹)
Study on Biological Characteristics of Different Genotypes Ditylenchus destructor Infected on Different Sweet Potato Varieties
WANG Hong-bao1,LI Ru1,FU You-sheng1,ZHAO Gui-dong1*,ZHAO Wen-liang2,ZHANG Kai1
(1.Jiangsu Institute of Agricultural Sciences Xuhuai Region Huaiyin,Jiangsu Huaian 223001,China;2.Yixing Entry-exit Inspection and Quarantine Bureau of People’s Republic of China,Jiangsu Yixing 214206,China)
The biological characteristics of different genotypes Ditylenchus destructor infected on the resistant and susceptible sweet by inoculation of different numbers of Ditylenchus destructor from different area in China were studied.The result showed that with the increase of DeYL,nematode population were significant increased in different varieties,and compared with the other 2 varieties,the amount of nematode propagation in Lizixiang sweet potato varieties were significantly different at 0.05 level;With the increase of inoculation rate in Lizixiang,sex ratio of nematode increased from 0.77 to 1.44 and showed the differentiation of female nematode;In Xushu-18 also shown as to the differentiation of female nematode,the correlation analysis showed that the fecundity and the ratio of nematode populations were positively correlated with three varieties of sweet potato,and their correlation coefficients were 0.987,0.477 and 0.984,respectively;Under the same quantity,the production and sex ratio of three different nematodes indicated that the different populations of nematodes had significant differences in different nutrition conditions,which was proved that the growth environment of the nematode had a certain effect on the growth and development of nematode;Three different nematode populations performed high production,high sex ratio and gender to female differentiation on Lizixiang sweet potato varieties.
Ditylenchus destructor;Nutrient environment;Production;Male and female ratio
S432.45
A
1001-4829(2017)1-0093-06
10.16213/j.cnki.scjas.2017.1.017
2016-01-20
江苏省农业科技自主创新资金项目[SCX(14) 5086];淮安市农科院院长基金(NKY20150518)
王宏宝(1982-),男,山西大同人,硕士,助理研究员,从事植物病原线虫防治与新技术开发,E-mail:wanghongbao252@126.com,*为通讯作者:赵桂东,研究员,主要从事植物病原线虫的生物防治研究。
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
广西林业科学(2022年1期)2022-03-03
山西林业(2021年2期)2021-07-21
环境昆虫学报(2020年5期)2020-11-11
读友·少年文学(清雅版)(2018年12期)2018-04-04
文学少年(有声彩绘)(2018年1期)2018-02-26
读写算·小学中年级版(2017年2期)2017-02-28
Coco薇(2015年12期)2015-12-10
海峡姐妹(2015年3期)2015-02-27
江苏农业科学(2014年3期)2014-07-16
