
野大豆和花蔺的分布和群落特征及就地保护对策
2017-04-14 09:14:20朱思雨
生物学杂志 2017年2期
朱思雨, 宗 雪, 张 玲, 李 静, 雷 霆
(1. 中国野生动物保护协会, 北京 100714; 2. 国家林业局调查规划设计院, 北京 100714;3. 北京市土地权籍事务中心, 北京 100029; 4. 北京林业大学 自然保护区学院, 北京 100083)
野大豆和花蔺的分布和群落特征及就地保护对策
朱思雨1, 宗 雪2, 张 玲1, 李 静3, 雷 霆4
(1. 中国野生动物保护协会, 北京 100714; 2. 国家林业局调查规划设计院, 北京 100714;3. 北京市土地权籍事务中心, 北京 100029; 4. 北京林业大学 自然保护区学院, 北京 100083)
北京郊区湿地类型单一、受人为活动干扰较大,一些植物因此数量减少或分布区萎缩,成为濒危物种。通过对北京湿地野大豆和花蔺的分布特征和种间联接的分析,针对其就地保护工作提出一些可供参考的措施和建议。通过样带结合样方法,调查并记录样方内植物的种名、平均高度、盖度等,计算样方内野大豆和花蔺的重要值,以重要值为基础,对北京湿地2003—2005年和2007—2009年野大豆和花蔺的重要值变化进行比较及方差分析,分析其差异及显著性。通过联结系数AC法和卡方检验法对野大豆和花蔺群落进行种间联结的测定,分析这两个物种与群落中其他物种间的联结性。得出群落中蒙古蒿、三叶鬼针草等和野大豆为正联结,狼把草、鬼针草及浮萍与野大豆是负相关,泽泻、金鱼藻、马来眼子菜等与花蔺间为正联结,且关联程度较高的正联结物种在χ2检验中均是极显著关联,表现出与χ2检验一致性。得出北京湿地野大豆和花蔺面临诸多干扰威胁,如生长环境退化等,针对这些问题,提出改善水质、恢复退化湿地、加强伴生种的保护、控制竞争种的数量等可行性对策,以供参考。
野大豆;花蔺;群落;种间关联;就地保护
北京郊区湿地由于特殊的生境位置导致其类型较为单一,并且受人为活动干扰较大。因此,一些植物数量急剧减少或分布区迅速萎缩,成为濒危物种[1]。野大豆(GlycinesojaSieb. et Zucc.),又称乌豆,豆科,系一年生蔓生缠绕草本[2]。据现有资料报道,野大豆在世界范围内分布相对较为狭窄,仅存在于亚洲东部的中国、日本、朝鲜、俄罗斯远东地区[3]。但是其在我国分布范围广泛,是国家二级重点保护野生植物[4],除新疆、宁夏、青海、西藏外,几乎遍布全国,在北京各个湿地都有记录,尤其在一些湿地的半干燥区域或退化湿地中生长非常茂盛,主要分布在溪流旁、潮湿山坡、河漫滩、河边小树林及苇塘[5]。野大豆是大豆属中唯一能和栽培种Glycinemax杂交的举世瞩目的大豆种质资源,具有高抗逆性和耐旱、耐热等优势,其根系发达且具有根瘤,固氮活性较高,是原产我国的重要作物[6-7]。近年来由于大规模的开荒、放牧、农田改造等原因,致使野大豆自然分布区日益缩减。因此非常有必要对野大豆种质资源进行有效保护。
花蔺(Butomusumbellatus),又名蒲子莲,花蔺科[8]。主要分布在北美洲、欧洲北部地区、亚洲北部地区,是欧亚大陆北温带的广布种。在我国主要集中分布在东北、华北、西北、华中、华东部分省区,尤其在东北、华东及华北等长江以北地区广为分布,但现存种群数量不多[9]。花蔺是北京市二级保护野生植物,喜水,在湖泊、水塘、沟渠的浅水中或沼泽中都能寻见它的踪迹。花蔺科植物北京仅此一种,种群数量不多,较少见,生于湖畔、河畔、沟边、沼泽,以及湖泊、河流、水沟浅水中,是比较珍稀的湿地植物。宿敏在针对北京市的怀柔、延庆等18个区县的重点野生植物分布情况进行调查中,发现花蔺主要分布在海淀西山、吕平南口、延庆松山、永宁镇、怀柔喇叭沟门、怀柔镇、桥梓镇、大兴六合庄林场、顺义汉石桥等地,较为全面地调查了北京地区花蔺的实际分布,但是针对一些典型湿地的花蔺分布情况没有详细涉及[7]。花蔺根茎含淀粉37%~40%,酿造60度酒的出酒率达24%~26%;花美丽,可栽培供观赏;叶可作编织及造纸原料[10]。花蔺科植物北京仅此一种,种群数量不多,是比较珍稀的湿地植物,应该加强保护。
就地保护是保护湿地生物多样性最有效的手段之一。北京湿地具有很高的经济、社会和生态价值,但由于人类的活动等因素,目前已遭到一定程度的破坏。本文根据对野大豆和花蔺的分布及群落特征的研究分析,对其所面临的问题进行探讨,并查阅与就地保护相关的资料,确定相应的就地保护对策。
1 研究方法
1.1 研究区及物种概况
北京市位于华北平原的北部。地理坐标为39°28′~41°05′N、115°25′~117°30′E。北京地区地层发育比较完全,地貌具体由西部山地、北部山地和东南部平原三大地貌单元组成,地势由西北向东南倾斜,主要包括侵蚀构造山地、剥蚀构造台地、丘陵和堆积构造平原等类型。北京地区的植被类型复杂,其中湿地植被以草丛沼泽和浅水植物群落为主,多分布在河流及库塘湿地沿岸滩涂或浅水中。湿地维管束植物中以菊科(Compositae)、禾本科(Gramineae)和豆科(Leguminosae)植物的种类最多,其次是莎草科(Cyperaceae)、唇形科(Labiata)和蓼科(Polygonaceae),区系成分以北温带和世界广布成分为主[8]。
选取汤河、永定河、潮河、汉石桥水库、金牛湖、玉渡山水库、怀九河、怀沙河、怀柔水库以及密云水库10个典型湿地,采用样带法结合样方法,沿着湿地边缘等距离设置采样点,每个采样点设置4~8条样带,样带均垂直于水路交界线,每条样带由4~6个样方组成。调查并记录样方内植物的种名、株数、平均高度及盖度等。根据调查区域内湿地植物的生活型和种类组成,从低到高依次划分为:1)沉水植物层;2)漂浮植物+浮叶植物层;3)垫状植物+蔓生植物层;4)均高50 cm以下的根状茎草+丛生草+挺水植物+直立草本+藤本+寄生植物层;5)均高50 cm以上的高大丛生草+高大挺水植物+高大直立草本+灌木+乔木层,分别命名为沉水层、水面层、地面层、低草层及高草层。
1.3 数据分析
重要值是某个种在群落中的地位和作用的综合数量指标,表示物种在群落中的优势程度。因其简单、明确,因此近年来普遍采用[11-12]。计算公式如下:
(1)
(2)
(3)
(4)
注:Dr为相对密度,D为某个种的株数;Pr为相对盖度,P为某个种的盖度;Fr为相对频度,F为某个种的频度;Iv为重要值。
计算各个湿地每处取样点野大豆和花蔺的重要值,以2003—2005年和2007—2009年两个时间段分别分析野大豆和花蔺的分布情况以及在湿地中的优势度,然后将两个时间段进行比较分析。将各个湿地每处取样点野大豆和花蔺的重要值,以2003—2005年、2007—2009年(花蔺为2007年)两个阶段,利用SPSS进行方差分析,研究这两个阶段内野大豆和花蔺重要值的差异及显著性。采用联结系数AC法来表征种间关联度,卡方检验法进行检验,将样地调查的数据排成2×2列联表,用a表示两个种均出现的样方数,b、c表示两个种分别出现的样方数,d表示两个种均不出现的样方数,N为取样总数(表1)。

表1 物种出现频次列联表
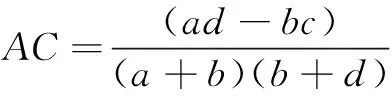
(5)
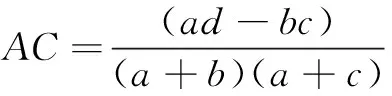
(6)

为进一步提升土壤修护能力,解决经销商“知病不会看病”等问题,2017年,瑞丰生态成立了土壤修护研究院,凭借中国农科院国家测土配方实验室及众多专家学者的技术支持,为农民提供土壤健康优化方案、作物营养管理施肥套餐、作物全程健康植保方案及作物效益提升服务,实现了作物生长过程的安全可追溯,帮助农民实现了增产增收。同时,通过在基层各地设立土壤修护工作站,开展土壤数据分析和为农服务等工作,瑞丰生态土壤修护研究院正在致力于打造中国规模最大的基层土壤修护服务体系。
(7)
AC的值域为[-1,1]。AC值越趋近于1,表明物种间的正联结性越强;AC值越趋近于-1,表明物种间的负联结性越强;AC值为0,物种间完全独立[11-12]。根据2×2列联表,用卡方检验法检验其联结强度:
(8)
若χ2<3.841时,则认为两个种独立分布,即中性联结;若3.841<χ2<6.635时,则认为种间联结显著;若χ2>6.635时,则认为种间联结极显著。
2 结果与分析
2.1 北京湿地野大豆和花蔺优势度年际变化
将野大豆在2003—2005年和2007—2009年两个阶段在北京湿地的重要值进行汇总后发现(如表2):汤河、怀沙河、怀柔水库3处湿地,野大豆的重要值2003—2009年的变化幅度不大;而其余7处湿地野大豆重要值的变化较大,其中潮河、密云水库2003—2005年的野大豆重要值远远比2007—2009年的大,而永定河、汉石桥水库、金牛湖、玉渡山水库和怀九河野大豆的重要值变化则相反,其2007—2009年的值反而大于2003—2005年。
在上述10处典型湿地的调查中,仅在金牛湖、怀柔水库及怀沙河3处湿地的样方内发现了花蔺。因此,将花蔺在金牛湖和怀柔水库2003—2005年与2007年的重要值进行比较(如表2)可知:怀柔水库花蔺的重要值变化较大,其在2007年的重要值远远大于2003—2005年;而花蔺在金牛湖的重要值变化虽然不大,但也是有所降低。
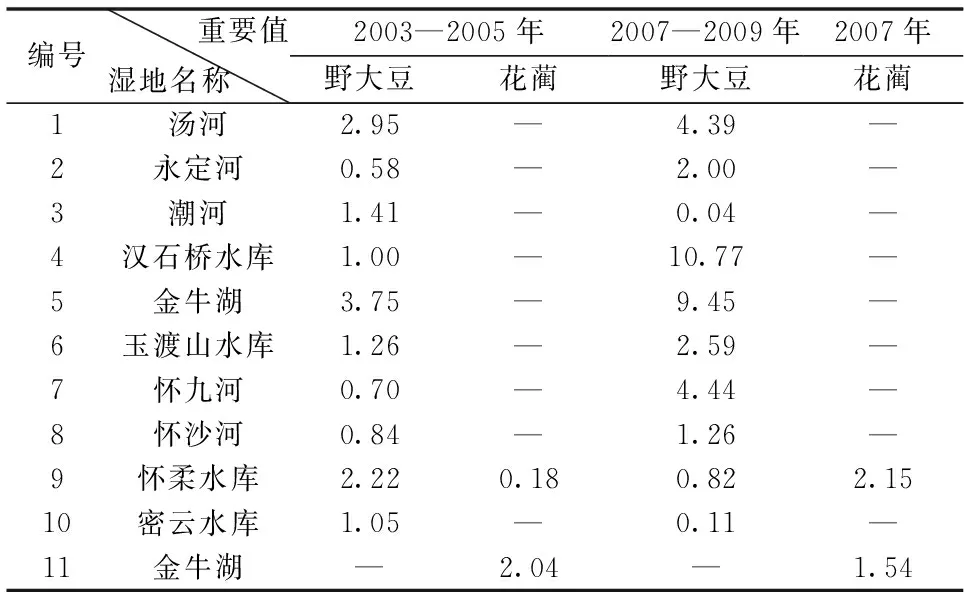
表2 北京湿地野大豆和花蔺重要值统计
2.2 北京湿地野大豆和花蔺重要值差异显著性分析
在调查的10处湿地中,金牛湖、汤河和怀柔水库在2003—2005年间野大豆的重要值较大,而2007—2009年间汉石桥水库、金牛湖、怀九河、汤河和玉渡山水库野大豆的重要值较大,且2007—2009年野大豆在各湿地中的重要值明显高于2003—2005年(见表2)。花蔺的分布区相对较为狭窄,且在金牛湖的优势度有所降低,而在怀柔水库的重要值增长较大。并且经过检验,北京湿地野大豆与花蔺在2003—2005年和2007—2009年的重要值不符合正态分布,因此,将其进行对数转换后进行方差分析,具体见表3所示。
由表3可以看出,野大豆和花蔺的P值均大于0.05,因此,调查的10处湿地的两种植物的重要值在2003—2005年与2007—2009年(2007年)间的差异性不显著。
2.3 群落特征分析
在本研究的典型湿地调查区域中,野大豆所在群落多为零星分布,且主要作为其他群落的伴生种或一些群落的中层优势种存在,在汉石桥湿地分布较多。野大豆群落常分为两层,野大豆为草质藤本,常在上层形成优势,下层植物由于缺少阳光导致生长不良甚至死亡。调查范围内常出现的是地草层、高草及乔灌层,部分群落层次结构缺失,仅剩下单层的结构,这是湿地环境退化的一个重要指示。野大豆所在的植物群落中,居于优势地位的是湿中生、中生和盐生植物。

表3 北京湿地野大豆和花蔺重要值方差分析
调查中发现花蔺典型湿地的分布范围主要集中在靠近水域的位置,如池塘边与河边浅水中,其群落主要分为两层,上层以花蔺为优势物种,盖度在40%上下,有时上层还生有薄荷、地笋、水棘针等典型的湿生植物;下层通常生有眼子菜等水生植物,有时水面上还生有荇菜。
由于调查时样方是采用沿着水位梯度变化顺次排列的,因此,样方在样带中的位置体现其距离水的远近。野大豆生长在离水相对较远的地方,而花蔺喜湿润,多生长在常年积水的池沼,洼地或沿河湿沙地。随着水分梯度的变化,远水区和近水区的植物差异大,野大豆以及花蔺的伴生种依次从水生或湿生高大草本到湿中生草本逐渐过渡,两种植物的水平分布格局均表现为斑块状,没有明显的带状分布。
2.4 种间联结
由表4可知,10处典型湿地野大豆与其他物种的关系多数为正联结,大约有1/5的物种与野大豆为负联结。其中,苍耳、狗尾草、葎草、蒙古蒿、山楂叶悬钩子、问荆等与野大豆的正关联性极强;狼把草、鬼针草以及浮萍与野大豆的AC值为0,则表明物种间完全独立。汤河有11种植物与野大豆正联结,4种植物负连接,1种植物完全独立;永定河23种植物与野大豆正联结,9种植物负连接;潮河有4种植物强正联结;汉石桥水库有20种植物与野大豆正联结,6种植物负连接,2种植物完全独立;玉渡山水库有3种植物强正联结,4种植物正连接;怀九河有18种植物与野大豆正联结,4种植物负连接;密云水库有8种植物与野大豆正联结,1种植物负连接;金牛湖有21种植物与野大豆正联结,4种植物负连接;怀沙河有16种植物与野大豆正联结,3种植物负连接;怀柔水库有12种植物与野大豆正联结,1种植物负连接。
花蔺仅在金牛湖、怀柔水库以及怀沙河3处被发现,其与其他物种间的关系大多数是正联结的,只有水棘针和野大豆与其为负联结。其中,紫萍、金鱼藻、水鳖和假稻与花蔺间有极强的正联结。金牛湖有16种植物与花蔺正联结,1种植物负连接;怀沙河有16种植物与花蔺正联结,1种植物负连接;怀柔水库有4种植物与花蔺强正联结,2植物与花蔺正联结,1种植物负连接。
将调查的10处典型湿地当中的野大豆和花蔺与其他物种的种间联结度进行分析,检验结果以是否显著关联为依据,其他不显著的物种未列出,具体如表4所示。
由表5可以看出,野大豆与绵毛酸模叶蓼和三叶鬼针草等在内的18种植物显示极显著关联,与车前和芦苇等在内的19种植物显示显著关联;花蔺与鼠掌老鹳草等在内的10种植物显示极显著关联,与蒙古蒿等在内的4种植物显示显著关联。结合表3和表5,发现与野大豆和花蔺两个物种间关联程度较高的正联结物种在χ2检验中均是极显著关联,表现出与χ2检验一致性。野大豆的伴生物种常常是芦苇、蒙古蒿、葎草、三叶鬼针草、薄荷等;花蔺的伴生物种常常是泽泻、金鱼藻、紫萍、马来眼子菜等,这些伴生物种与野大豆和花蔺间的关联性极显著(表5)。上述10个湿地(汤河、永定河、潮河、汉石桥水库、玉渡山水库、怀九河、密云水库、金牛湖、怀沙河及怀柔水库)当中与大豆存在显著或极显著关联的物种数分别是1,7,1,4,3,4,6,8,2,3;与花蔺存在显著或极显著关联的物种数分别是0,0,0,0,0,0,0,3,4,7。
3 讨论
本研究以北京10处典型湿地所分布的野大豆和花蔺为研究目标,通过对其不同年份重要值变化及与不同植物的种间联结性分析,得出两种植物的分布特征及群落特征。植物的重要值可以反映某个种在群落中的地位和作用,重要值的高低也可以一定程度反映植物群落数量及频次和密度的相对变化[12]。野大豆在2003—2005年和2007—2009年两个阶段在10个典型湿地重要值的变化情况存在差异,其中汤河、怀沙河、怀柔水库3个位置的重要值变化不大,表明野大豆在这些地区的生长状况及种群数量相对较稳定;而潮河、密云水库野大豆重要值前一个阶段要显著高于后一个阶段,表明野大豆的数量、分布区域和出现频次有所下降;永定河、汉石桥水库、金牛湖、玉渡山水库和怀九河5个湿地的野大豆的重要值后一个阶段高于前一个阶段,表明近年来野大豆的生长状况相对良好。花蔺在怀柔水库的重要值变化较大,其在2007年的重要值远远大于2003—2005年,说明在怀柔水库花蔺的生长情况相对北京其他湿地较好一些;而花蔺在金牛湖的重要值变化虽然不大,但也说明花蔺的种群数量已经有所减少。方差分析结果表明,野大豆和花蔺的重要值在2003—2005年和2007—2009年皆无显著性差异。而由前面所述可知,大部分湿地在2003—2005年和2007—2009年野大豆和花蔺的重要值差异明显,说明从整个北京角度或更大尺度来看,野大豆或花蔺的重要值差异不明显,也就表明湿地之间的此消彼长互相抵消了差异。
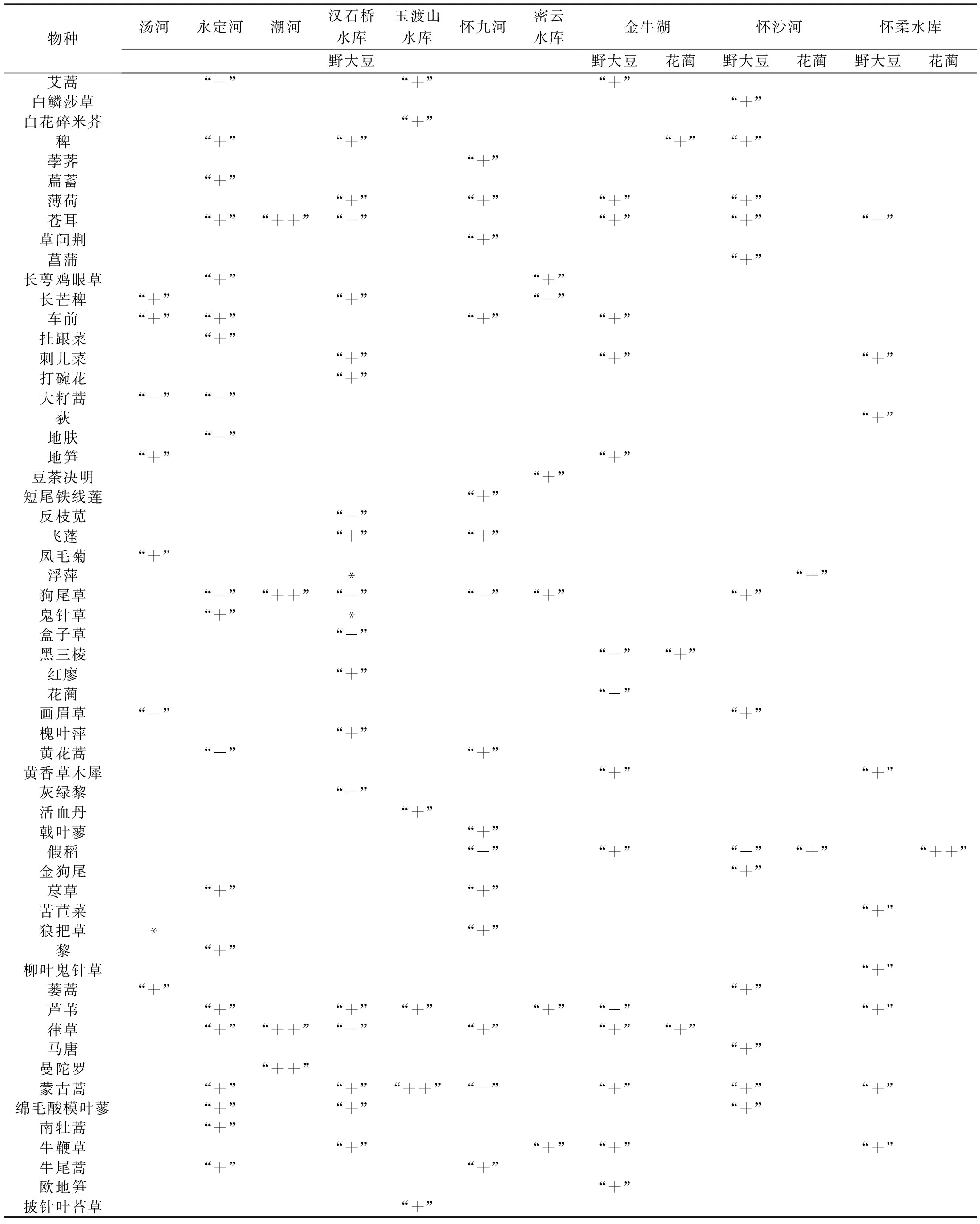
表4 北京湿地野大豆和花蔺与其他物种间的种间关联性
续表4 (Continued table 4)

物种汤河永定河潮河汉石桥水库玉渡山水库怀九河密云水库野大豆金牛湖野大豆花蔺怀沙河野大豆花蔺怀柔水库野大豆花蔺千屈菜“-”“+”荨麻“+”苘麻“+”求米草“+”“+”“+”“+”“+”球果蔊菜“+”“+”球穗莎草“+”三裂叶豚草“+”三叶鬼针草“+”山莴苣“+”山楂叶悬钩子“++”鼠掌老鹳草“+”“+”水棘针“+”“-”“+”“+”“-”“-”“-”水金凤“+”水蓼“+”“+”水芹“+”水莎草“+”粟草“+”酸模叶蓼“-”“-”“+”“+”“+”碎米莎草“+”透茎冷水花“+”菟丝子“+”豚草“+”萎陵菜“+”问荆“++”狭叶荨麻“+”香蒲“-”“+”小红菊“-”小箭叶蓼“+”小香蒲“+”旋覆花“+”“+”“+”“+”“+”鸭跖草“+”益母草“+”翼果苔草“+”茵陈蒿“-”“+”“+”圆叶牵牛“+”“+”沼生蔊菜“+”针蔺“+”“+”猪殃殃“+”扁秆藨草“+”藨草“+”“+”茶菱“+”长鬃蓼“+”单穗漂浮草“+”牛毛毡“+”萍“+”水鳖“++”水蜈蚣“+”四叶苹“+”线叶眼子菜“+”黑藻“+”狐尾藻“+”“+table_title+”“++”金鱼藻“++”鳢肠“+”马来眼子菜“+”“+”满江红“+”毛地笋“+”荇菜“+”
续表4 (Continued table 4)

物种汤河永定河潮河汉石桥水库玉渡山水库怀九河密云水库野大豆金牛湖野大豆花蔺怀沙河野大豆花蔺怀柔水库野大豆花蔺野慈姑“+”“+”野大豆“-”叶下珠“+”泽泻“+”
注:“+”表示正联结;“++”表示强正联结;“-”表示负联结;“*”表示物种间完全独立
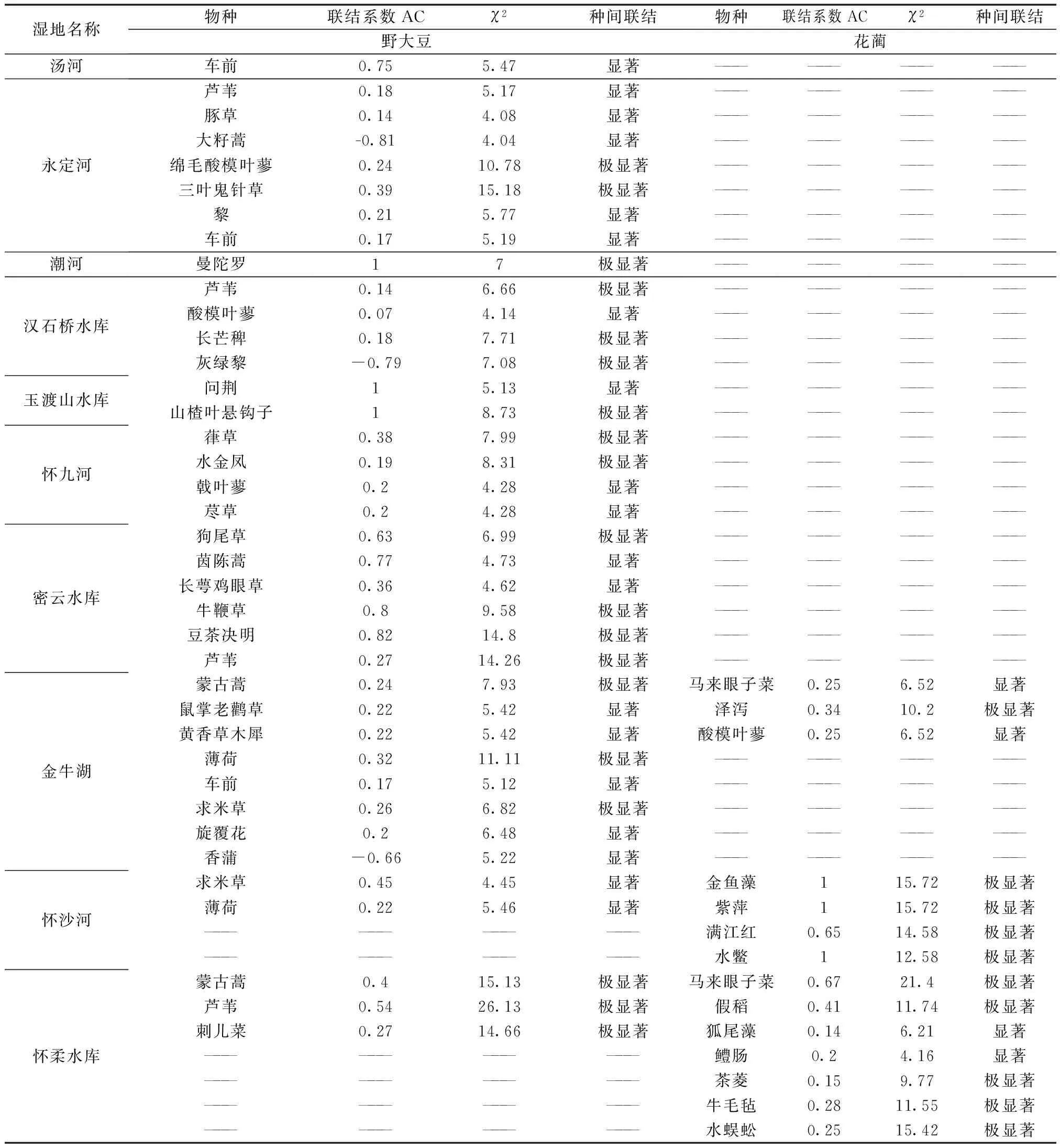
表5 北京湿地野大豆(花蔺)与其他物种的种间联结度
本研究发现调查区域的群落层次出现频率最高的是地草层、高草及乔灌层,但是也有一部分群落层次结构缺失,仅剩下单层的结构,这是湿地环境退化的一个重要指示。野大豆群落常分为两层,在上层形成优势,在调查的典型湿地当中主要作为其他群落的伴生种或一些群落的中层优势种存在,有时也可与不同植物形成群落,如:薄荷+野大豆+葎草群落(Ass.Menthahaplocalyx+Glycinesoja+Humulusscandens)[8]。花蔺群落也可以分为两层,上层以花蔺为优势物种,下层通常生有眼子菜等水生植物,有时水面上还生有荇菜,因而,在一些湿地的半干燥区域或退化湿地中其生长非常茂盛[8,11]。野大豆经常出现在远水区,表明野大豆对水分条件的要求并不高;花蔺则更多地出现在近水区,说明花蔺喜欢温暖湿润的环境。野大豆群落和花蔺群落水平分布格局均表现为斑块状,没有明显的带状分布[12]。
种间联结指标是对一定时期内群落组成物种之间相互关系的静态描述,随群落演替阶段的变化而变化[13]。野大豆在调查的10处典型湿地当中与其他物种的关系多数为正联结,其中,苍耳、狗尾草、葎草、蒙古蒿、山楂叶悬钩子、问荆等与野大豆的正关联性极强,这可能是由于它们对综合环境条件具有相同或相似的需求与适应,生态习性相似,却都避免相同的生态因子;狼把草、鬼针草以及浮萍与野大豆之间则完全独立。种间联结测定值在一定程度上衡量了物种间的相互关系和植物对环境因子的适应程度。正联结体现了物种利用资源的相似性和生态位的重叠性,说明它们之间具有相似的生境要求[14]。在金牛湖、怀柔水库以及怀沙河三地,花蔺与其他物种间的关系大多数是正联结的,只有水棘针和野大豆与其为负联结。其中,紫萍、金鱼藻、水鳖和假稻与花蔺间有极强的正联结,可见它们对周围环境的差异都有相似的反应。并且与野大豆和花蔺两个物种间关联程度较高的正联结物种在χ2检验中均是极显著关联,表现出与χ2检验一致性。野大豆的伴生物种常常是芦苇、蒙古蒿、葎草、三叶鬼针草、薄荷等;花蔺的伴生物种常常是泽泻、金鱼藻、紫萍、马来眼子菜等,这些伴生物种与野大豆和花蔺间的关联性极显著。联结系数和点相关系数分析表明:与野大豆显著或极显著正关联的物种占总物种数目的31.09%,其中显著正联结19种,极显著正联结18种;与花蔺显著或极显著正关联的物种占总物种数目的11.76%,显著正联结4种,极显著正联结10种。总体而言,野大豆与花蔺与周围物种呈现无显著关联分别占所有植物总数的68.91%和88.24%。由此看出:野大豆与花蔺与周围物种中间联系较少,种间联结关系较为松散而趋向于相互独立,有很大的随机性。胡理乐等在研究濒危植物毛柄小勾儿茶的种间联结性时也得到类似的结果[15]。
同时需要指出的是宿敏采用样方调查法针对北京众多地区的濒危植物进行调查,发现花蔺的分布区与本研究区域存在差异[7],其主要原因之一是两调查时间不一致,前者主要是在2007—2009年,本文是在2003—2009年之间;原因之二是针对调查范围不一致,前者主要针对所有北京地区,而本文主要针对选取的10个典型湿地进行分析,当然这也成为本文的局限所在,后续应该扩大范围进行更加全面的研究。
4 就地保护对策与建议
野大豆的生长对光照要求较高,因此在对其种群进行保护的同时,要保证充足的光照强度和光照时间。花蔺对水分和光照条件的要求也较高,尤其是对水分的需求。北京湿地面临水体污染、水资源缺乏以及湿地退化等重大的问题,当务之急是要改善水质,恢复退化的湿地,为野大豆和花蔺的生长提供一个较为理想的生境,这样才能继续进一步地进行就地保护工作[16-17]。
野大豆和花蔺的就地保护工作,还要包含其伴生种的保护。这些伴生种在群落中虽然不起主要作用,但是在群落中经常出现,与野大豆和花蔺间具有一定的联系。因此,在对野大豆和花蔺进行相关的就地保护的同时,也要加强对其伴生物种的保护力度[18-19]。野大豆和花蔺所在群落中,也有与野大豆和花蔺负关联的物种,说明它们所需的环境条件不同或是一个种的存在对另一个种有排斥作用。在对野大豆和花蔺的伴生种进行适当保护的同时,也要控制其竞争种的种群数量。
野大豆在密云水库和潮河的优势度很低,种群数量也不多,应加强对此处野大豆的保护力度,建议建立自然保护小区或对野大豆的集中分布区域设置围栏、警示牌等,降低其所受人为干扰的强度[20]。花蔺在北京湿地中整体的分布区较为狭窄,种群数量也不多,应对其现存的种群加大保护力度,并适当的扩展其分布区和种群数量。
[1]陈 燕. 北京市湿地水生植物多样性研究[D].北京:北京林业大学,2008: 3-21.
[2]智颖飙,赵利清,王再岚,等. 内蒙古重点保护的野生植物及其基本特征[J].内蒙古大学学报(自然科学版),2003,34(6): 657-662.
[3]徐 豹. 中国野生大豆(G.soja) 研究十年[J]. 吉林农业科学,1989,29(1): 5-13.
[4]冯光海. 野大豆在黄河三角洲的现状及保护措施[J]. 中国林业,2007,7(6):39.
[5]胡志昂,王洪新. 北京地区野大豆天然群体遗传结构[J]. 植物学报,1985,20(6): 599-604.
[6]王书锦,付沛云,蔺继尚,等. 中国东北地区野大豆及其共生根瘤菌的生态分布与特性研究[J]. 微生物学杂志, 1989, 11(3): 35-40.
[7]宿 敏. 北京重点保护野生植物的分布特征研究[D].北京:北京林业大学,2011:12-28.
[8]雷 霆,崔国发,卢宝明. 北京湿地植物研究[M]. 北京:中国林业出版社,2010:121-125.
[9]王 辰,王英伟.中国湿地植物图鉴 [M]. 重庆:重庆大学出版社,2011:89-102.
[10]王 斌. 水生花卉花蔺[J]. 园林,2012,13(2): 60-61.
[11]李成茂,张 勇,田子珩. 北京植物种质资源调查内容与方法探究[J]. 北京林业大学学报,2010(s1):210-214.
[12]方精云,王襄平,沈泽昊,等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性,2009,17(6):533-548.
[13]刘萍萍, 程积民. 植物种间联结关系的研究[J]. 水土保持研究, 2000, 7(2): 179-184.
[14]MOFFATT S F, MCLACHLAN S M, KENKEL N C. Impacts of land use on riparian forest along an urban - rural gradient in southern Manitoba[J]. Plant Ecology, 2004, 174 (1) :119-135.
[15]胡理乐, 江明喜, 党海山, 等. 从种间联结分析濒危植物毛柄小勾儿茶在群落中的地位[J]. 植物生态学报, 2005, 29(2): 258-265.
[16]陈 燕,雷 霆. 北京湿地维管植物群落分类研究[J]. 河北林业科技,2008(5):13-18.
[17]吴东丽,张金屯,王春乙,等. 野生大豆群落主要种群生态位特征研究[J]. 草地学报,2009,17(2):166-173.
[18]袁 秀,刘 义,李俊清,等. 北京市自然保护小区建设方案与管理模式探讨[J]. 北京林业大学学报(社会科学版), 2010,9(1):59-64.
[19]张殷波,马克平. 中国国家重点保护野生植物的地理分布特征[J]. 应用生态学报, 2008,19(8):1670-1675.
[20]林大影. 北京山地珍稀濒危植物分布及其生存群落特征研究[D]. 北京:北京林业大学,2008.
Distribution and community characteristics of Glycine soja andButomusumbellatusand countermeasures of in situ conservation
ZHU Si-yu1, ZONG Xue2, ZHANG Ling1, LI Jing3, LEI Ting4
(1. China Wildlife Conservation Association, Beijing 100714; 2. Survey and Planning Institute of State Forestry Administration, Beijing 100714; 3. Municipal Land Transaction Center of Beijing, Beijing 100029;4. School of Nature Conservation, Beijing Forestry University, Beijing 100083, China )
Beijing suburbs wetlands are single and interfered greatly by human activities, thus some plants reduced or suffered distribution atrophy and would become endangered species. This article is based on the analysis about the distribution characteristics and interspecific association of the communities ofGlycinesojaandButomusumbellatusin Beijing wetlands, putting forward some measures and suggestions about the situ~conservation activities as a reference combining transect method with sampling method in Beijing wetlands. The investigations and records of the plants in the quadrats included name, quantity, average height and coverage and so on. It calculates the important values ofGlycinesojaandButomusumbellatusin the quadrats which are so important value as the basis. Then it made a comparison and variance analysis about the changes of the important values ofGlycinesojaandButomusumbellatusin Beijing wetlands at 2003-2005 and 2007-2009 and analyzed the difference and significance between those different data. It made the measure about the interspecific association of the community ofGlycinesojaandButomusumbellatusthrough the method of coupling coefficient AC and chi~square and analyzed the associativity between these two species and the other species in the community. The relation between the species like Mongolia Artemisia, sticktight andGlycinesojaorButomusumbellatusin the community was positive association while the the relation between the species like Bidens Tripartita,sticktight, duckweed andGlycinesojain the community was negative association. The relation between the species likeRhizomaalismatis,hornwort,Malay Pondweed andButomusumbellatusin the community was positive association. The positive association species of the higher degree of correlation were highly significant correlation in the χ2test, showing the consistence with the χ2test. We got the conclusion that there are a lot of interferences and threats which theGlycinesojaandButomusumbellatusin Beijing wetlands facing such as the environmental degereration and so on. There are some feasible countermeasures such as improvement of water quality, recovery of degraded wetland, conservations of the auxiliary species and controlling the population of the competitive species put forward in the paper as references .
Glycinesoja;Butomusumbellatus; community; species association; in situ conservation
2016-08-12;
2016-08-29
北京林业大学科技创新计划项目(Yx2010-34)
朱思雨,助理工程师,主要从事野生动植物保护与繁育,E-mail:zhusiyuemail@163.com
雷 霆,副教授,主要研究方向为湿地生态学,E-mail:leiting@bjfu.edu.cn
Q948.15
A
2095-1736(2017)02-0091-08
doi∶10.3969/j.issn.2095-1736.2017.02.091
猜你喜欢
植物研究(2023年5期)2023-09-09 08:01:22
今日农业(2022年16期)2022-11-09 23:18:44
中国化肥信息(2022年5期)2022-08-30 01:58:26
河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
今日农业(2021年20期)2021-11-26 01:23:56
今日农业(2021年14期)2021-10-14 08:35:34
食品安全导刊(2021年20期)2021-08-30 06:40:50
杂草学报(2015年2期)2016-01-04 14:57:55
水生生物学报(2015年1期)2015-02-28 16:01:05
