
热浪式连续高温胁迫对小菜蛾幼虫和蛹的影响
2017-04-12 22:32常向前吕亮张舒
湖北农业科学 2016年22期
常向前++吕亮++张舒

摘要:模拟高温热浪的温度模式,研究了热浪式连续高温胁迫对小菜蛾(Plutella xylostella)幼虫和蛹的影响。结果表明,高温胁迫时的初始虫态、胁迫温度、胁迫模式及各因子之间的交互作用都可能对小菜蛾3龄幼虫和蛹的羽化率及生殖力有显著影响。小菜蛾3龄幼虫比蛹耐热,蛹更易受到高温危害。虽然小菜蛾较耐高温,但40 ℃,持续3 d每天8 h(3 d×8 h/d)的高温胁迫下,相比30 ℃ 1 d 8 h(1 d×8 h/d),小菜蛾羽化率降低了55.48%。从生殖角度看,蛹经高温胁迫后生殖力显著低于3龄幼虫,3 d持续高温胁迫与 1 d高温相比,生殖力降低了48.84%,40 ℃胁迫相比30 ℃,生殖力降低了64.32%。持续长时间的高温热浪天气可能会导致小菜蛾种群数量的大幅度减少。
关键词:高温热浪;高温胁迫;羽化率;生殖力;小菜蛾(Plutella xylostella)
中图分类号:Q968.1;S433.4 文献标识码:A 文章编号:0439-8114(2016)22-5835-04
DOI:10.14088/j.cnki.issn0439-8114.2016.22.027
The Effects of Continuous High Temperature Stress on the Larvae
and Pupae of Plutella xylostella
CHANG Xiang-qian, L?譈 Liang, ZHANG Shu
(Institute of Plant Protection and Soil Science,Hubei Academy of Agricultural Sciences/Hubei Key Laboratory for Crop Diseases,
Insect Pests and Weeds Control, Wuhan 430064, China)
Abstract: The effects of continuous high temperature on the 3rd larvae and pupae of diamondback moth (DBM)(Plutella xylostella) emergence rate and fecundity were conducted by simulating heat waves. The results showed that the initial development stage of heat stress, stress temperature, heat stress patterns and the interaction between these factors all significantly may affect the emergence rate and fecundity of 3rd larvae and pupae. The larvae were more tolerant to the heat stress than the pupae. The pupae were more susceptible to high temperature stress. The emergence rate was decreased by 55.48% under the stress of successive 3 d 8 h/d of 40 ℃ compared to 1 d 8 h/d. The fecundity of pupae was significantly lower than larvae under high temperature stress; The fecundity of 3 d stress was decreased by 48.84% compared to 1 d stress and the fecundity of 40 ℃ stress was decreased by 64.32% compared to 30 ℃ respectively. The heat waves may lead the collapse of the DBM population in the fields.
Key words: heat waves; high temperature stress; emergence rate; fecundity; diamondback moth(Plutella xylostella)
近年来,随着全球平均气温的升高,极端天气现象,如高温热浪、暴雨、骤热骤冷等现象明显增加[1]。根据联合国政府间气候变化专门委员会的报告,全球范围内夏季高温热浪天气将持续频繁出现[2]。高温热浪天气意味着连续多天出现超过35 ℃的高温。昆虫是变温动物,更容易受到这种天气变化的影响[3]。
目前关于夏季高温对昆虫的影响已有报道,如夏季高温抑制了温室白粉虱(Trialeurodes vaporariorum)的发生[4],对水花生的重要天敌莲草直胸跳甲(Agasicles hygrophila)成虫生殖力有迅速降低的作用[5];夏季高温可能是制约二化螟(Chilo suppressalis)田间种群增长的重要因子[6];夏季高温导致了新疆地区棉蚜(Aphis gossypii)种群产生明显的崩溃[7,8],但高温热浪式的夏季高温如何影响昆虫的生物学特性尚无详细报道。
小菜蛾(Plutella xylostella)是危害十字花科作物的重要害虫[9]。在内蒙、新疆多次大暴发,造成近70万hm2油菜绝收[10,11],在长江中下游流域如湖北、安徽、江苏等及长江以南广大地区如浙江、福建、广东、广西等地发生世代多,抗药性突出。在夏季,长江中下游十字花科蔬菜田中小菜蛾种群数量普遍极低。已有的研究表明,在恒温条件下,30 ℃时,小菜蛾可以正常存活,但当温度超过30 ℃时,小菜蛾各发育阶段的存活率迅速下降;35 ℃时小菜蛾卵不能孵化,4龄幼虫、蛹均死亡,不能完成世代[12,13]。但在夏季田间,小菜蛾并不会受到恒定高温的胁迫,常会受到热浪式的高温胁迫。用气象记录仪Kestrel 4500 Pocket Weather Meters(Nielsen-Kellerman Co.,USA) 记录武汉甘蓝田中太阳直射条件下的24 h瞬时空气温度,发现在2012年7月20~24日,连续5 d出现持续6~8 h超过35 ℃的高温天气。田间发生的小菜蛾种群世代重叠严重[9],各个发育阶段同时大量存在,在高温季节各个发育階段都会受到田间高温胁迫。因此超过35 ℃的热浪式高温胁迫对小菜蛾不同发育阶段有何作用,并如何影响其夏季种群数量值得深入探讨。
本试验将模拟高温热浪天气对小菜蛾的作用,将经过不同模式高温胁迫后的小菜蛾转移至室温,观察其后继存活、生殖情况,探讨热浪式连续高温胁迫对小菜蛾不同发育阶段存活率及生殖力的影响,从而更好地揭示田间小菜蛾种群对夏季高温胁迫的响应机制。
1 材料与方法
1.1 虫源及饲养方法
以2008年秋采自湖北省农业科学院花椰菜(Brassica oleracea L. var. botrytis L.)上的小菜蛾为初始虫源,在25±1 ℃、RH 60%~70%、L∶D=16 h∶8 h的养虫室中用3~4叶盆栽甘蓝(Brassica oleracea L.var.capitata L.)饲养1代后,人工饲料(Southland products incorporatied,USA)在养虫盒(直径9 cm,高7 cm)内。
1.2 连续高温胁迫初始虫态为3龄幼虫的小菜蛾
将1~2日龄3龄幼虫放入12孔塑料培养板(型号TPP 92012,体积12.5 cm×8.3 cm×2.2 cm,每孔直径2.4 cm)的孔内,每孔3头幼虫,并加入适当的人工饲料,重复3个培养板,然后置于人工气候箱(HP300GS-C,武汉瑞华仪器设备有限责任公司)中,将3台人工气候箱分别于10:00~18:00(8 h)设置为各胁迫温度(30、35、40 ℃),其余时间(16 h)为25 ℃。分别在高温胁迫1 d、连续高温胁迫3 d后将小菜蛾移至25 ℃养虫室中观察幼虫的存活、化蛹、羽化情况。将羽化成虫成对单独饲养于培养皿(直径6 cm)内,逐日观察产卵量。计算幼虫的羽化率及生殖力,作为统计分析指标。幼虫羽化率为经高温胁迫后羽化的成虫数量与初始幼虫数量的比值。
1.3 连续高温胁迫初始虫态为蛹的小菜蛾
将30头蛹放入直径10 cm培养皿内,置于人工气候箱中,重复3个培养皿。气候箱的温度设置方法同“1.2”。处理后将蛹移至25 ℃养虫室观察蛹羽化及生殖情况。羽化成虫的生殖情况观察方法同“1.2”。计算蛹的羽化率及生殖力,作为统计分析指标。
1.4 统计方法
用SAS 6.12多因素试验统计进行方差分析,Duncans新复极差法进行多重比较。
2 结果与分析
2.1 连续高温胁迫后小菜蛾的羽化率及生殖力
3龄幼虫及蛹经高温胁迫后羽化及生殖情况见表1。由表1可知,除40 ℃ 3 d×8 h/d处理组合外,经各处理组合胁迫后小菜蛾的羽化率均超过70%,生殖力也超过30粒/雌。而40 ℃ 3 d×8 h/d处理后羽化率不足35%,生殖力基本为0。
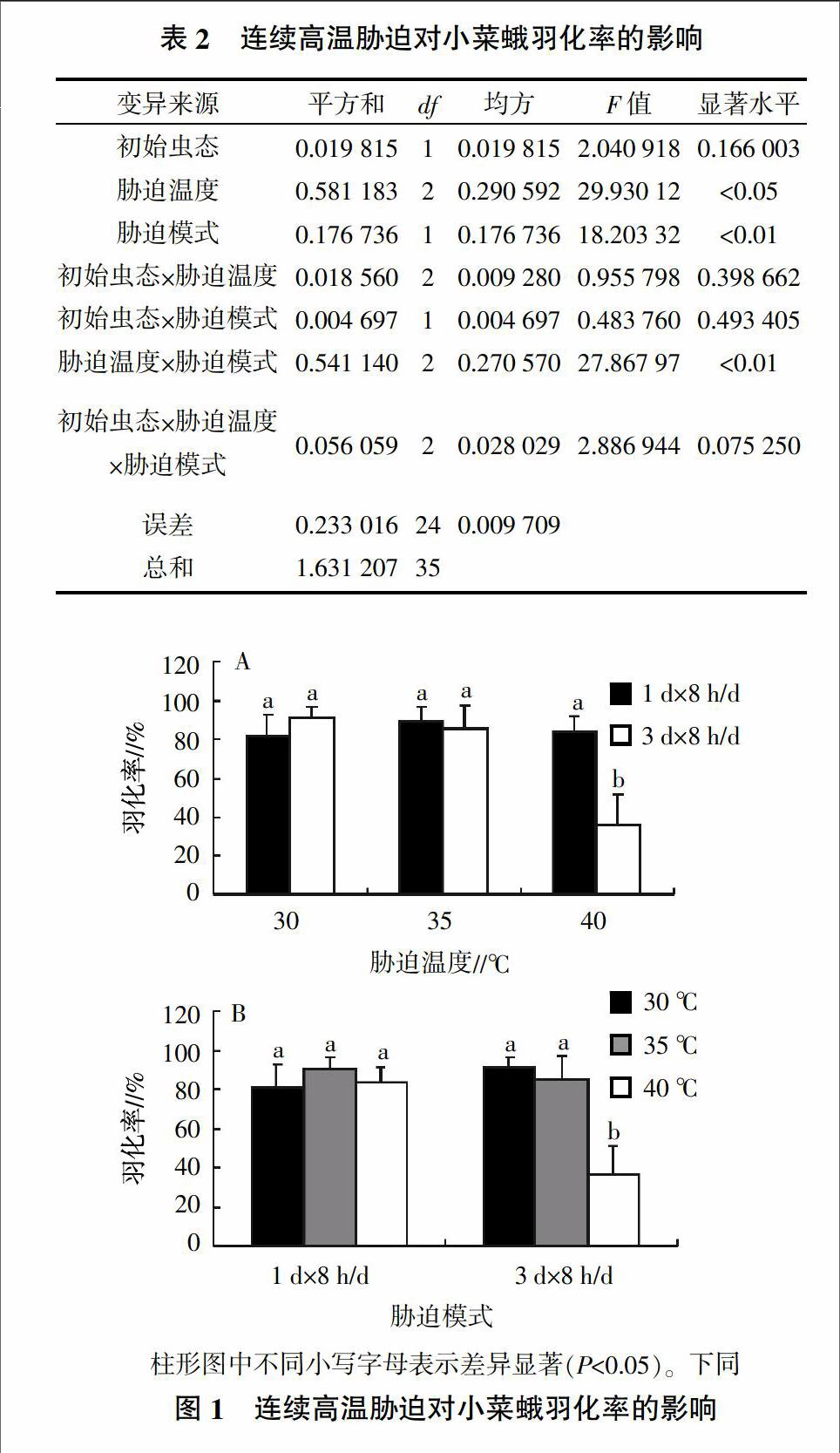
2.2 连续高温胁迫对小菜蛾羽化率的影响
多因素方差分析(表2)表明,胁迫温度及胁迫模式对小菜蛾的羽化率有显著影响;但初始虫态对小菜蛾的羽化率影响不显著,因此只对胁迫温度及胁迫模式进行多重比较分析(图1)。
当温度为30、35 ℃时,连续高温胁迫对小菜蛾的羽化率没有显著影响,羽化率为81.11%~91.11%;当温度为40 ℃时,经连续3 d×8 h/d的脅迫后羽化率显著下降为36.11%,与30 ℃ 1 d×8 h/d相比,羽化率降低了55.48%(图1A)。当胁迫时间为1 d时,各温度处理间没有显著差异,当连续时间为3 d时,40 ℃处理的羽化率显著下降(图1B)。
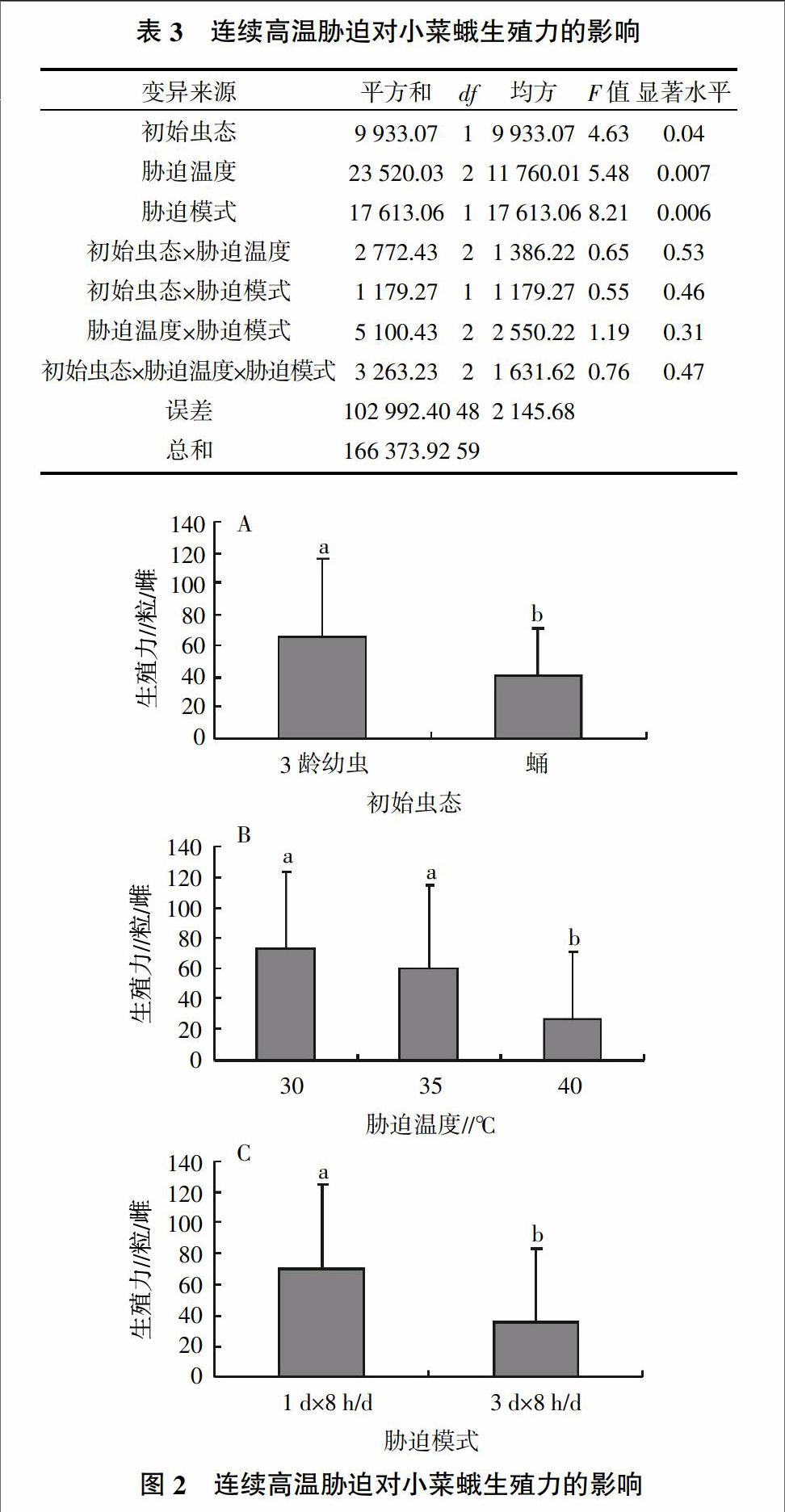
2.3 连续高温胁迫对小菜蛾生殖力的影响
多因素方差分析(表3)表明,小菜蛾初始虫态、胁迫温度及胁迫模式对生殖力均有显著影响,但二级和三级交互作用没有显著差异。因此,分别对初始虫态、胁迫温度及胁迫模式进行多重比较分析(图2)。
初始虫态为蛹时,受到高温胁迫后,生殖力显著低于初始虫态为幼虫的小菜蛾(图2A);而40 ℃相对于30、35 ℃显著降低了小菜蛾的生殖力,40 ℃相比30 ℃降幅为64.32%(图2B);3 d的连续高温可显著降低小菜蛾的生殖力,3 d持续高温胁迫与1 d高温相比,生殖力降低了48.84%(图2C)。
3 小结与讨论
有许多研究报道了高温对昆虫的致死效应[4-8]。Harcourt[14]系统研究了影响田间小菜蛾种群密度的生物因子及非生物因子,发现非生物因子温度的权重很大。在日本茨城县发现,夏季温度越高,小菜蛾种群密度越低[15]。昆虫不同龄期、性别及地理种群的耐热性可能不同[16]。本试验初步研究了热浪式高温胁迫对小菜蛾不同发育阶段的作用特点。结果表明,小菜蛾3龄幼虫比蛹耐热。虽然连续高温胁迫下,初始虫态为幼虫与初始虫态为蛹的羽化率差异不显著,但蛹的生殖力却显著下降。蛹在小菜蛾各个发育阶段中几乎不能移动,不能主动躲避高温胁迫,而幼虫却可以在小范围内主动躲避高温胁迫,蛹比幼虫耐热性差,导致当高温热浪发生时,很容易造成当代种群数量的大量减少。
小菜蛾整体比较耐热[17],但持续的热浪天气对小菜蛾种群数量有显著的负面作用。2龄幼虫在35 ℃处理12 h后死亡率仅为10%[18];在35 ℃处理8 h后,3龄幼虫的死亡率仅为3.13%[19];本研究发现,持续1 d×8 h/d的35 ℃胁迫未显著降低小菜蛾羽化率及生殖力,以上研究均说明当小菜蛾没有处于连续高温胁迫时,可忍受35 ℃高温,即小菜蛾本身对高温具有一定的抗性。但当连续胁迫时间为3 d时,40 ℃胁迫相对于30 ℃,羽化率降低了55.48%,而生殖力几乎降为0。根据气象仪的田间监测数据,夏季高温热浪天气常持续超过3 d,甚至达到5 d,如果热浪持续时间超过3 d,小菜蛾的羽化率将会进一步剧烈下降,说明持续长时间的热浪确实对小菜蛾种群数量有致命威胁。
本试验只选取了小菜蛾的两个发育阶段进行研究,因此仍需要对小菜蛾的其他虫态进行试验,同时根据田间温度的变化,设置更合理的模拟温度,使试验状态更接近田间环境,以期获得更接近实际条件的试验结果。
参考文献:
[1] 邵 勰,廖要明,柳艳菊,等.2015年全球重大天气气候事件及其成因[J].气象,2016,42(4):489-495.
[2] IPCC. Climate Change 2007:The Physical Science Basis—Summary for Policy Makers[M].Cambridge:Cambridge University Press,2007.
[3] DENLINGER D L,YOCUM G D. Physiology of Heat Sensitivity[M].Oxford:Westview Press,1998.
[4] 崔旭红,谢 明,万方浩.短时高温暴露对B型烟粉虱和温室白粉虱存活以及生殖适应性的影响[J].中国农业科学,2008,41(2):424-430.
[5] 赵 鑫,傅建炜,万方浩,等.短时高温暴露对莲草直胸跳甲生殖特性的影响[J].昆虫学报,2009,52(10):1110-1114.
[6] 罗 举,张孝羲,翟保平,等.高温对二化螟实验种群生长、存活和繁殖的影响[J].生态学报,2005,25(4):931-936.
[7] 吕昭智,张江国,罗 亮,等.短时间高温处理对棉蚜存活的影响[J].应用昆虫学报,2010,47(4):685-689.
[8] 高桂珍,吕昭智,夏德萍,等.高温胁迫及其持续时间对棉蚜死亡和繁殖的影响[J].生态学报,2012,32(23):7568-7575.
[9] 柯礼道,方菊莲.小菜蛾生物学研究——生活习性的观察[J].植物保护学报,1980,7(3):139-144.
[10] 王成德,李 谦,张士元,等.锡盟油菜主要虫害种类及其综合防治技术[J].内蒙古农业科技,1995(4):18-20.
[11] 王承君.小菜蛾的发生与综合防治[J].新疆农业科学,1999(6):287-288.
[12] 但建国,梁广文.不同温度条件下小菜蛾实验种群的研究[J].华南农业大学学报,1995,16(3):11-16.
[13] 陈非洲,刘树生.恒温和变温对小菜蛾发育速率的影响[J].生态学报,2003,23(4):688-694.
[14] HARCOURT D G. Biology of the diamondback moth,Plutella maculipennis(Curt.)(Lepidoptera:Plutellidae),in Eastern Ontario.II. Life-history,behaviour,and host relationships[J].The Canadian Entomologist,1957,89(12):554-564.
[15] SHIRAI Y. Temperature tolerance of the diamondback moth, Plutella xylostella(Lepidoptera:Yponomeutidae) in tropical and temperate regions of Asia[J].Bulletin of Entomological Research,2000,90(4):357-364.
[16] LI H B,SHI L,LU M X,et al. Thermal tolerance of Frankliniella occidentalis:Effects of temperature,exposure time,and gender[J].Journal of Thermal Biology,2011,36(7):437-442.
[17] 常向前,馬春森,张 舒,等.小菜蛾的耐热性[J].应用生态学报,2012,23(3):772-778.
[18] GU H. Cold tolerance and overwintering of the diamondback moth(Lepidoptera:Plutellidae) in southeastern Australia[J].Environmental Entomology,2009,38(3):524-529.
[19] 田素芬,高 飞,顾晓军.氟虫腈对小菜蛾3龄幼虫取食、化蛹、羽化的影响[J].江西农业学报,2008,20(7):73-76.
猜你喜欢
植物保护(2022年1期)2022-02-10
农业技术与装备(2022年11期)2022-01-01
上海农业科技(2020年5期)2020-10-24
植物保护(2020年2期)2020-06-08
四川动物(2019年3期)2019-07-08
蔬菜(2018年9期)2018-09-21
上海农业学报(2017年5期)2017-06-04
山西农业大学学报(自然科学版)(2015年3期)2015-04-19
华南师范大学学报(自然科学版)(2011年2期)2011-11-20
华南师范大学学报(自然科学版)(2010年1期)2010-11-25
